感染國槐帶化病植株內源激素及轉錄組分析
2022-03-26 03:44:18劉文汝冀勝鑫甄志先
林業科學研究 2022年1期
關鍵詞:差異
劉文汝,冀勝鑫,甄志先,2*
(1.河北農業大學林學院,河北保定 071001;2.河北省林木種質資源與森林保護重點實驗室,河北保定 071000;
3.河北農業大學園藝學院,河北省蔬菜種質創新與利用重點實驗室)
國槐作為我國的特色樹種,樹冠碩大,姿態優美,具有極高的園林觀賞價值,在城市綠化中發揮著重要作用。國槐帶化病是近年國槐苗木繁育及園林景觀營造中危害比較嚴重的一種侵染性病害,其發病后導致國槐生長減緩,枝條扁莖、頂端小葉叢生,并最終發生枯死,不僅給園林生產帶來較大的經濟損失,而且近年來該病發生日趨嚴重,有些苗圃中國槐受害率已達30%,嚴重威脅園林綠化生態效益與觀賞價值[1-2]。自上世紀90 年代首次證實國槐帶化病為植原體侵染造成以來,國內外對其關注較少且研究并不深入。直到近年隨著分子檢測技術的快速發展,杜銀銀[3]通過采用PCR 快速擴增及生物信息學技術首次明確了國槐帶化病植原體的分類地位,并對國槐帶化病植原體2 個株系(16Sr ID 與16Sr I-B)之間的遺傳相關性和親緣關系進行了鑒定分析。但由于植原體觀察及體外培養難度較大[4],因而很難對植原體病害的致病機理及病菌-寄主間互作機制進行深入探究。轉錄組學研究可以比較精準的反映寄主受到侵染后不同功能基因的響應機制,現已逐漸成為互作解析和差異基因分析的主要技術手段[5]。Fan 和Mardi 等人通過轉錄組分析發現,泡桐和墨西哥檸檬在被植原體侵染后,植原體-病菌互作基因表達上調,并且功能注釋結果表明植株的相關代謝活動和信號通路主要與植物激素有關[6-7];萬倩蕓[8-9]等人在對茅蒼術扁莖機制進行轉錄組分析時也發現扁莖癥狀可能與多種激素代謝通路協同作用相關,并且可能主要與病菌寄生后誘發植物代謝紊亂。而張舒儀等[10]通過對棗瘋病植株進行轉錄組檢測及qPCR 后期驗證后,認為植原體在侵染寄主后可能會迅速激起激素合成及相關保護酶合成基因的表達上升,進而導致激素含量和酶活性代謝紊亂,最終引發叢生、小葉等癥狀。前人結果表明轉錄組學與生理功能驗證相結合可以較為準確、高效地揭示出植原體病害的致病機理,但目前關于國槐帶化病內源激素與轉錄組學研究尚無報道。
本研究通過運用高效液相色譜對病健組織中的生長素、赤霉素、脫落酸、玉米素含量進行比較分析,并對感病和健康國槐枝條進行轉錄組測序和對比,結合GO 和KEGG 功能注釋,篩選出可能導致國槐激素代謝紊亂的關鍵差異基因,旨在通過轉錄組學與生理水平相結合,揭示出國槐感病枝條扁莖機理。為國槐帶化病的防治和抗病品種選育提供理論依據。
1 材料與方法
1.1 材料
試驗樣品在2019 年5、8、9、11 月中每個月份的20 日于保定地區東林水村國槐苗圃園選擇6 年生國槐進行取樣,隨機采取6 組樣品(3 株健康樹、3 株感病樹及3 株感病樹的無癥狀枝條)用消毒高枝剪剪下樹冠頂端2 年生的健康與帶病枝,并將枝條用無菌剪刀剪成1 cm 小段脫去木質部立即放入2 mL 離心管,并用標簽標記后迅速置于液氮冷凍,分成兩組,一組帶回實驗室于零下20℃~80℃超低溫冰箱保存,用于測定激素含量。另一組送轉錄組高通量測序。
1.2 內源激素的高效液相色譜測定
