基于同質園實驗的兩種典型荒漠植物葉片功能性狀變異研究
2022-03-28 04:55:34單立山解婷婷種培芳師亞婷
草地學報 2022年3期
馬 麗, 單立山, 解婷婷, 種培芳, 楊 潔, 師亞婷
(甘肅農業大學林學院, 甘肅 蘭州 730070)
植物功能性狀是指植物在個體水平上所表現出的植物形態、生理等特征,能夠有效表達植物在長期進化過程中對外界環境變化的響應[1-2]。葉片作為植物地上部分的主要器官,是植物進行光合作用和物質生產的主要場所[3],其功能性狀如葉片含水量、比葉面積、葉片碳氮磷含量和解剖結構等最能體現植物對環境因子的適應性進化[4-6]。目前,植物功能性狀研究已成為植物功能生態學研究中的前沿和熱點問題之一[7-8]。雖有大量研究報道了植物葉功能性狀變異及其與環境因子的關系[9-11],但并不能說明植物葉功能性狀是受環境影響還是遺傳因素的影響,而同質園實驗能夠在一定程度上控制環境因素所造成的影響,進而說明不同種源植物葉功能性狀的變異是由于原種源地環境差異造成的植物遺傳性基因變異。因此,基于同質園實驗探究植物葉功能性狀的變異及其與環境因子的關系具有重要意義,有助于我們更好地理解不同種源地植物對環境梯度變化的響應和適應機制。
植物葉片通過改變自身的形態和化學元素來適應環境變化[12-13]。目前,關于植物葉功能性狀受環境因子影響的研究主要集中在全球等大尺度層面[14],但這種研究會忽略一些小尺度層面的環境因子如微地形、生境等,對這些環境因子的研究也許能夠揭示大尺度下所無法解釋的環境過濾效應[15]。植物為適應環境會形成以該環境為主的特定地理種源,遺傳變異就是植物為適應該環境變化的適應結果,而由于短期環境因子如土壤質量等引起的植物葉功能性狀的變異是植物適應短期環境變化的結果[16]。此外,在以往試驗中沿自然環境梯度采樣時,很難分辨植物表現型、基因型和環境之間的關系,如黃河上游的寧夏平原和黃河下游的黃河三角洲地區的蘆葦(Phragmitesaustralis)植物葉功能性狀的差異同時受到環境和遺傳兩個因素的共同影響[17];全緣葉綠絨蒿(Meconopsisintegrifolia)隨著海拔升高,株型變矮,葉片組織結構緊密度增大[18]。然而,同質園實驗將不同生境的物種栽培在環境因素相對一致的同一園地內,進行統一栽培管理,使其生境保持一致;該方法能夠減少立地條件和氣候變化造成的影響,在局部適應性研究中發揮重要作用[19]。近年來,雖然有關同質園的生態學研究已有諸多的報道,但主要集中在森林植物葉功能性狀變異和對環境的適應性研究等方面[20-21],而關于荒漠植物在其葉功能性狀變異及影響機制方面的研究則較少。
白刺(Nitrariatangutorum)和紅砂(Reaumuriasoongarica)是半干旱、干旱地區常見的鹽生、旱生荒漠植物,具有抗旱、抗寒、耐高溫和耐鹽堿等特性以及防風固沙、水土保持等作用[22-23]。白刺和紅砂廣泛分布于我國西北荒漠地區,并在當地形成了以它們為建群種或優勢種的荒漠群落[23]。以往對白刺和紅砂的研究多集中在自然環境梯度下葉片形態解剖、根系的形態和功能、對極端環境的適應性及群落功能等方面[24-27],但是采用同質園實驗方法對不同種源兩種典型荒漠植物葉功能性狀的研究相對較少。