指狀青霉V-ATP酶H亞基蛋白的功能
2022-03-30 02:12:22彭麗桃楊書珍
食品科學 2022年6期
關鍵詞:生長
閆 等,彭麗桃,李 杰,楊書珍
(華中農業大學食品科技學院,湖北 武漢 430070)
柑橘是世界和我國第一大類水果,其種植面積和產量均具世界前列,柑橘產業是南方農村經濟發展的支柱產業[1]。但柑橘果實含水量高,營養豐富,在采后貯運、加工過程中極易遭受病原真菌的侵染而腐爛變質,其中指狀青霉(Penicillium digitatum)的侵染性最強,對柑橘的危害最為嚴重,給生產帶來巨大的損失[2-4]。化學殺菌劑處理是目前生產上控制柑橘采后侵染性病害的主要方法,但隨著化學殺菌劑在生產中長期、大量使用,指狀青霉對化學殺菌劑的抗藥性大幅度提高。由此產生的食品安全、農藥殘留、環境污染等問題嚴重限制了柑橘產業的健康發展[5-7]。因此,研究與開發具有新型、高效、低耐藥性的殺菌劑對柑橘產業的健康發展具有重要意義。
分子生物學的發展開啟了“靶點-高通量篩選-先導物優化”揭示的現代藥物發現模式。作用靶點是研發新型殺菌劑的前提。通過解析生物體生命代謝通路中關鍵蛋白和基因的功能,從中篩選有效作用靶點已經成為研發新型多靶點藥物的重要途徑。pH值穩態系統是維持真菌細胞內環境穩定,保證細胞代謝與生理功能正常的重要前提[8]。pH值穩態系統的破壞顯著抑制白色念珠菌的生長[9],影響球孢白僵菌細胞骨架的維持[10];此外,pH值穩態系統與病原真菌侵染寄主的過程密切相關。越來越多的研究發現,細胞內幾乎所有蛋白質和各細胞器功能的行使、物質代謝、氧化應激、離子穩態、細胞分化、內吞作用以及侵染寄主等生命過程均離不開細胞內pH值的精準調控,是發現新型藥物靶點的重要途徑[11]。V-ATP酶是調控真核細胞pH值穩態的關鍵酶,廣泛分布于真菌和其他真核細胞的液泡、質膜、內質網膜以及高爾基體等內膜系統中,負責將H+從細胞液跨膜運輸到各細胞器中,維持細胞內膜系統pH值穩態[12]。V-ATP酶是一個由多亞基組成的復合物,主要由V1和V0兩個結構域組成。其中,V-ATP酶H亞基蛋白(V-ATPase membrane subunit H,VMAH)主要負責調控復合體V1和V0的組裝以及通過水解ATP調控H+的跨膜運輸[13-14]。釀酒酵母細胞中VMAH的缺失會導致菌體出現致死現象[15]。