洮河自然保護區大峪溝林區紫果云杉群落木本植物種群生態位特征
2022-03-30 10:58:22曹家豪陳學龍高本強趙艷麗劉錦乾曹秀文
生態學報 2022年5期
趙 陽,曹家豪,李 波,齊 瑞,劉 婷,陳學龍,高本強,趙艷麗,劉錦乾,曹秀文,*
1 甘肅省白龍江林業管理局林業科學研究所,蘭州 730046 2 甘肅白龍江森林生態系統定位觀測研究站,舟曲 746300
生態位(niche)概念自被Grinnell[1]引入生態學研究領域以來,有關其定義和內涵一直存在諸多爭議,截至目前尚未形成統一共識[2—3],因此生態位研究一直是生態學的重要課題。100多年來,生態位理論在不斷豐富和發展中已成為解釋自然群落中物種共存與競爭機制的基本理論[4—5],被應用到種群、群落以及生態系統等復雜組織水平上。研究森林群落優勢種群生態位能客觀地反映種群對資源的利用能力及其在群落或生態系統中的功能和地位[6—7],還可以量化物種之間、物種與環境之間的相互關系[8],反映群落結構與功能、生物多樣性、物種進化以及群落演替的方向等[9—10]。這對群落生物多樣性保護,揭示種群生存策略和機制[11]及預測群落發展趨勢具有重要意義。
洮河自然保護區位于洮河上游,地處青藏高原東部邊緣的甘南山區與黃土高原過渡區,屬青藏高原濕潤氣候與高原大陸性季風氣候交匯帶[12]。獨特的地理環境和氣候條件孕育出豐富多樣的生態類型,作為黃河的一級支流,洮河上游的高原濕地、草原、森林等共同構成生態屏障維持著洮河乃至黃河上游區域的生態平衡,森林作為最頂級的生態類型,發揮著第一屏障作用。以紫果云杉(Piceapurpurea)為主要建群種的原始林群落在本區域中高海拔有著大面積分布,經過長期的適應與自然選擇已形成完整的生態系統結構,在物種多樣性保護、局域生態安全維護等方面發揮著極為重要的作用。截止目前,本區域有關紫果云杉的研究多存在于種群結構[12—13]和育苗等方面,但其群落組成與結構如何,生存狀態如何,種間競爭與共存關系如何,相關研究尚不多見。為此,本研究以紫果云杉群落為對象,通過分析木本植物種群生態位特征,研究群落組成與結構、物種多樣性、種間關系以及物種與群落環境的關系,揭示種群對資源和空間的利用能力、群落生存狀態與發展趨勢,為種群和群落的保護提供理論依據。
1 研究區與研究方法
1.1 研究區概況