因此,本研究分析了不同地理種源兩種典型荒漠植物葉片的肉質化程度(Degree of fleshiness,DOF)、含水量(Water content,WC)、組織密度(Tissue density,TD)、比葉面積(Specific leaf area,SLA)、碳(Carbon,C)含量、氮(Nitrogen,N)含量、磷(Phosphorus,P)含量、碳氮比(Carbon and nitrogen ratio,C∶N)、碳磷比(Carbon and phosphorus,C∶P)和氮磷比(Nitrogen and phosphorus ratio,N∶P)指標,旨在探討不同地理種源兩種典型荒漠植物葉功能性狀在種源間是否存在差異以及其與種源環境因子的關系,以期為兩種典型荒漠植物選擇優良種源、適宜的種植環境及兩種典型荒漠植物對環境適應策略等提供理論依據。
1 材料與方法
1.1 研究地概況
同質園位于甘肅省武威市涼州區羊下壩鎮甘肅武威市良種繁育中心(38°24′ N,103°9′ E),地處河西走廊北部,屬于典型的溫帶大陸性荒漠氣候,干旱少雨,土壤性質為典型砂壤土,pH值為8.2,有機碳含量為14.30 g·kg-1,全氮含量為0.68 g·kg-1,全磷含量為1.42 g·kg-1,速效鉀含量為152.6 mg·kg-1,0~20 cm土壤容重為1.57 g·cm-3,平均土壤含水量0.4%,海拔為1 378 m,年均降雨量為113.2 mm,年均蒸發量為2 604.3 mm,年均氣溫為6.9℃,7月平均溫為24.9℃,1月平均氣溫為—9.9℃,日最高氣溫為35.9℃,日最低氣溫為—25.1℃[28-29]。
1.2 試驗材料的培養及選取
2011年在白刺和紅砂的成熟季節,沿自然環境降水梯度在我國甘肅酒泉(JQ)、張掖(ZY)、武威(WW)和蘭州(LZ)天然分布區分別選取每個種源中生長良好、無病蟲害的多個優勢單株,采集種子,并進行標記裝袋;2012年1月在武威市涼州區羊下壩鎮甘肅武威市良種繁育中心苗圃溫室進行穴盤育苗,基質成分為沙土10%~20%,砂壤土60%~80%,腐熟羊糞10%~20%,播種后進行日常的苗木管理。2013年4月,土壤解凍后,進行移栽,建立白刺和紅砂種源試驗地。移栽時采用隨機區組試驗設計,株行距0.5 m×1.5 m。移栽后隨即灌水,保證苗木的成活率。待苗木成活后,適時除草,按日常管理方法對苗木進行管理[29]。
2020年在苗木生長旺季,以同質園內定植的4個種源地的白刺和紅砂作為研究對象(4個種源地的地理位置與環境因子見表1[30]),分別選取每個種源中生長健壯、長勢良好、無病蟲害的10個成年植株,剪取每個植株直立莖中部的葉片放入打濕的信封袋中,并且裝入封口塑料袋內封口,立即帶回實驗室用于葉片功能指標測定。
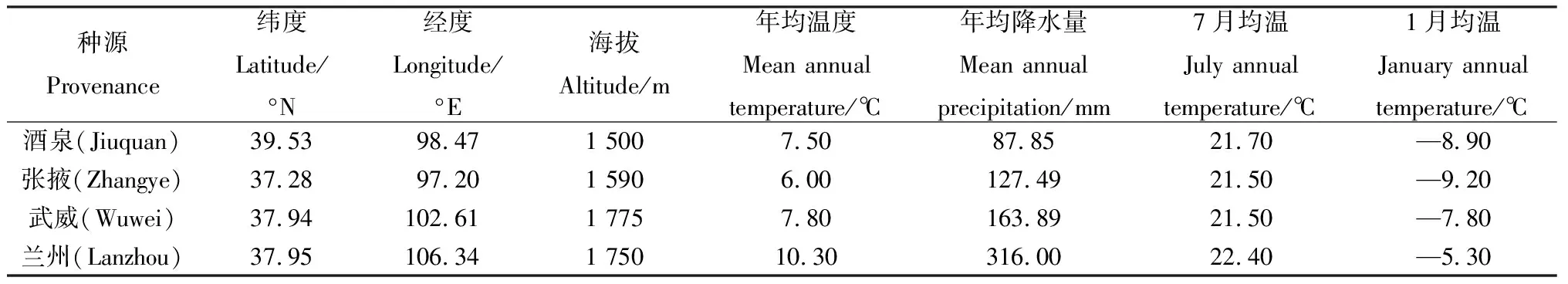
表1 白刺和紅砂4個種源地的地理位置與環境因子Table 1 Geographical location and environmental factors of the four provenances of N. tangutorum and R. soongarica
1.3 測定方法
1.3.1葉片形態性狀的測定 將采集葉片上的雜質清理掉,稱量葉鮮重,再將葉片用根系分析軟件(Win-RhIZO 2008a)測量葉片表面積、體積等形態特征。測量完成之后,將葉片置于75℃烘箱烘至恒重,稱量干重并記錄。每種源各有3次重復。白刺和紅砂葉片的肉質化程度、含水量、比葉面積和組織密度的計算公式如下:

(1)
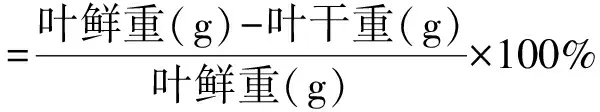
(2)

(3)

(4)
1.3.2葉片碳(C)、氮(N)和磷(P)含量的測定 將以上測定完的植物樣品用碾槽碾碎,裝進密封袋用于進行葉片C,N,P含量的測定。C含量測定采用重鉻酸鉀氧化加熱法;N含量測定采用凱氏定氮法;P含量測定采用釩鉬黃比色法測定[31]。
1.4 氣象數據來源
本研究選擇了對植物葉功能性狀具有影響的環境因子。氣象數據以酒泉、張掖、武威和蘭州4個種源地的數據為主(http://www.weather.com.cn/)。通過對數據的整理,計算出每個種源附近各站點的經度(Longitude,Lng)、緯度(Latitude,Lat)、海拔(Altitude,ALT)、年均溫度(Mean annual temperature,MAT)、年均降水量(Mean annual precipitation,MAP)、1月均溫(January annual temperature,JaT)、7月均溫(July annual temperature,JuT)等數據,作為本次研究的氣象數據。
1.5 數據處理
本試驗所有數據統計分析和相關作圖均是使用Excel 2016,SPSS 24.0(America),Origin 2018(America)和Canoco5.0(America)軟件完成。采用單因素方差分析法分析同一物種在不同種源間葉片各性狀差異顯著性,并用Duncan檢驗法和最小顯著差異法(LSD)進行多重比較;用Pearson相關系數檢驗白刺和紅砂各葉功能性狀之間的相關性。采用冗余分析(RDA)法和逐步線性回歸法分析白刺和紅砂葉功能性狀對不同種源地環境因子的響應關系;在RDA分析之前,先對葉功能性狀進行去趨勢對應分析(DCA),確定白刺和紅砂是采用以線性模型排序的冗余分析(RDA)還是以單峰模型排序的典型對應分析(CCA)。若最大排序軸長小于3,則適合線性模型排序分析的RDA方法;若最大排序軸大于4,則適合單峰模型排序的CCA方法;若最大排序軸長在3~4之間,則兩種方法都可以。本研究中最大排序軸長為0.1,小于3,故采用RDA方法分析,且將種源地環境因子作為解釋變量(空心箭頭),將葉功能性狀作為響應變量(實心箭頭)。文中數據均為平均值±標準誤差。
2 結果與分析
2.1 白刺和紅砂葉片形態性狀的種源間差異
由圖1可知,白刺葉片肉質化程度在JQ種源與其他地理種源差異顯著(P<0.05),且在JQ種源最小(3.99),與ZY種源、WW種源和LZ種源相比,JQ種源白刺葉片肉質化程度減小8.62%,14.36%和10.95%;含水量在JQ種源與其他地理種源差異顯著(P<0.05),且在JQ種源最小(74.88),與ZY種源、WW種源和LZ種源相比,JQ種源白刺葉片含水量減小2.56%,4.47%和3.22%;組織密度在種源間差異顯著(P<0.05),且在ZY種源最大(0.08 g·cm-3),在WW種源最小(0.06 g·cm-3),與WW種源相比,ZY種源白刺葉片組織密度顯著增加了30.79%。
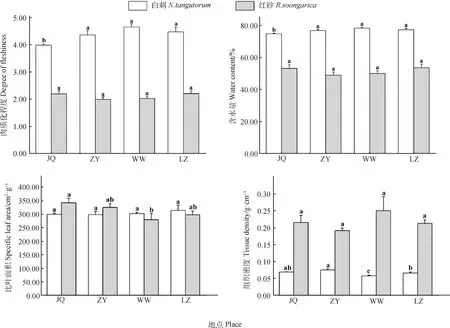