在球孢白僵菌中VMAH功能缺失會導致菌體細胞pH值穩態系統和Ca2+穩態系統的破壞[10]。前期研究也發現,VMAH是指狀青霉V-ATP酶的關鍵亞基蛋白,在病原菌生長和侵染過程中發揮著重要作用。此外,VMAH亞基蛋白的蛋白結構和氨基酸序列真菌特異性強,作為新型殺菌劑作用靶點具有潛在的研究前景[16],因此,在前期研究的基礎上,進一步利用RNA干擾技術和基因過表達技術,研究VMAH在指狀青霉生長過程中的生理功能以及作為抑制柑橘綠霉病的藥物作用靶點的可行性,為研發適合柑橘采后防腐保鮮的殺菌劑提供支持。
1 材料與方法
1.1 材料與試劑
野生型指狀青霉菌株WT(菌株編號:DSM62840)德國微生物菌種保藏中心;VMAH沉默和過表達指狀青霉突變株si14和oe12,實驗室分別利用RNA干擾技術和過表達技術獲得。
馬鈴薯葡萄糖瓊脂(potato dextrose agar,PDA)培養基:馬鈴薯200 g,加入1 L蒸餾水,煮沸后取濾液加入蔗糖10 g,瓊脂20 g,121 ℃高壓蒸汽滅菌30 min;馬鈴薯葡萄糖液體(potato dextrose broth,PDB)培養基:馬鈴薯200 g,加入1 L蒸餾水,煮沸后取濾液加入蔗糖10 g,121 ℃高壓蒸汽滅菌30 min。
質子泵抑制劑類殺菌劑聯苯芐唑、酮康唑、奧美拉唑、泰妥拉唑、蘭索拉唑、泮托拉唑、巴佛洛霉素A1、依布硒、克霉唑(除克霉唑純度大于99%外,其他純度均大于98%) 上海麥克林生化科技有限公司;NaCl、KCl、CaCl2、CuCl2、FeSO4、MgSO4、H2O2(均為分析純)國藥集團化學試劑有限公司。
1.2 儀器與設備
SPX-150BIII生化培養箱 北京鑫潤科諾儀器儀表有限公司;EX20Thermo Fisher顯微鏡 寧波舜宇儀器有限公司;SHZ-82A氣浴恒溫振蕩器 江蘇省金壇市宏華儀器廠;LGJ-10冷凍干燥機 北京松源華興科技發展有限公司。
1.3 方法
1.3.1 正常生長環境下指狀青霉菌絲生長特性評價
參考菌餅法[17]稍作修改,取在PDA培養基上26 ℃培養3 d的野生、沉默、過表達指狀青霉孢子,制成1×106spores/mL孢子菌懸液,吸取200 μL孢子懸浮液均勻涂布于PDA培養基中,培養至產生白色菌絲后,用7 mm打孔器打取菌餅,將菌餅反貼于PDA培養基,于(26±2)℃的生化培養箱中培養,每隔24 h采用十字交叉法測定病原菌的菌落直徑。