研究區位于洮河國家級自然保護區大峪溝林區。大峪溝林區位于甘肅省甘南州卓尼縣木耳鎮大峪溝(103°33′—104°02′E,34°10′—34°52′N),面積約10.5萬hm2,林區濕潤多雨,光照充足,四季不明,小氣候多樣。海拔2500 m—4920 m,相對高差大,為典型的高原山地地貌。年均日照2266 h,年均溫2.3℃,無霜期105 d,年均降水640 mm,河流縱橫,水資源豐富,土壤主要為棕、褐色森林土[14]。植被類型多樣,陽坡主要為灌叢、高山草甸等,陰坡、河谷地帶主要是以云杉(Piceaasperata)、冷杉(Abiesfabri)等為建群種的原始暗針葉林群落,森林結構相對完整,覆蓋率50.74%,林下天然更新良好[15]。主要樹種有紫果云杉、岷江冷杉(Abiesfaxoniana)、樺木(Betula)、柏木(Cupressus)等,森林質量總體較高,野生動植物資源豐富。
1.2 樣地設置
2020年7—8月,在對大峪溝林區進行了全面踏查的基礎上,采用典型抽樣原則設置50 m×50 m的紫果云杉樣地8塊(表1),逐一測量并記錄樣地內胸徑大于3 cm的全部喬木樹種的胸徑、樹高、冠幅,喬木層郁閉度等指標。并在每個樣地設置3個10 m×10 m的灌木樣方,記錄各樣方內的灌木種類、數量、地徑、高度、蓋度。各樣地用GPS定位,記錄海拔、坡度、坡向等立地因子。
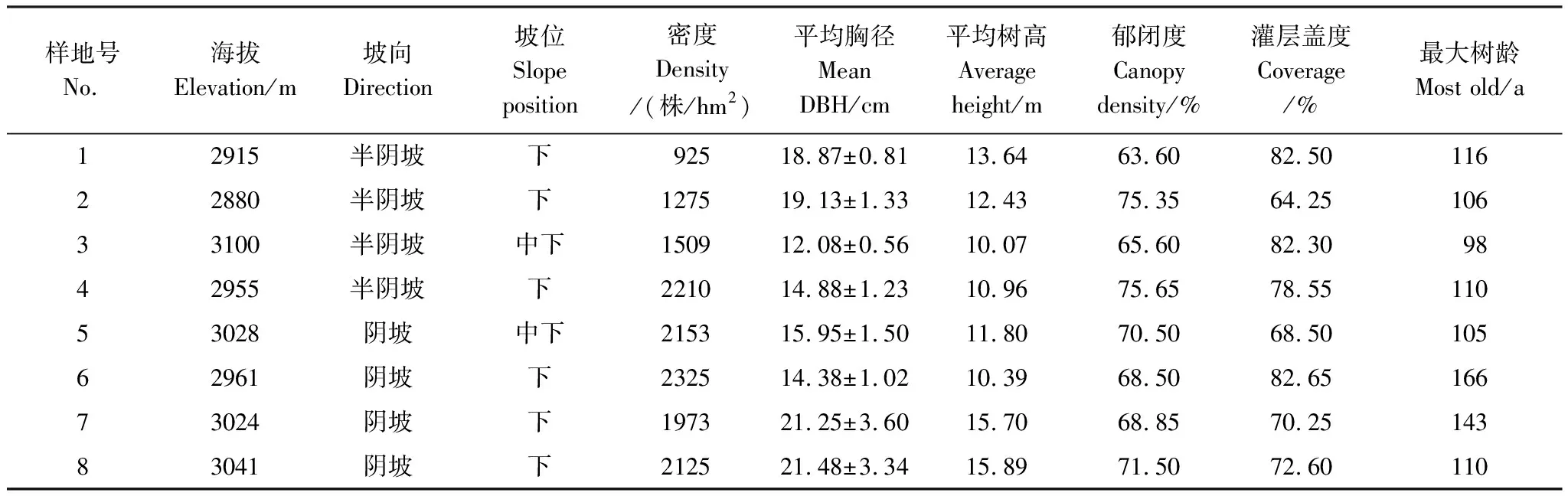
表1 樣地基本信息
1.3 研究方法
1.3.1重要值和頻數
重要值取物種在每個樣地的平均值,計算方法參見文獻[16—17],頻數為該物種出現的樣地、樣方數。
1.3.2生態位
以樣地代表多種資源的綜合狀態[16],用Levins指數BL[17]和Shannon指數BS[6]計算生態位寬度:
(1)
(2)
式中,Bi為第i物種的生態位寬度,Pij為第i物種在第j個資源水平上的重要值占該物種在所有資源水平上重要值總和的比例,Pij=nij/∑nij,nij為物種i在第j資源位上的重要值,r為樣方數。
生態位總寬度[18]B:
(3)
式中,Bi為物種i的生態位寬度,n為物種數。
生態位相似[19]Cik:
(4)
式中,Cik為物種i和k的生態位相似性,Cik∈[0,1],Pij和Pkj分別為物種i和k在資源位j上的重要值。
生態位重疊[20]Oik:
(5)
式中,Oik為物種i和物種k的生態位重疊指數,當Oik= 1時,物種i和k在所有資源位中分布完全相同,即物種i與k生態位完全重疊;當Oik= 0時,兩個物種無共同資源狀態,生態位完全不重疊。