圖1 不同地理種源白刺和紅砂葉片形態結構Fig.1 Leaf morphological structure of N. tangutorum and R. soongarica from different geographic provenances注:JQ-酒泉、ZY-張掖、WW-武威、LZ-蘭州;不同小寫字母表示同一物種在不同種源間差異顯著(P<0.05)。下同Note:JQ-Jiuquan,ZY-Zhangye,WW-Wuwei,LZ-Lanzhou;different lowercase letters indicate significant difference between different provenances under the same species at the 0.05 level. The same as below
紅砂葉片的比葉面積在JQ和WW種源間差異顯著(P<0.05),與WW種源相比,JQ種源的紅砂葉片的比葉面積增加了22.24%;肉質化程度、含水量和組織密度在各個種源均無顯著差異。
2.2 白刺和紅砂葉片C,N,P含量及其計量比的種源間差異
由圖2可知,白刺葉片N含量在種源間差異顯著(P<0.05),在JQ種源最大,為3.79 g·kg-1,在LZ種源最小,為3.31 g·kg-1;葉片N∶P在JQ種源與其他種源間均差異顯著(P<0.05),且在JQ種源比值最大,為10.37,與ZY種源、WW種源和LZ種源相比,JQ種源葉片N∶P增加了47.95%,24.72%和63.72%。
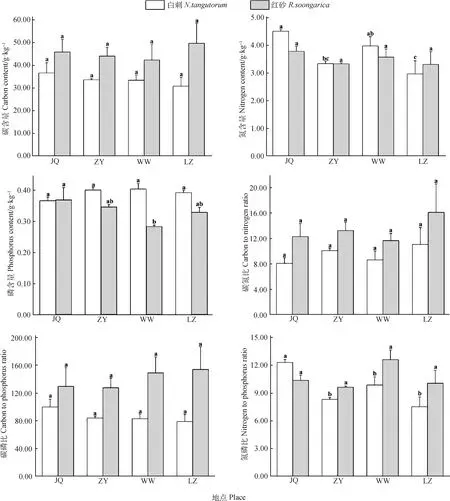
圖2 不同地理種源白刺和紅砂葉片C,N,P含量及其計量比Fig.2 Contents of C,N,and P in leaves of N. tangutorum and R. soongarica and their stoichiometric ratio from different geographic provenances
紅砂葉片P含量在JQ和WW種源間差異顯著(P<0.05),在JQ種源最大,為0.37 g·kg-1,在WW種源最小,為0.28 g·kg-1,與WW種源相比,JQ種源紅砂葉片P含量增加了30.40%。
二者的C含量、C∶N和C∶P在各個種源均無顯著差異。
2.3 白刺和紅砂葉片功能性狀間的相關性
通過Pearson相關性分析得出(表2),白刺葉片的組織密度與肉質化程度、含水量和比葉面積呈極顯著負相關關系(P<0.01);N含量與比葉面積呈顯著負相關關系(P<0.05),C∶P與C含量呈極顯著正相關關系(P<0.01);N∶P與N含量呈極顯著正相關關系(P<0.01),與C∶N呈顯著負相關關系(P<0.05)。通過Pearson相關性分析可以看出(表3),紅砂葉片的組織密度與比葉面積呈極顯著負相關關系(P<0.01);C∶N與C含量呈極顯著正相關關系(P<0.01),與N含量呈顯著負相關關系(P<0.05);C∶P與C含量呈極顯著正相關關系(P<0.01),與P含量呈顯著負相關關系(P<0.05),與C∶N呈極顯著正相關關系(P<0.01);N∶P與N含量呈顯著正相關關系(P<0.05)。
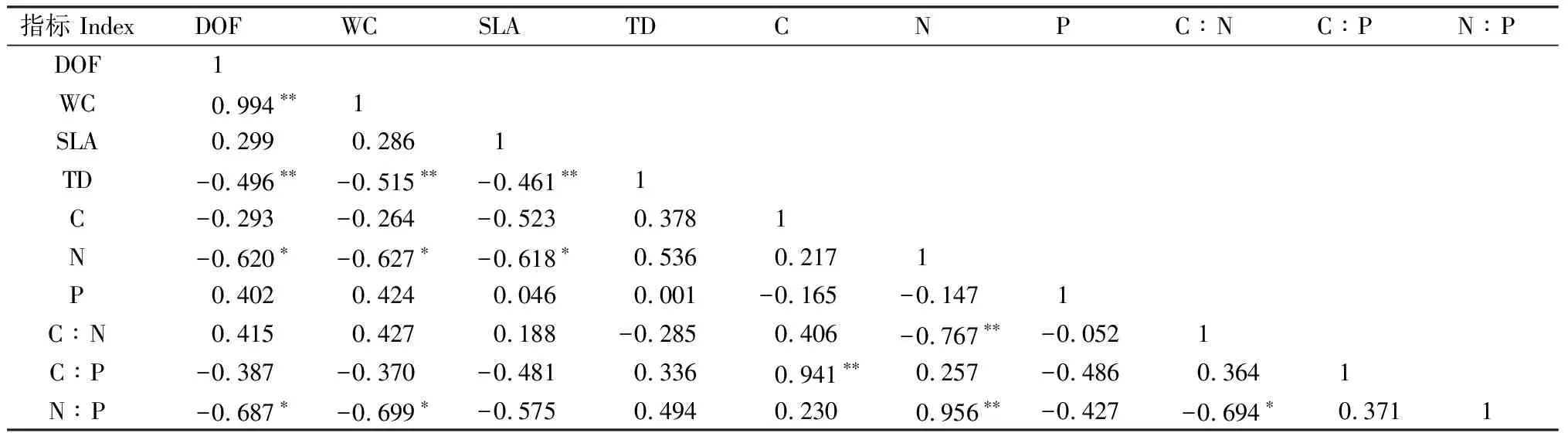