1.3.2 不同酸堿環境下指狀青霉生長特性評價
取在PDA培養基上26 ℃培養3 d的野生、沉默、過表達指狀青霉孢子,制成1×106spores/mL孢子菌懸液,取100 μL孢子懸浮液接種于含50 mL pH值分別為2、4、6、8、10的PDB培養基中,透氣膜封口并搖勻,置于恒溫搖床中(26±2)℃、120 r/min培養48 h。拍照觀察菌絲形態;然后用紗布過濾取菌絲,用蒸餾水沖洗2 次,濾紙吸干表面水分后冷凍干燥機上干燥48 h,分析天平記錄菌絲干質量,菌絲干質量表示菌體生物量。
1.3.3 不同離子脅迫環境下指狀青霉生長特性評價
參考1.3.1節采用菌落反貼的方法研究VMAH沉默對指狀青霉離子脅迫環境適應性的影響。將生長狀態一致的野生型、沉默和過表達指狀青霉的7 mm菌落,分別反貼至含有不同濃度的NaCl、KCl、CaCl2、CuCl2、FeSO4、MgSO4的PDA培養基中,置于26 ℃恒溫培養箱培養并定期觀察。以正常PDA培養為空白對照。每個菌株設置3 個重復,培養7 d,采用十字交叉法測定菌落直徑。按下式計算不同脅迫條件對菌落生長的相對抑制率:

1.3.4 活性氧脅迫環境下指狀青霉生長特性的評價
將生長狀態一致的野生型、沉默和過表達指狀青霉的7 mm菌落,分別反貼至含有不同濃度H2O2的PDA培養基中,置于26 ℃恒溫培養箱培養并定期觀察。以正常PDA培養為空白對照。每個菌株設置3 個重復,培養7 d,采用十字交叉法測定菌落直徑。計算其不同脅迫條件對菌落生長的相對抑制率。
1.3.5 指狀青霉對質子泵抑制劑類殺菌劑的敏感性評價
利用孢子萌發法[18]研究VMAH基因沉默對指狀青霉質子泵抑制劑敏感性的影響。用二甲亞砜分別配制不同質子泵抑制劑的母液,通過二倍稀釋法將藥物稀釋成不同系列濃度,取10 μL各藥液與5 mL 0.5%的PDA培養基混勻后,均勻鋪在載玻片上,培養基凝固后,分別吸取40 μL濃度為1×106spores/mL野生、沉默、過表達指狀青霉孢子懸液滴在載玻片的培養基上,(26±2)℃恒溫培養箱中培養12 h左右,觀察孢子萌發的情況,統計孢子萌發數量,計算孢子萌發率和抑制率,以藥物質量濃度為橫坐標,孢子萌發抑制率為縱坐標,根據藥物毒力回歸方程計算藥物抑菌的半最大效應濃度(the median effective concentration,EC50)[19]。
1.4 數據處理