采用Excel2010和SPSS21.0軟件對數據進行統計、計算和分析。
2 結果與分析
2.1 物種組成與重要值特征
紫果云杉群落喬木層共有樹種8種(表2),隸屬于4科6屬,重要值大小為紫果云杉>岷江冷杉>祁連圓柏>紅樺>糙皮樺>秦林柳>云杉>刺柏,紫果云杉重要值遠大于其它樹種,為該群落的建群種,岷江冷杉為次優勢種。灌木層共有植物50種,隸屬于16科25屬(表3),重要值大于2的有22種,其中最大的為峨眉薔薇(16.690),其次為唐古特忍冬(14.103)、藍靛果(12.174)和蔥皮忍冬(10.567)等。喬木樹種在灌木層均有幼苗存在,其中紫果云杉與岷江冷杉的幼苗重要值較大,分別為4.872和4.912。總體來看,灌木層薔薇科物種最多(14種),其重要值總和為45.32,小于忍冬科(11種)的61.64,表明忍冬科為灌木層第一優勢科,其次為薔薇科,茶藨子科(也叫醋栗科)(6種)以9.20的重要值位居第3。
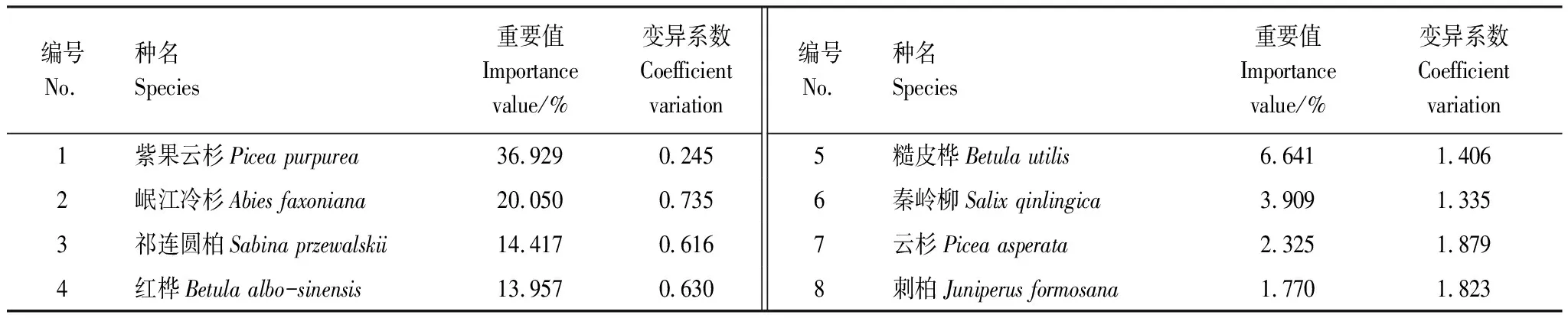
表2 紫果云杉群落喬木層重要值
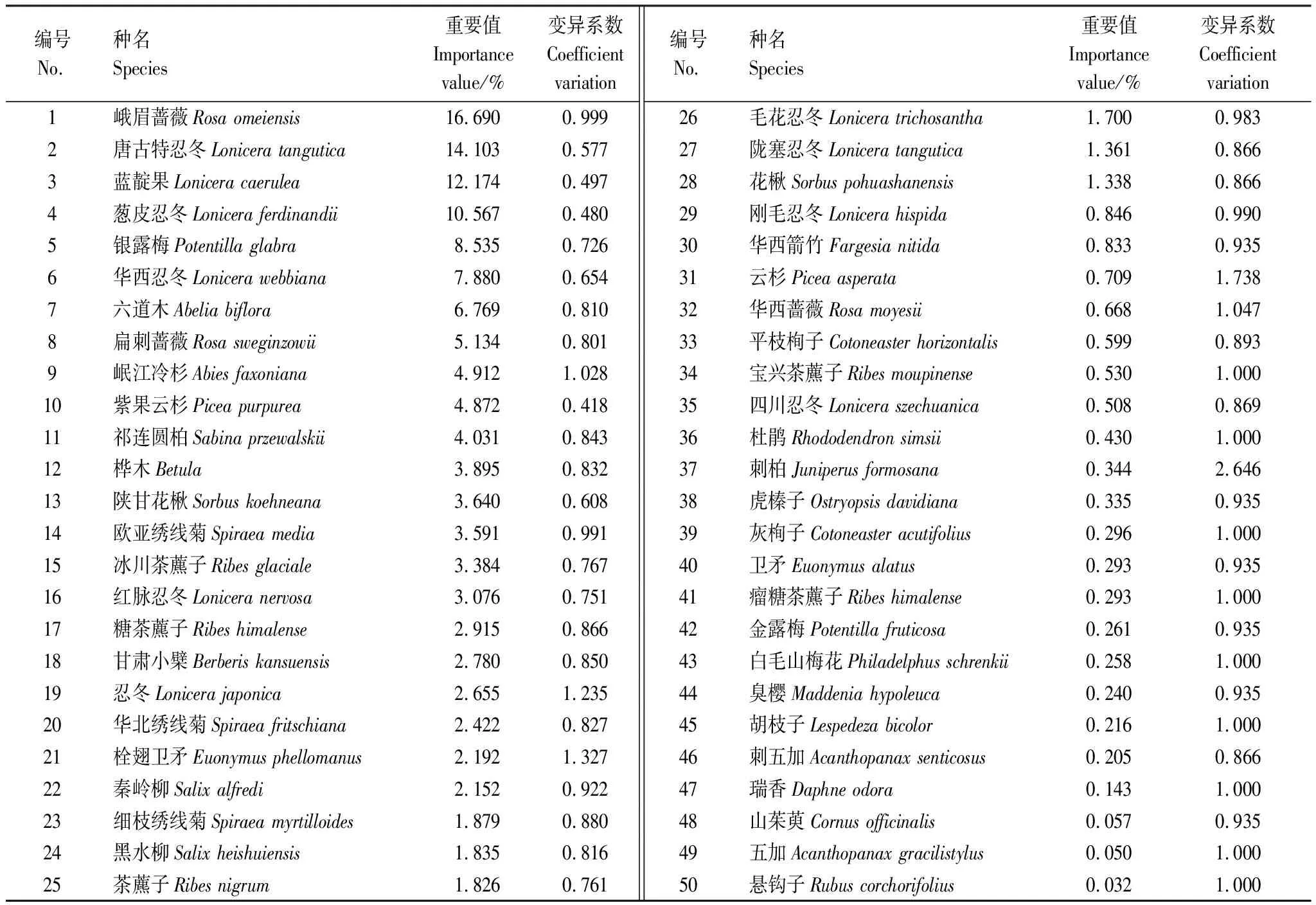
表3 紫果云杉群落灌木層重要值
2.2 生態位寬度
如表4所示,喬木層重要值最大的紫果云杉其生態位寬度(BL、BS)也最大,為7.548和2.050,其次,紅樺、祁連圓柏和岷江冷杉三者生態位寬度差異不大,云杉和刺柏生態位寬度最小。前4種優勢樹種BL和BS生態位寬度大小均為紫果云杉>祁連圓柏>紅樺>岷江冷杉,而后4種弱勢種BL和BS生態位寬度大小出現差異,可見不同物種Levins指數和Shannon指數大小變化不同,但總體趨勢一致。
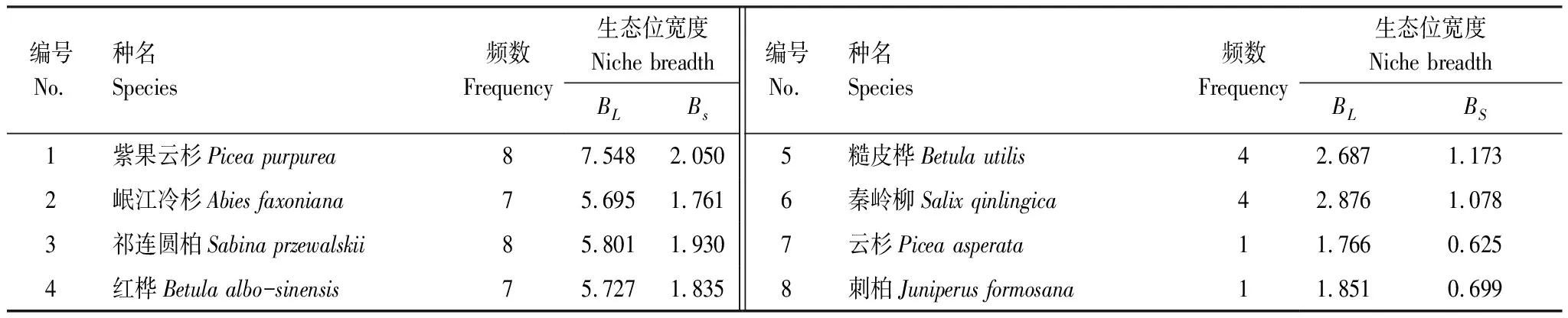
表4 紫果云杉群落喬木層生態位寬度
灌木層生態位寬度(BL、BS)最大的為紫果云杉幼苗(6.807、1.979)(表5),其次為忍冬科的藍靛果(6.414、1.958)、蔥皮忍冬(6.249、1.885)、唐古特忍冬(6.002、1.892)和華西忍冬(5.601、1.860),薔薇科生態位寬度最大的3個種為陜甘花楸(5.468、1.787)、銀露梅(4.795、1.717)和扁刺薔薇(4.402、1.701),這8個種除扁刺薔薇BS外,生態位寬度均大于重要值最大的峨眉薔薇(4.005、1.707)。