表2 不同地理種源白刺葉片功能性狀間的相關性Table 2 Correlation among leaf functional traits of N. tangutorum from different geographic provenances
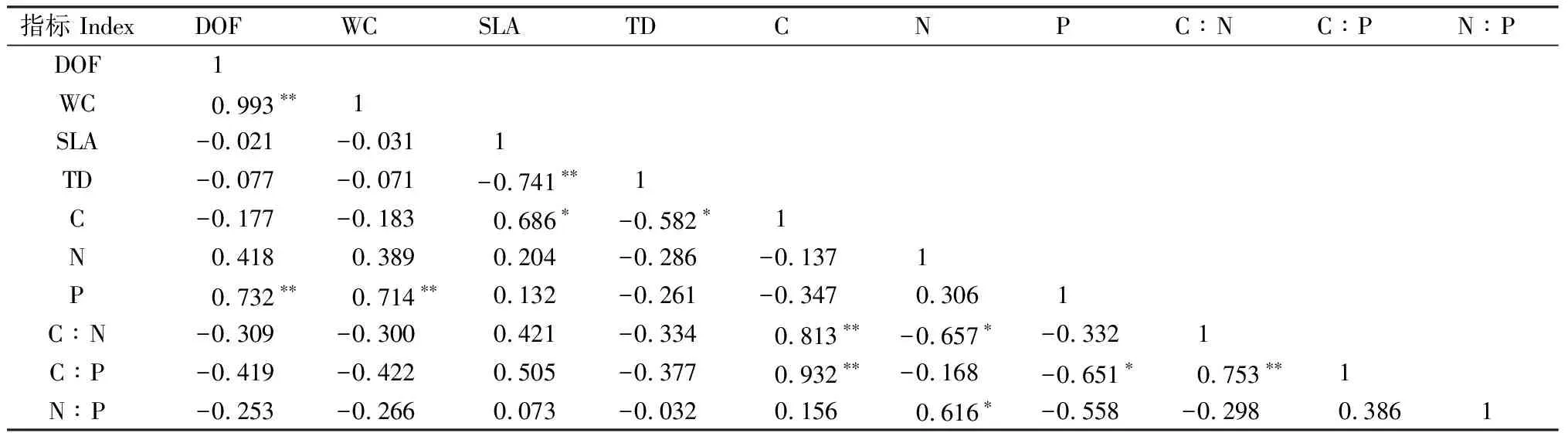
表3 不同地理種源紅砂葉片功能性狀間的相關性Table 3 Correlation among leaf functional traits of R. soongarica from different geographic provenances
2.4 不同地理種源白刺和紅砂葉片功能性狀與種源地環境因子的關系
2.4.1葉片功能性狀與種源地環境因子的冗余分析 通過RDA分析(表4),第1,2軸的白刺葉功能性狀與環境的相關系數分別為0.9163,0.9213,特征值分別為0.3265,0.1574,共解釋了48.40%的葉功能性狀和92.37%的葉功能性狀與種源地環境的關系,表明前兩個排序軸能夠較好地反映白刺葉功能性狀與種源地環境因子的關系。蒙特卡羅檢驗第1排序軸和所有排序軸均達到極顯著水平(P<0.01),表明結果具有統計學意義。由圖3可知,海拔與肉質化程度、含水量呈正相關關系,與組織密度呈負相關關系;緯度與N∶P呈正相關關系,與P含量呈負相關關系;年均降水量與C含量、N含量、C∶P呈負相關關系。RDA分析(表5)表明,第1,2軸的特征值分別為0.804 6,0.006 9,共解釋了81.16%的紅砂葉功能性狀和99.69%的葉功能性狀與種源地環境的關系,表明前兩個排序軸能夠較好地反映紅砂葉功能性狀與種源地環境因子的關系。蒙特卡羅檢驗第1排序軸和所有排序軸均達到極顯著水平(P<0.01),表明結果具有統計學意義。由圖4可知,海拔、年降水量和年均溫度與第1排序軸呈正相關關系,且相關性依次為海拔>年降水量>年均溫度;海拔與紅砂比葉面積、P含量呈負相關關系。
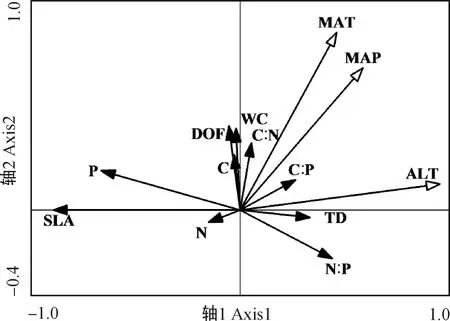
圖4 紅砂葉功能性狀與種源地環境因子的RDA排序Fig.4 Redundancy analysis ordination of leaf functional traits of R. soongarica and geographic environmental factors
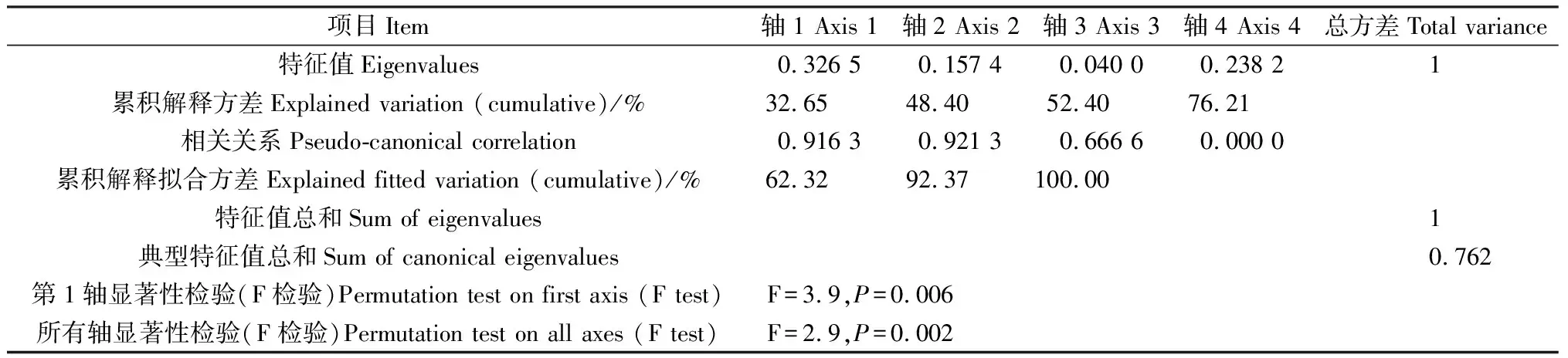
表4 白刺葉性狀與環境因子的RDA排序Table 4 Redundancy analysis ordination of leaf traits and environmental factors of N.tangutorum
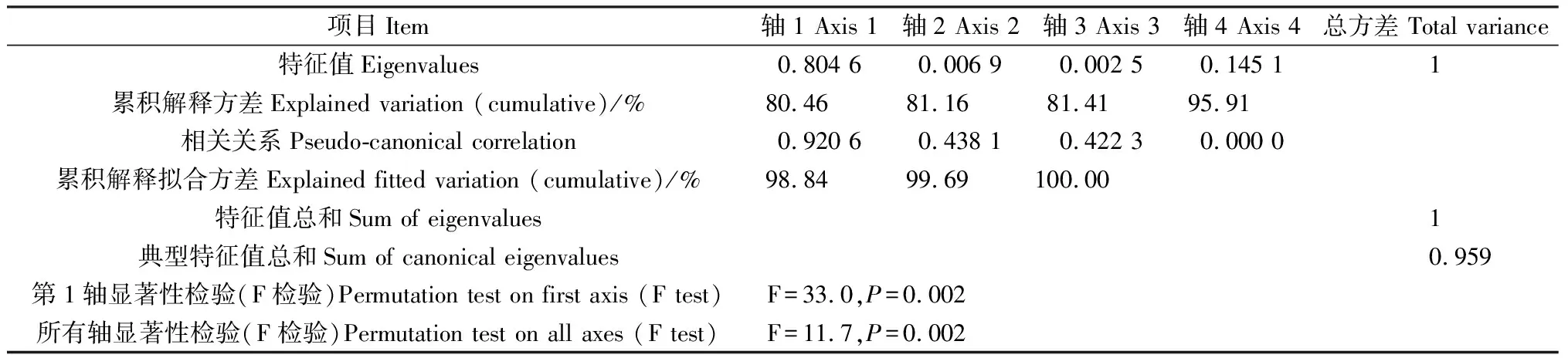
表5 紅砂葉性狀與環境因子的RDA排序Table 5 Redundancy analysis ordination of leaf traits and environmental factors of R. soongarica
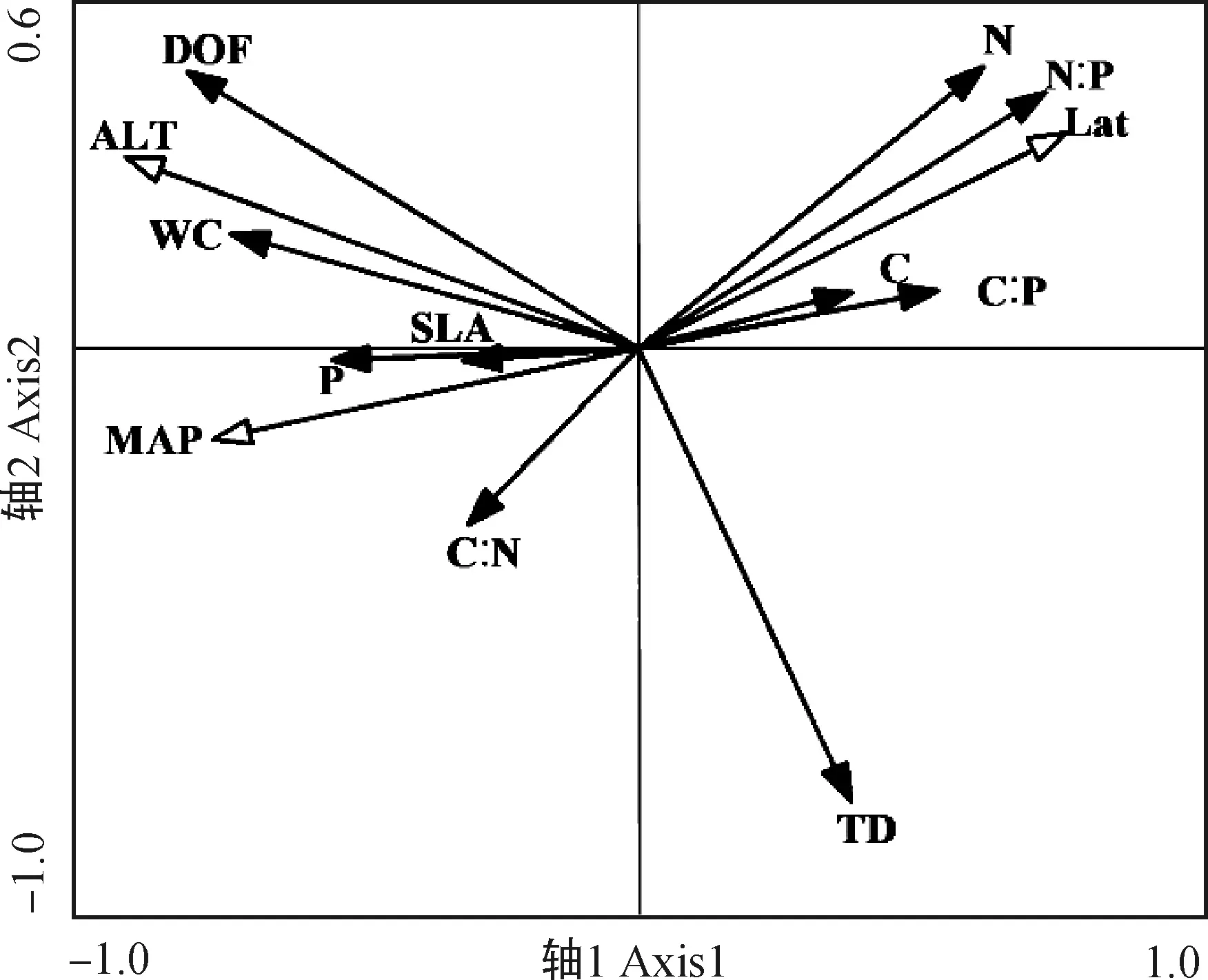