用Excel軟件進行數據處理和作圖,所有數據均為3 次重復所得,分析采用SPSS 23軟件對實驗數據進行單因素方差分析,應用最小顯著差數法檢驗差異顯著性(P<0.05,差異顯著)。
2 結果與分析
2.1 VMAH沉默和過表達對正常條件下指狀青霉菌絲生長特性的影響
由圖1A可知,與同時期的野生株相比,基因沉默株的生長勢明顯變弱,產孢量顯著降低;而過表達株的生長勢強于野生株,菌落直徑和產孢量均大于野生株。進一步測定菌絲體的生長曲線(圖1B)發現,在整個培養過程中,基因沉默株si14的菌落直徑始終小于野生株;而過表達株oe12的菌落直徑大于野生株。因此,沉默VMAH抑制了指狀青霉的生長,而過表達VMAH促進了指狀青霉的生長。
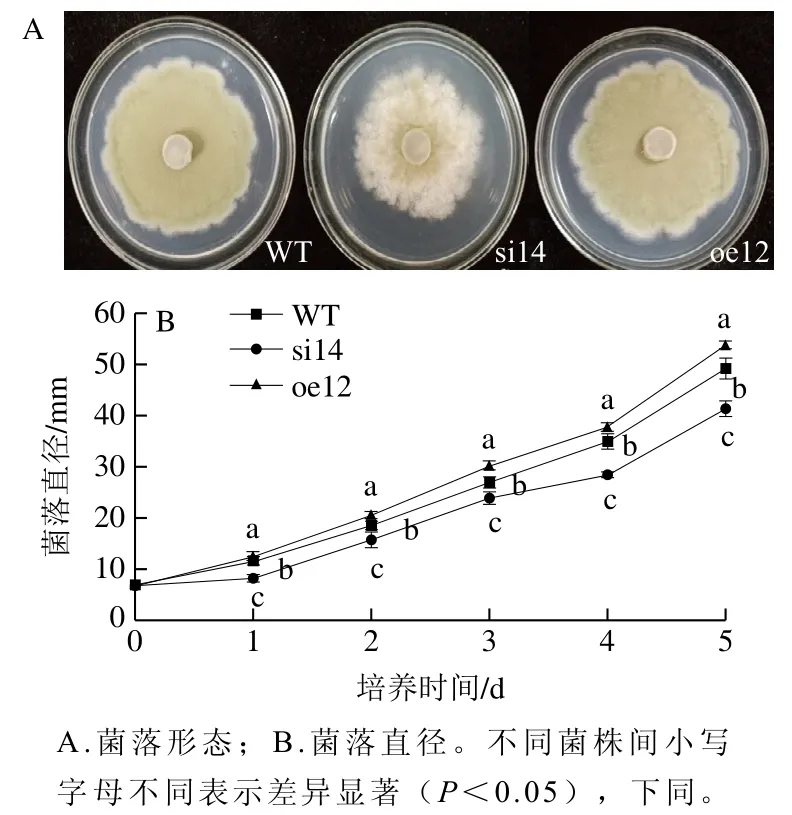
圖1 VMAH基因對指狀青霉菌絲生長的影響Fig. 1 Effect of VMAH gene on the growth of P. digitatum hyphae
2.2 VMAH沉默和過表達對不同pH值環境下指狀青霉生長特性的影響
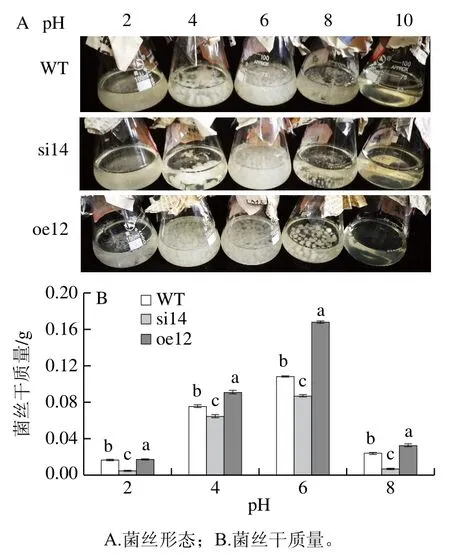
圖2 VMAH基因對指狀青霉pH值適應性的影響Fig. 2 Effect of VMAH gene on pH adaptability of P. digitatum
如圖2所示,WT、si14和oe12的菌絲對偏酸和偏堿條件比較敏感,在pH 10的培養基中,菌絲均不生長。在pH 2的培養基中,過表達和野生菌株都有少量的菌絲生長,無明顯成球的菌絲體,沉默菌株的菌絲量相對更少,菌絲分散。在pH 8的培養基中,WT、si14和oe12都可以看到球狀菌絲,而沉默菌絲球體積較小,呈米粒狀,整體密度不大,分散比較明顯;野生型相對于沉默菌株,其菌絲球體積偏大,在培養基中分布也比較均勻,密度適中;過表達相對于前者,其菌絲球體積最大,而且菌絲球致密且規則。同時,在不同pH值環境下,沉默菌株的生物量顯著低于野生株,在pH 2和pH 8時差異更為顯著;而過表達株的生物量顯著高于野生株,在pH 6時差異最為顯著。
2.3 VMAH沉默和過表達對不同離子脅迫條件下指狀青霉生長特性的影響
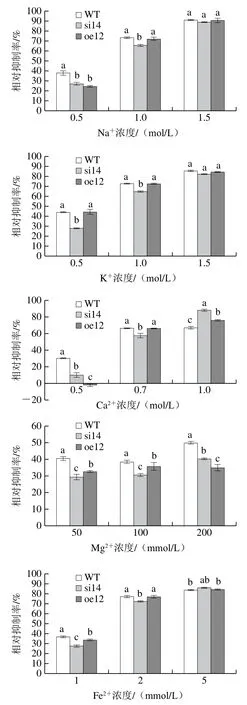
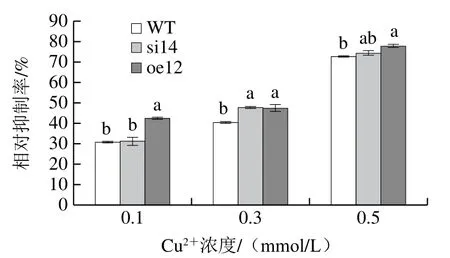