茶藨子科生態位寬度最大的3個種為冰川茶藨子(4.133、1.525)、茶藨子(3.458、1.291)和糖茶藨子(3.123、1.340)。在3個優勢科中,忍冬科11種植物的生態位總寬度(13.654、4.466)大于薔薇科14種植物(11.183、4.079)及茶藨子科6種植物的生態位總寬度(6.464、2.406),再次表明忍冬科為灌木層第一優勢科。此外,樺木(包括紅樺和糙皮樺)(4.728、1.662)、祁連圓柏(4.675、1.625)及岷江冷杉(3.891、1.489)的幼苗也有較大的生態位寬度。生態位寬度BL和BS最小值為1和0的有16種,如虎榛子、臭櫻等,表示該物種在所有樣方中僅出現1次,為該群落的偶見種。
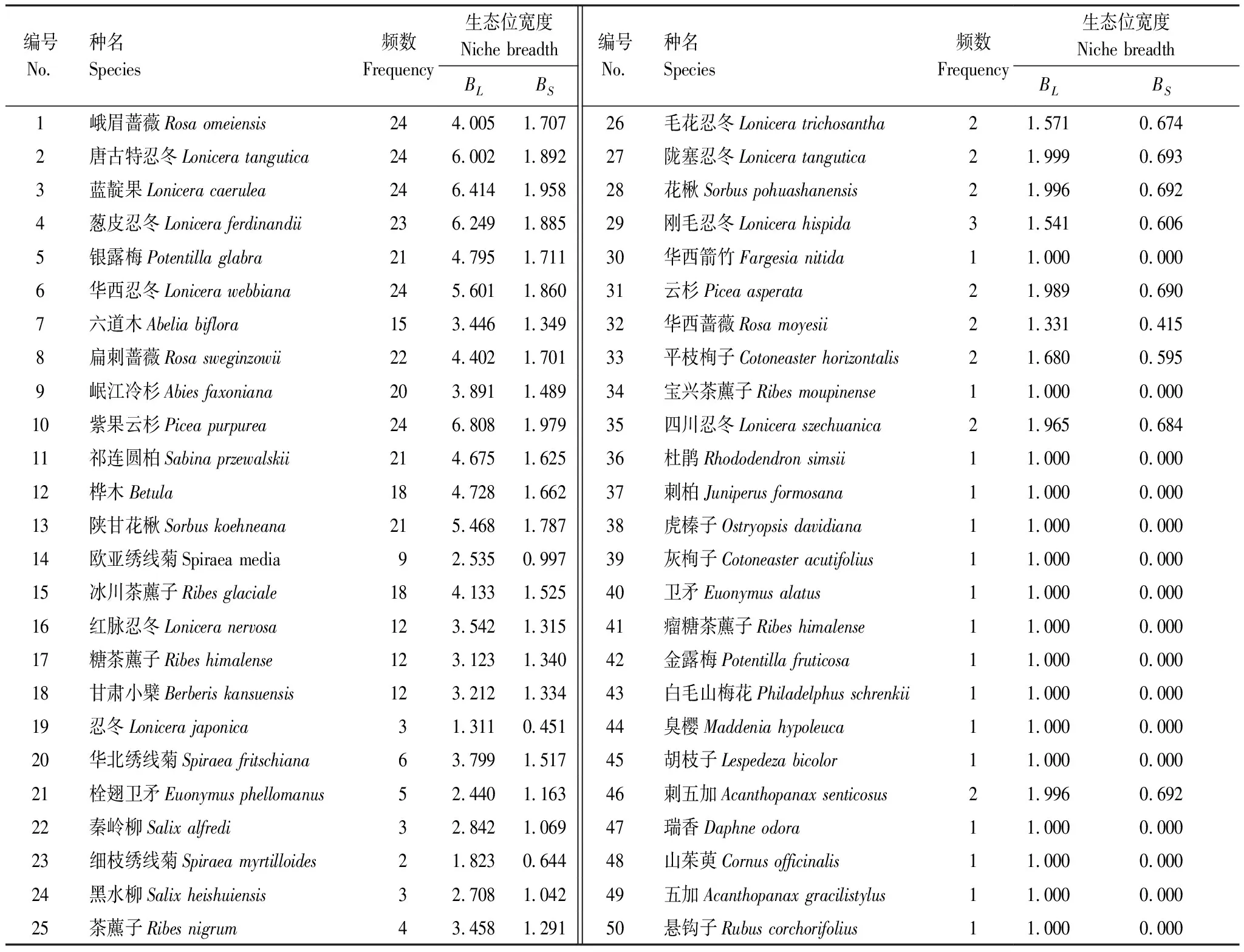
表5 紫果云杉群落灌木層生態位寬度
綜上可知,重要值大的物種生態位寬度往往較大,但也存在一定差異。如在喬木層,岷江冷杉重要值大于祁連圓柏,但生態位寬度卻小于祁連圓柏。灌木層中峨眉薔薇重要值最大,但生態位寬度卻僅為第12,紫果云杉、岷江冷杉的幼苗重要值分別為第10和第9,但生態位寬度卻為第1和第10。結合表2、3和4、5可以看出,生態位寬度除與重要值有關外,還與物種頻數及重要值變異系數有關,即物種頻數越大、重要值變異系數越小,其生態位寬度越大。
2.3 生態位相似
紫果云杉群落8種喬木樹種共構成28個種對(表6),生態位相似平均值為0.390,相似性大于0.6的有7對,占總對數的25.0%,前4個優勢種之間生態位相似平均值大于后4個弱勢種。相似性最大的種對為岷江冷杉與紅樺(0.722),其次為紫果云杉與祁連圓柏(0.717)、紫果云杉與紅樺(0.715),弱勢種秦嶺柳與云杉、糙皮樺與刺柏的相似性也在0.6以上。相似性最低的為岷江冷杉與刺柏(0.073),其次為岷江冷杉與云杉、秦嶺柳,紅樺與云杉等。
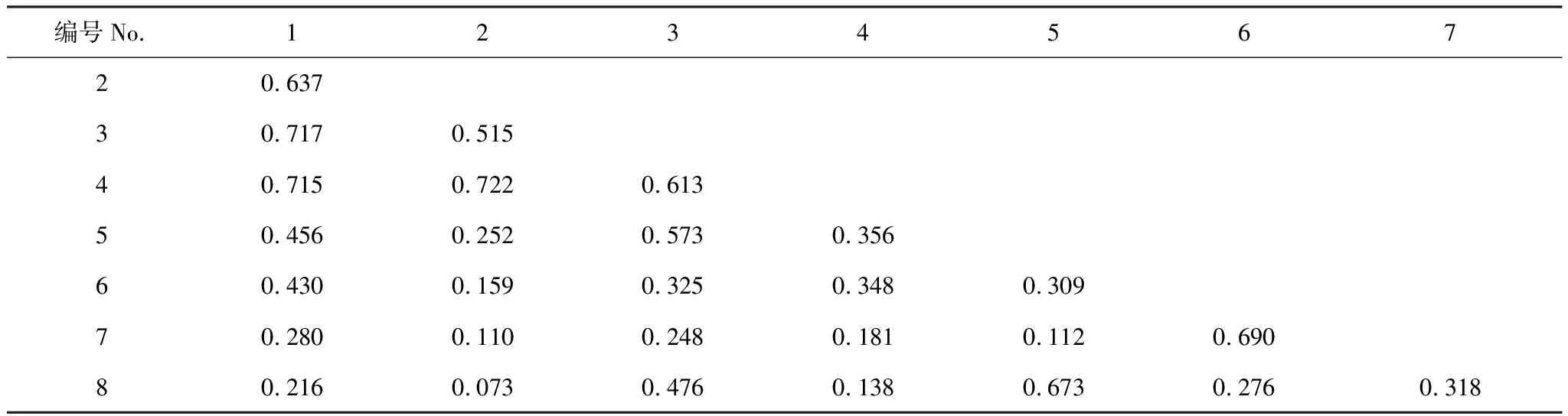
表6 紫果云杉群落喬木層生態位相似
灌木層50種植物共構成1225個種對,生態位相似平均值為0.217,總體相似性較低。其中相似性小于0.6的有1141對,占總對數的93.14%,絕大多數種對相似性介于0—0.4之間,包括相似性為0的365對。相似性大于0.6的有84對,僅占6.86%,其中0.6—0.8的59對,0.8—0.9的7對,0.9—1.0的2對,相似性為1的有16對。