圖3 白刺葉功能性狀與種源地環境因子的RDA排序Fig.3 Redundancy analysis ordination of leaf functional traits of N. tangutorum and geographic environmental factors注:經度-Lng,緯度-Lat,海拔-ALT,年均溫度-MAT,年均降水量-MAP,下同Note:Longitude-Lng,Latitude-Lat,Altitude-ALT,Annual average temperature-MAT,Annual average precipitation-MAP,the same as below
2.4.2葉功能性狀與種源地環境因子的逐步線性回歸分析 由表6可知,白刺葉片肉質化程度、含水量與海拔呈極顯著正相關關系(P<0.01);葉組織密度與海拔、緯度均呈極顯著負相關關系(P<0.01),且海拔的標準化回歸系數的絕對值(|—0.697|)大于緯度的標準化回歸系數的絕對值(|—0.441|),表明影響葉組織密度的主要環境因子是海拔,其次是緯度;葉片N含量與年均降水量呈顯著負相關關系(P<0.05),與1月均溫呈顯著正相關關系(P<0.05),且年均降水量的標準化回歸系數的絕對值(|—2.704|)大于1月均溫的標準化回歸系數(2.126),表明影響葉片N含量的主要環境因子是年均降水量,其次是1月均溫;葉片P含量與緯度呈顯著負相關關系(P<0.05),葉片N∶P與緯度呈顯著正相關關系(P<0.05),與7月均溫呈顯著負相關關系(P<0.05),且緯度的標準化回歸系數(0.708)大于7月均溫的標準化回歸系數的絕對值(|-0.441|),這表明影響葉片N∶P的主要環境因子是緯度,其次是7月均溫。因此,白刺葉片肉質化程度、含水量、組織密度主要受海拔的影響,葉片N含量主要受年均降水量的影響,葉片P含量、N∶P主要受緯度的影響。而紅砂葉片的比葉面積和P含量主要受海拔影響。
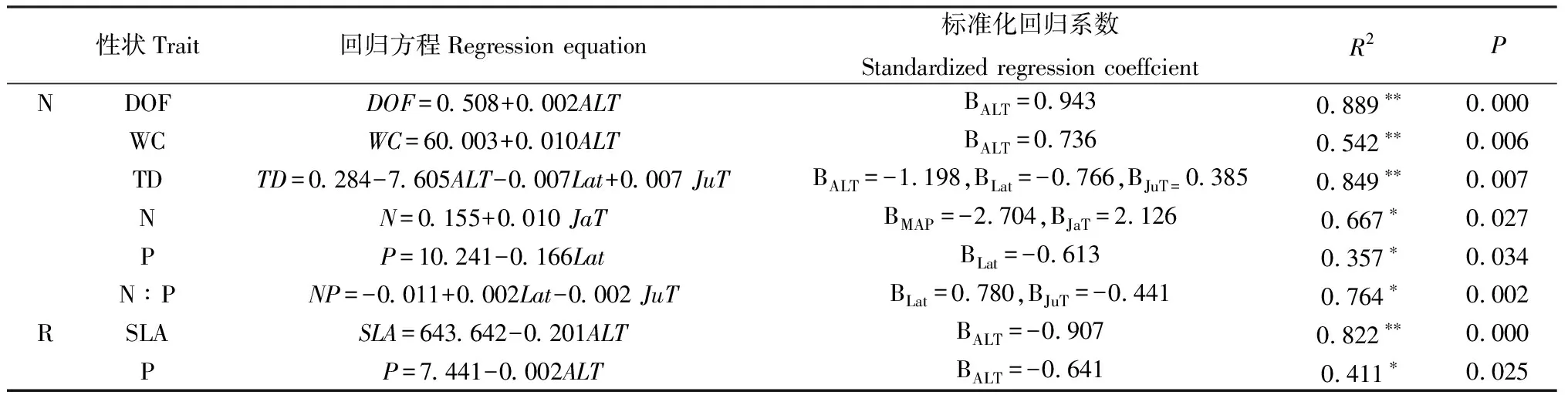
表6 白刺和紅砂葉功能性狀與環境因子的逐步回歸分析Table 6 Stepwise regression analysis of leaf functional traits and environmental factors of N. tangutorum and R. soongarica
3 討論
3.1 不同地理種源白刺和紅砂葉片形態性狀的變異特征
植物在長期適應種源環境下,必然會產生生理生態特性的分化,形成不同的地理種源,導致同一葉功能性狀在不同種源間差異顯著[32]。大量研究表明,植物長期受種源地環境影響,導致其葉功能性狀產生遺傳分化[33-35]。本研究結果也顯示,不同地理種源白刺在同質園環境中葉片肉質化程度、含水量、組織密度表現出顯著性差異(圖1),表明白刺受種源地環境的影響,其葉功能性狀差異可能與遺傳分化有關。有研究表明,在野外環境和同質園環境中,植物由于受種源環境的影響,產生了遺傳穩定性[17]。楊彪生等[36]研究發現,野外環境下白刺葉片肉化程度、含水量和組織密度均呈現顯著性差異,這一野外研究結果與本研究中同質園實驗的研究結果一致,說明白刺葉功能性狀通過對種源環境的適應可能形成了遺傳穩定性。楊彪生等[36]對野外環境中紅砂葉片肉質化程度、含水量和組織密度的研究發現這些指標均呈顯著性差異。然而,本研究中同質園內不同地理種源紅砂葉片的肉質化程度、含水量、組織密度均無顯著性差異(圖1),說明紅砂在同質園環境中不受種源環境的影響。