圖3 VMAH基因對指狀青霉離子脅迫環境適應性的影響Fig. 3 Effect of VMAH gene on environmental adaptability of P. digitatum to ion stress
如圖3所示,在6 種金屬離子的脅迫環境下,沉默突變株si14對高濃度的Ca2+、Fe2+、Cu2+比較敏感,對Na+、K+、Mg2+不敏感,其中,沉默突變株對高濃度Ca2+敏感度最高,當Ca2+達到1 mol/L時,其菌絲的相對抑制率可達90%左右,而且抑制率顯著高于野生型和過表達菌株。過表達菌株對高濃度的Cu2+比較敏感,對其他5 種金屬離子不敏感。其中,低濃度的Ca2+對過表達菌株起輕微促進作用。
2.4 VMAH沉默和過表達對指狀青霉H2O2脅迫環境的影響
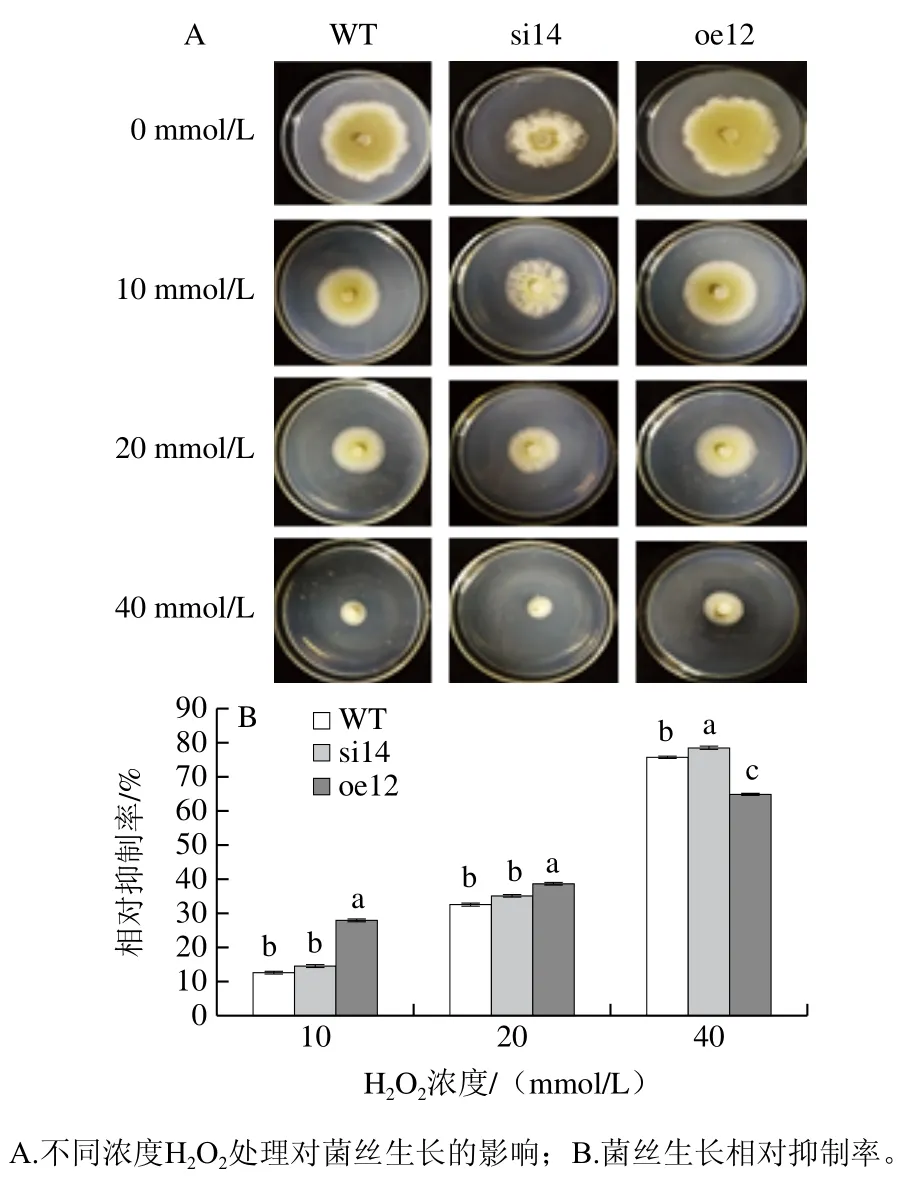
圖4 VMAH基因對指狀青霉H2O2脅迫環境的影響Fig. 4 Effect of VMAH gene on the response of P. digitatum to H2O2 stress
如圖4所示,用不同濃度的H2O2處理后,其生長狀況受到不同程度的抑制,菌落直徑隨著H2O2濃度的增大不斷減小,并且菌絲的產孢量也不斷下降,菌落形態沒有發生明顯變化。從菌絲生長的相對抑制率可以看出低濃度H2O2對野生菌株和沉默菌株的抑制作用較小,無顯著差異,而對過表達菌株的相對抑制率較高。隨著H2O2濃度的增大,菌絲生長的相對抑制率不斷增大,當其濃度為40 mmol/L時,沉默菌株對H2O2更為敏感,其菌絲生長的相對抑制率顯著大于野生株,而過表達菌株的相對抑制率明顯小于野生菌株和沉默菌株。
2.5 VMAH沉默和過表達對指狀青霉質子泵抑制劑敏感性的影響
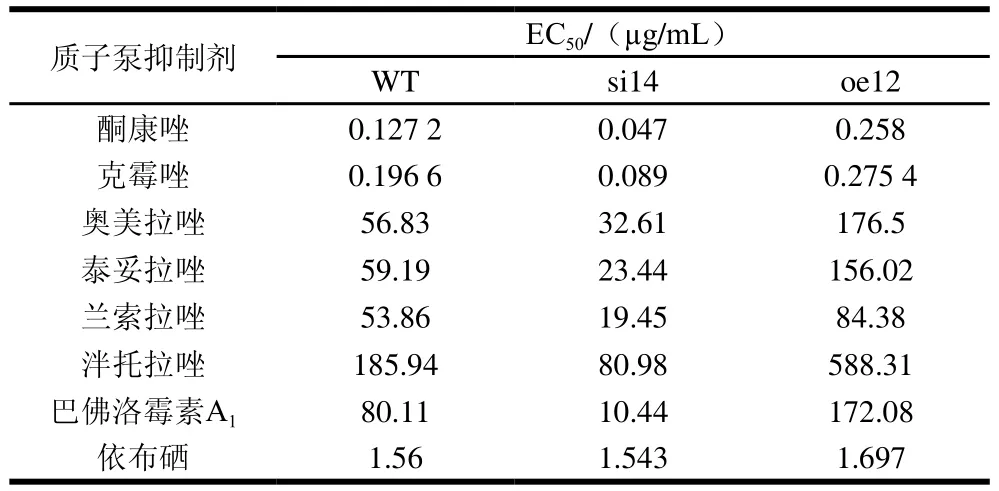
表1 VMAH基因對指狀青霉質子泵抑制劑抑菌EC50的影響Table 1 Effect of VMAH gene on the median effective concentration(EC50) of proton pump inhibitors against P. digitatum