相似性大的如剛毛忍冬和毛花忍冬(0.911)、云杉與花楸(0.978)等以及相似性為1的種對多出現在非優勢種之間。前22個優勢或相對優勢種構成的231個種對(表7)生態位相似平均值為0.452,相似性大于0.6的有46對,占19.91%,其中0.6—0.8的43對,0.8以上3對,相似性最大的為藍靛果與蔥皮忍冬(0.815),其次為唐古特忍冬與銀露梅(0.806)、蔥皮忍冬與扁刺薔薇(0.800)。
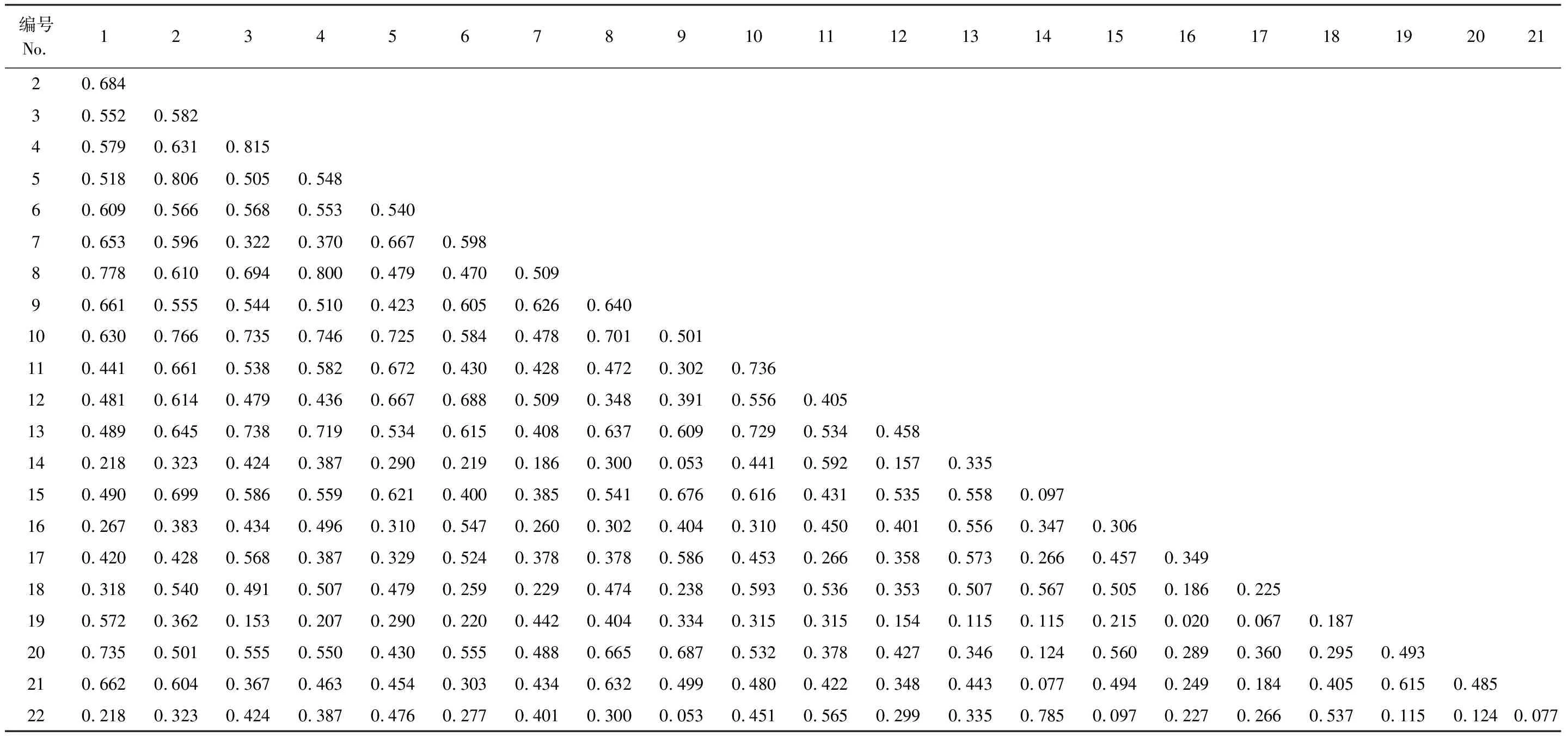
表7 紫果云杉群落灌木層生態位相似
綜上可知,紫果云杉群落喬木層各物種之間生態位均有相似性,且相似性大于灌木層。生態位寬度大的物種間生態位相似性一般較大,但非優勢種之間也可見較大的生態位相似性。灌木層前22個優勢或相對優勢種之間未出現相似性為0或1的種對。相似性為0,表明兩物種未在同一資源位中出現,相似性為1,表示兩物種在同一資源位出現的頻度相同,此外同科或同屬的物種之間可見較大的生態位相似性。
2.4 生態位重疊
紫果云杉群落8種喬木樹種間均存在生態位重疊(表8),重疊均值為0.462,重疊值大于0.6的有9對,占總對數的32.14%,其中大于0.8的有4對,最大為糙皮樺和刺柏(0.890),其次為糙皮樺和祁連圓柏(0.851)、祁連圓柏和刺柏(0.846)、秦嶺柳和云杉(0.845),但這幾個種對中除祁連圓柏外,其它均為弱勢種。前4個優勢種構成的6個種對生態位重疊值均在0.5以上,其重疊平均值大于后4個弱勢種。
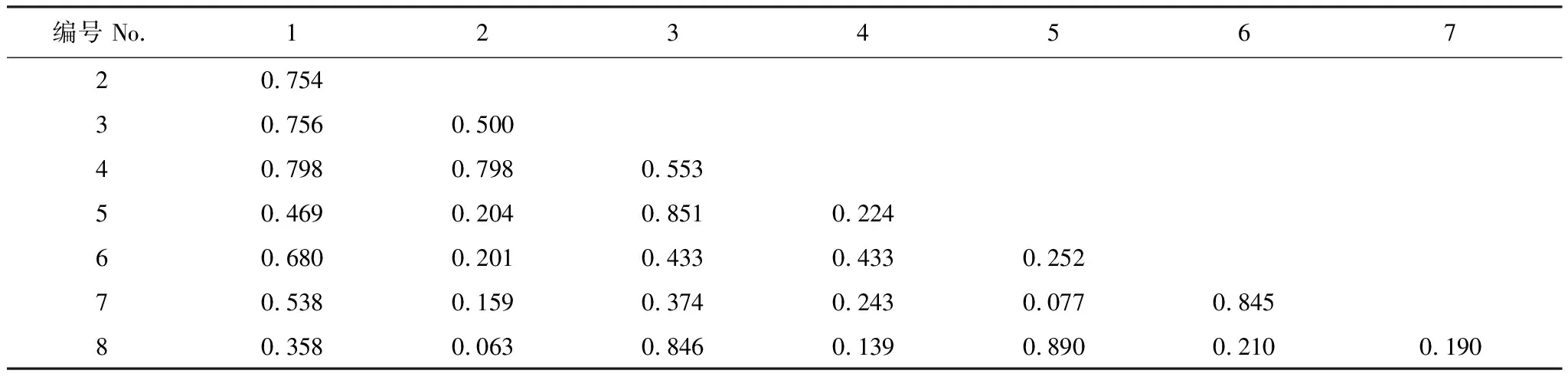
表8 紫果云杉群落喬木層生態位重疊
灌木層1225個種對生態位重疊平均值為0.298,總體重疊程度較低,重疊值小于0.6的有974對,占總對數的79.51%,絕大多數介于0—0.4之間,其中365個種對(占29.80%)重疊值為0,表現出生態位分化現象。重疊值大于0.6的有251對,占比20.49%,其中0.6—0.8的180對,0.8—0.9的34對,0.9—1.0的21對,有16個種對重疊值為1,表現出生態趨同現象。50種灌木僅有峨眉薔薇、唐古特忍冬、藍靛果、華西忍冬和紫果云杉幼苗這5個種與其它所有種均有生態位重疊,其中紫果云杉幼苗與其它種重疊的均值最大,為0.504。前22個種構成的231個種對(表9)生態位重疊均值為0.539,其中重疊值小于0.6的有121對,占比52.38%,未出現重疊值為0或1的種對。0.6以上110對,占比47.62%,其中0.8—0.9的21對,0.9—1.0的6對,重疊值最大的為忍冬和栓翅衛矛(0.923),其次為峨眉薔薇和忍冬(0.923)、峨眉薔薇與扁刺薔薇(0.912)等。