比葉面積可以反映植物對資源的獲取能力,且也能夠反映出植物對不同環境的適應性特征,是研究植物功能性狀的重要指標之一[37-38]。有研究認為,植物在資源豐富的環境中生長比葉面積較大,在資源匱乏的環境中生長往往比葉面積較小[39]。在本研究中,同質園中紅砂的比葉面積(311.26 cm2·g-1)高于野外環境中紅砂的比葉面積(277.32 cm2·g-1),這可能是同質園環境有較充足的水分、光照等條件,與野外環境相比,同質園中比葉面積利用光合的能力較強,能夠快速獲取所需資源。
3.2 不同地理種源白刺和紅砂葉片C,N,P含量化學計量及其計量比的變異特征
葉片C,N,P含量對異質環境的適應是表型分化和遺傳分化共同的結果,并在長期的進化過程中形成不同的生理生態和養分利用特征,若在同質園環境中,這種生理生態的分化依然存在,則認為植物為了適應環境產生了遺傳分化[17]。有研究表明,同質園試驗植物葉片的C,N,P含量及其計量比在種源間表現出顯著性差異,說明植物葉片中化學計量特征在對種源地環境條件的長期進化適應過程中產生了遺傳分化[40-41];本研究發現白刺葉片N含量,N∶P在不同地理種源間表現出的結果與上述結果相同(圖2),說明白刺受種源環境的影響可能使其葉片化學元素產生了遺傳分化。不同種源翅莢木(Zeniainsignis)葉片C含量、N含量及C∶N存在差異顯著,說明其主要葉片養分性狀可能主要受長期的種源地環境影響產生了遺傳分化[32],這與本文研究中紅砂葉片C含量、N含量及C∶N無顯著差異的結論相悖(圖2),這可能是因為翅莢木研究地屬于中亞熱帶海洋性季風氣候,雨水充沛,氣候溫暖,而本文研究地屬于干旱性大陸氣候,干旱少雨,兩個研究地的立地條件和氣候環境截然不同,所以對其葉功能性狀的影響程度也不同。
C,N,P含量化學計量比對植物營養元素的平衡起限制性作用,并已經廣泛運用于植物養分循環和限制的研究[42-43]。N∶P常被看作植物在生長過程中獲取環境養分的限制性因素[1,44]。有研究表明,植物生長發育受N和P限制的N∶P比值為14和16,即當N∶P<14時,植物生長發育主要受N限制;當N∶P>16時,植物生長發育主要受P限制;當14 植物葉片在長期適應地理環境的過程中,通過內部不同功能間的協調,形成一系列適應環境的葉功能性狀間的不同組合,并非葉片性狀孤立發揮作用[47-48]。通常認為,植物葉片的比葉面積與組織密度呈負相關的關系,即比葉面積增大時,組織密度會變小。比葉面積和組織密度作為植物資源分配利用的重要指標,均能夠反映植物對環境的適應策略(水分利用策略)[49-51]。本研究發現,白刺和紅砂的比葉面積與組織密度呈極顯著負相關關系(表2、表3),出現這種現象可能與植物地處干旱少雨的環境密切有關,即當植物處于干旱環境時,植物為了適應干旱環境,從周圍環境中不斷汲取養分,使得比葉面積增大,但此過程中植物對光照需求較大,而對葉片中儲存的養分和水分的需求較小,故養分資源的利用效率相對較低,因此植物葉片相對較薄,葉組織密度相對較小。因此,白刺和紅砂葉片通過比葉面積和組織密度的平衡形成對不同水分環境的適應策略[47,52]。