通過孢子萌發法研究VMAH基因沉默后指狀青霉對質子泵抑制劑類殺菌劑的敏感性。本研究選取了包括唑類在內的9 種質子泵抑制劑,由表1可見,相比于野生株,沉默菌株對于除依布硒之外,其他7 種質子泵抑制劑類殺菌劑的敏感性都顯著提高;其中沉默菌株對V-ATP酶的專一抑制劑巴佛洛霉素A1的敏感性提高最為顯著,其EC50為10.44 μg/mL,僅為野生株EC50(80.11 μg/mL)的13%。過表達菌株對質子泵抑制劑的敏感性顯著降低,其中巴佛洛霉素A1的敏感性下降最為顯著,其EC50值為172.08 μg/mL,是野生株的2 倍。因此,沉默VMAH基因顯著提高了指狀青霉質子泵類抑制劑的敏感性,尤其是對巴佛洛霉素A1的敏感增加最多;而過表達VMAH基因顯著降低了指狀青霉對質子泵抑制劑類殺菌劑的敏感性。
3 討論與結論
V-ATP酶在真菌生長發育過程中發揮重要作用。Keenan等[15]在釀酒酵母突變菌株中發現,V-ATP酶中游離V1復合體H亞基的缺失會出現致死現象。絲狀真菌Ashbya gossypii Vma1基因缺失突變株的生長速率下降,并且無法形成生殖孢子[20]。白色念珠菌的Vma5亞基缺失突變株也出現菌絲發育缺陷、生長抑制以及毒力減弱等表征[21]。Lin Meng等[22]發現VMAH功能缺失會影響釀酒酵母液泡內營養物質的貯存和穩定,進而影響酵母絲狀體的形成,導致其生長速率慢于野生型。球孢白僵菌中也有類似發現,VMAH基因缺失的突變株,產孢量下降,產孢時間延長,同時出現相當程度的生長缺陷及致病力的減弱[23]。本研究發現,VMAH基因沉默的轉化子,菌絲生長速率明顯下降,產孢量也顯著減少。因此,VMAH在指狀青霉生長發育過程中發揮著關鍵作用。
V-ATP酶參與調節細胞內的pH值應答,在維持細胞內外pH值平衡中發揮著重要的作用。VMAH基因敲除后會導致球孢白僵菌液泡pH值顯著升高,菌絲生長環境酸化加快,胞內外pH值失衡[23]。光滑念珠菌的VMAHVPH2基因缺失突變株的液泡pH值穩態受損,對堿性條件較為敏感[24]。Martinez-Munoz等[25]研究發現VMAH基因突變會導致酵母菌液泡pH值的顯著增加,pH值穩態遭到破壞。白念珠菌V-ATP酶亞基Vma7p缺失后,菌體液泡酸化度降低,在堿性條件下生長有一定缺陷[26]。而本研究發現,VMAH基因沉默突變顯著降低了指狀青霉在極酸(pH 2)和偏堿(pH 8)條件下的生長力,菌絲生長量顯著減少。因此,VMAH通過調控V-ATP酶活性,在指狀青霉pH值脅迫條件下pH值穩態的維持中發揮重要作用。