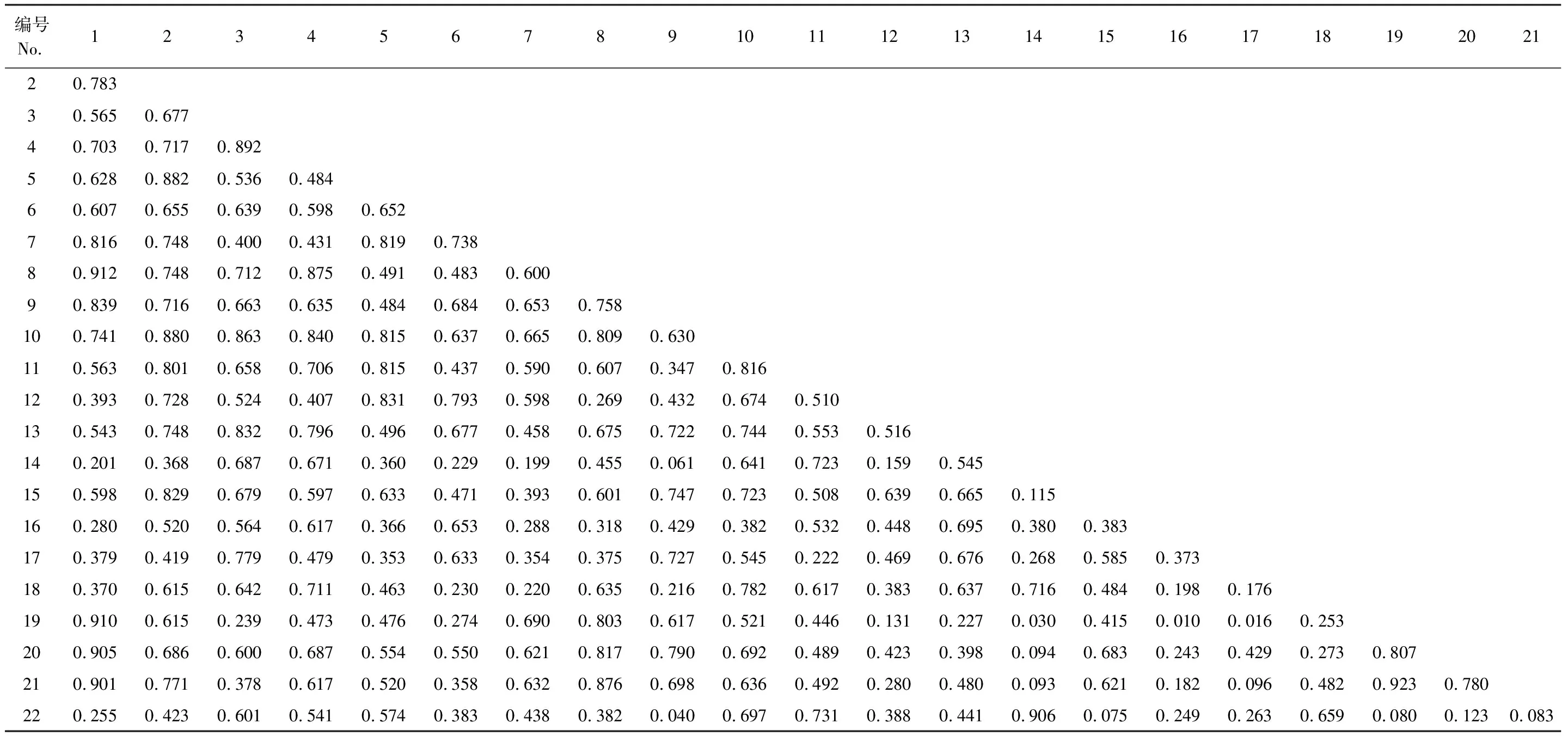
表9 紫果云杉群落灌木層生態位重疊
綜上可知,紫果云杉喬木層普遍存在生態位重疊,且重疊程度大于灌木層。灌木層表現出生態位分化和生態趨同特征,表明物種間可能存在著激烈的競爭。生態位寬度較大的物種之間重疊程度一般較大,且與其它物種重疊的概率高、重疊程度大,非優勢種間、非優勢種與優勢種之間也可見較大的生態位重疊,同科或同屬的物種間生態位重疊往往較大。
3 結論與討論
3.1 物種重要值與生態位寬度
重要值表示植物在群落中的優勢程度[17],生態位寬度表示植物對環境的適應和對資源的利用能力[21]。本研究表明,植物重要值越大生態位寬度也往往越大,但也存在一定差異。在喬木層,紫果云杉重要值和生態位寬度均遠大于其它種,占絕對優勢,但作為次優勢種,岷江冷杉的頻數與生態位寬度卻均小于祁連圓柏和紅樺,資源利用和適應能力均不如二者。灌木層生態位寬度最大的紫果云杉幼苗重要值僅為第9,而重要值最大的峨眉薔薇和唐古特忍冬的生態位寬度則為第12和第4。可見,生態位寬度與重要值無明確相關規律[5],其值大小還與重要值在資源位中分布的均勻程度有關[18],即物種在各資源位分布越均勻,重要值變異系數越小,生態位寬度越大[22]。紫果云杉在灌木層雖無明顯優勢,但所有樣方均有其幼苗存在,且分布均勻變異系數最小(表3),因此生態適應能力最強,而峨眉薔薇、唐古特忍冬等雖然所有樣方也均有出現,但其變異系數較大,因此生態位寬度小于紫果云杉幼苗。
生態位寬度不僅與物種自身生態適應性有關,也與其分布范圍密切相關,并受競爭和環境因子共同影響[5]。在灌木層,重要值和生態位寬度均較大的物種多屬忍冬科和薔薇科,且此二科所有種的重要值總和及生態位總寬度均大于其它所有種之和。大量研究表明,忍冬科植物根系發達生根力強,喜陽,耐陰、耐寒,適應性極強;薔薇科植物多喜光,亦耐半陰,較耐寒,萌蘗性強,適生范圍廣;茶藨子科植物多耐寒,喜濕潤環境。本研究區紫果云杉群落喬木層郁閉度不高,林隙較大,高異質性的林下環境更有利于忍冬科和薔薇科物種的生長。因此,紫果云杉群落灌木層是以藍靛果、蔥皮忍冬、唐古特忍冬等忍冬科植物和以銀露梅、峨眉薔薇等薔薇科植物為主體,茶藨子科及其它科(種)為輔構成的物種多樣和結構穩定的群落。