有研究表明,作為植物生長中主要的營養元素,N和P具有一定的相互促進作用,N∶P可以反映N和P兩種營養元素供給狀況的相對有效性[53]。白刺和紅砂葉片N含量與N∶P呈顯著正相關的關系(表2、表3),這是由于植物對養分的利用受到了環境和自身需求的影響,植物通過調節自身葉片營養元素及化學計量比來適應環境中養分的供給情況[41]。葉片C∶N和C∶P表示植物吸收N,P元素所能同化碳的能力,是植物養分利用效率的重要指標[54]。本研究中,紅砂葉片C∶N與C∶P呈極顯著正相關關系(表3),說明紅砂葉片具有較高的碳同化能力。 廣泛分布的植物對其環境適應的范圍較廣,可以通過表型可塑性、局域適應等來面對環境變化,其選擇的策略將決定植物應對氣候變化的能力[55],一般來說,葉性狀若與種源地環境因子顯著相關,則可認為發生局域適應[56]。結合RDA分析和逐步線性回歸法分析,白刺葉片N含量與年均降水量有相關性(圖3,表6),這可能是因為本研究4個種源地干旱少雨,除LZ種源外,其余種源年均降水量均不足200 mm,而且水分是植物生長發育過程中所必需的因子,與其他因子相比,植物功能性狀對水分的響應更為顯著[33],因此說明白刺葉片N含量主要受水分的影響。一般來說,海拔對植物功能性狀的影響機理主要是隨著海拔的升高,植物生長發育逐漸受到溫度和水分的限制,因而植物對環境的適應性會發生顯著變化[57]。本研究結果顯示,白刺肉質化程度和含水量與海拔呈極顯著正相關關系,組織密度與海拔呈極顯著負相關關系(圖3,表6),并且海拔對植物比葉面積、組織密度、干物質含量等影響顯著的結論已得到證實[57-58],因此白刺肉質化程度、含水量和組織密度隨海拔的變化受溫度和水分的限制。海拔對紅砂葉片比葉面積和P含量的影響呈顯著負相關關系(圖4,表6),并且隨著海拔的上升,比葉面積和P含量趨向于降低(圖1,圖2),這可能是由于本研究所采樣的4個種源地均屬于高海拔地區,使得植物生長所需要的溫度和水分受到了影響,植物為了適應高海拔的惡劣環境,其物質代謝發生改變,導致植物獲取資源的能力和對環境的適應能力發生了變化[59]。因不同種源地植物葉功能性狀的變異受到多個地理環境因子的共同影響,本文只選取了緯度、經度、海拔、年均降水量、年均溫度等環境因子對葉片功能性狀進行研究,沒有涉及到各種源地物種所在地的土壤養分、坡向、相對濕度、積溫等環境因子。因此,在今后的研究中,還需在這方面作進一步研究。 通過研究同質園中各地理種源白刺和紅砂葉功能性狀的種間差異,發現白刺在適應環境的過程中可能產生了遺傳分化,且受環境影響可能形成了遺傳穩定性,而紅砂在同質園中不受種源環境的影響;兩種植物葉片生長發育主要受氮素限制;受環境的影響,兩種植物葉片內部功能性狀的平衡變化形成了對不同環境的適應策略;分析白刺和紅砂葉功能性狀與種源環境因子的關系,發現海拔是影響兩種植物的主導環境因子,故兩種植物生長受溫度和水分的限制。3.3 白刺和紅砂葉功能性狀間的關系
3.4 種源地環境對白刺和紅砂葉功能性狀的影響
4 結論
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國科技論壇(2017年7期)2017-07-25 08:49:53
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