V-ATP酶在維持真核生物的離子穩態和氧化應激方面也發揮著一定作用[27-28]。釀酒酵母中V-ATP酶缺失突變株對金屬離子和H2O2比較敏感,與胞質高度酸化和活性氧增加有關[29]。Jia Chang等[30]研究發現白色念珠菌V-ATP酶亞基Tfpl缺失突變株對重金屬離子和高濃度的Ca2+高度敏感,突變株細胞內活性氧水平升高。光滑念珠菌中V-ATP酶組裝相關基因VPH2缺失的突變株對胞外高濃度Ca2+、Na+等離子和氧化劑H2O2的敏感性增加[31]。Zhang Kai等[21]研究也發現,白色念珠菌的Vma5亞基缺失,不僅影響菌絲的形成和降解酶的分泌,還導致鈣穩態紊亂,Ca2+相關的氧化應激反應受到抑制。通過研究指狀青霉VMAH基因沉默突變株對于金屬離子和氧化劑H2O2脅迫的敏感性發現,VMAH沉默突變株對胞外高濃度的Ca2+、Fe2+、Cu2+和氧化劑H2O2的敏感性較高,尤其是高濃度的Ca2+對突變株抑制作用較顯著。推測突變體對金屬離子的高度敏感性與維持離子穩態的缺陷有關,這些金屬離子穩態失衡可能進一步引起內源氧化應激,從而對氧化劑H2O2比較敏感,表明VMAH通過調控V-ATP酶活性在維持細胞離子穩態、活性氧水平和氧化應激方面起重要的作用。
研究發現環境pH值影響病原菌對抗生素的敏感性[32]。編碼光滑念珠菌V-ATP酶組裝的VPH2基因的缺失,會增強對抗真菌劑(唑類和兩性霉素B)的敏感性[24]。Zhang Yongqiang等[33]發現抗真菌藥物的唑類化合物通過抑制細胞膜脂組分甾醇的合成和V-ATP酶的活性進而引起菌體細胞的死亡。位于細胞膜的P-ATP酶亞基Spfl的缺失會提高菌體對氟康唑、衣霉素和潮霉素B敏感性[34]。酵母菌V-ATP酶亞基a的突變體對巴佛洛霉素具有抗性[35]。本研究發現VMAH基因沉默對導致菌體對質子泵抑制劑類殺菌劑尤其是V-ATP酶的專一性抑制劑巴佛洛霉素A1的敏感性提高,而過表達突變株對質子泵抑制劑的敏感性降低。因此,VMAH與質子泵類殺菌劑具有協同殺菌作用,是多靶點殺菌劑研發的潛在藥物靶點。
綜上所述,VMAH在指狀青霉的生長、pH值和離子穩態系統的維持、活性氧水平和氧化應激方面發揮著重要作用,作為控制柑橘采后綠霉病殺菌劑的作用靶點值得進一步深入研究。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