3.2 生態位相似
生態位相似反映物種對資源利用的相似程度,生態位寬度大的物種之間生態位相似性一般較大[19],但也有研究認為,生態位相似與生態位寬度無明顯關聯[17]。本研究中,喬木層紫果云杉與3個優勢種及其相互之間生態位相似性大于后4個弱勢種,但作為弱勢種,云杉與秦嶺柳、刺柏與糙皮樺的相似性也大于0.6。灌木層紫果云杉幼苗與大多優勢種有較大的生態位相似性,與非優勢種的相似性則普遍較低。相似性較大的藍靛果與蔥皮忍冬、唐古特忍冬與銀露梅等均有較大的生態位寬度,但相似性大于0.8的9個種對有6個種對出現在非優勢種之間、非優勢種與優勢種之間,包括相似性大于0.9的剛毛忍冬與毛花忍冬、花楸與云杉幼苗,可見,生態位寬度較小的物種間也可出現高度的生態位相似性。本研究認為,生態位相似受物種生態特性及生境共同影響,在同一群落相同層次,生態特性相近的物種因資源需求相似常占有相似的生態位,因此,同科或同屬物種間常有較高的生態位相似性,如金露梅與平枝栒子等。
調查研究發現,本研究區紫果云杉群落多分布于中下坡位,并有輕、中度干擾存在。喬木層林木高大,受干擾影響小,灌木層受到干擾時,生態位寬度小的物種常有消失的風險,但由于種間生態位相似性,一個物種的消失會得到與其生態位高度相似的另一物種的補充,因此生態位相似還反映著群落的穩定性[23],而弱勢種或偶見種的消失通常不影響群落結構。紫果云杉幼苗、峨眉薔薇、唐古特忍冬、藍靛果和華西忍冬的生態位寬度大且與所有種均有生態位相似性,因此,任何物種的消失都可能由這5個種來補充,這有利于紫果云杉種群的發展和群落穩定。相似性為0或1的種對多出現在非優勢種之間,表現出生態位相斥和生態趨同現象,這可能是由于物種在長期共存中,競爭及干擾引起了生態位分化和對資源需求的互補或重疊[24],也可能是該研究區并不完全反映所有種的生態位特征。
3.3 生態位重疊
生態位重疊是兩個或多個物種對同一資源或多個資源共同利用的量化表達[21,25],通常生態位寬度大的物種分布均勻,資源利用能力強[6,26],與其它物種重疊概率高、重疊程度大[27]。本研究中紫果云杉與其它7個喬木樹種生態位重疊的均值最大,其次為祁連圓柏與紅樺,但弱勢種秦嶺柳和云杉的生態位重疊值也達0.8以上。在灌木層,僅生態位寬度最大的前5個種與其它所有種有重疊,其中紫果云杉幼苗與其它種重疊的均值最大,其次為唐古特忍冬、華西忍冬等。生態位寬度較大的前12個種與其它種重疊均值(0.356—0.504)大于16個偶見種與其它種重疊的均值(0.117—0.281),但生態位寬度較小的華西薔薇與寶興茶藨子,平枝荀子與金露梅等重疊值也達0.9以上,可見,生態位重疊值大小與生態位寬度無明確的相關規律[28]。本研究認為,弱勢種群適應性弱,易受排擠,再加上生境的異質性,當其在某個適宜生境集中生長時也會出現較大的生態位重疊[17],因此生態位重疊是受物種生態特性和生境共同影響[29]。同科或同屬的物種間由于趨于相同的生活型及相似的生態特性常有較大生態位重疊[30],如扁刺薔薇和峨眉薔薇等。
生態位重疊反映著不同物種在資源共享時存在的競爭及相互促進的關系[31],因此其值的大小并不完全代表種間競爭程度。紫果云杉群落喬木層普遍存在生態位重疊,且重疊均值較大,灌木層前22個優勢種間生態位重疊均值也較大,但所有種的生態位重疊均值僅為0.298,總體重疊程度較低。生態位重疊值越大,預示著占據共同生態空間的物種間潛在的競爭越大[32—33]。調查研究發現,紫果云杉群落喬木層垂直分層明顯,物種多樣性低,數量少,空間資源充足,因此,生態位重疊主要反映物種間的共存狀態與競爭發生可能性,以及具有相似生境需求的物種間的相互促進關系[31],如樺木等喜陽樹種的存在會為紫果云杉的更新提供有利條件。而灌木層植物多達50種,物種多樣性較高,且多在適宜的生境下集中分布,如林窗等,因此,必會產生因資源不足引起的競爭排斥現象,因此生態位重疊在反應物種共存的同時主要反映種間競爭程度,365個種對出現生態位分化及16個種對表現出生態趨同進一步證明了競爭的存在。
經本研究可知,紫果云杉作為建群種占據著群落大部分空間,主導著群落演替,岷江冷杉和祁連圓柏等為伴生種,共同影響著群落生存狀態,樺木等陽性樹種與紫果云杉等針葉樹種形成生態互補和相互促進的關系。紫果云杉、岷江冷杉等幼苗耐陰性強,更新初期需要蔭蔽環境[12],林內廣泛分布的忍冬、薔薇等優勢種(科)在與其幼苗競爭的同時也為其更新提供了有利條件。喬、灌木層共有物種及其分布狀況反映著物種的擴散能力[34],紫果云杉在喬、灌木層均有最大的生態位寬度,生態適應性和資源利用能力最強,具有進一步擴散的可能或正處于擴散階段。
致謝:感謝洮河林區和青海黃南州麥秀林場工作人員給予的幫助。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