參考杜紹華等[11]的HPLC 法進行提取、純化、分離略有改動。稱取樣品1 g 用液氮研磨,加入8 mL 80%的預冷甲醇,4℃避光浸提12 h;避光超聲1 h,4℃ 12 000 rpm 下離心15 min,將上清液轉移至10 mL 離心管;加入0.2 g PVPP 去除酚雜質,超聲0.5 h,4℃ 12 000 rpm 離心5 min,轉移上清液到新的離心管中;用冰乙酸調節pH 值為2.9,用C18純化移入10 mL 試管,加入1 mL pH=8 的甲醇沖洗即進樣瓶。試驗中以生長素(IAA)、赤霉素(GA)、玉米素(ZT)、脫落酸(ABA)為標樣,并建立標樣的標準曲線。本試驗采用高壓液相色譜儀為日本產HI-TACHI L-7420 型,流動相為100%甲醇:0.7%乙酸體積比為55∶40∶5,紫外檢測儀檢測波長為254 nm,流速0.7 mL·min-1,柱溫30℃,進樣量為10 uL。
1.3 總RNA 的提取、文庫構建和測序
將國槐韌皮部用液氮進行研磨,用天根生化科技有限公司試劑盒進行總RNA 的提取。用瓊脂糖凝膠電泳的方法檢測總RNA 是否存在污染、降解。用Nanodrop 檢測總RNA 的純度(OD260/280比值)、濃度核酸吸收峰是否正常。用Agilent 2100精確檢測RNA 的完整性,檢測指標包括:RIN值、28S/18S、圖譜基線有無上抬、5S 峰。樣品總RNA 檢測合格后寄到北京組學生物科技有限公司,進行cDNA 文庫的構建,文庫構建完成之后使用Agilent 2100 對文庫的insert size 進行檢測,insert size 符合預期后用qPCR 方法對文庫的有效濃度進行準確定量(文庫有效濃度> 2 nM),完成庫檢。庫檢合格后,用Illumina 測序平臺進行測序。
1.4 測序數據處理及分析
對Raw Data 進行數據過濾,去除其中的接頭序列及低質量Reads 獲得高質量的CleanData。然后用Quality(Score 或Q-score)進行堿基識別,使用Phred 的公式(Q-score=-10 log10p)來獲得Phred 堿基質量值,準確計算出堿基質量值與堿基識別出錯的概率的對應關系。堿基質量值越高表明堿基識別越可靠,準確度越高。經過質量控制之后得到的高質量的Clean Data,用一款專門為高通量轉錄組測序設計的組裝軟件Trinity[12]對其進行序列組裝拼接。以Trinity 拼接得到的轉錄本作為參考序列(Ref),將每個樣品的Clean Reads 對Ref做比對分析。并根據基因在不同樣品或不同樣品組中的表達量進行差異表達分析、差異表達基因功能注釋和功能富集等分析。
1.5 內源激素差異基因qRT-PCR 驗證
通過對轉錄組測序所得內源激素相關數據進行篩選差異基因,采用SYBR 法對篩選的差異基因進行實時熒光定量PCR 驗證。并以Actin 作為內參基因。Actin-F(5′-GAGGGCCCAGGTTCTGA TTT-3′)和 Actin-R(5′-GCACCCACCTGCATT TTACC-3′)。通過NCBI 設計引物并進行熒光定量分析。實時熒光定量PCR 按照以下步驟進行擴增:95℃預變性3 min,95℃變性5 s,72℃退火30 s,40 個循環。按照2-ΔΔCT[13]計算基因相對表達量。用 SPSS20.0 對數據進行單因素方差分析(ANOVA),采用LSD和 Duncan’s 方法進行兩兩比較、差異顯著性分析(P< 0.05)。
2 結果與分析
2.1 病、健國槐內源激素含量變化
通過對處于生長期的感病國槐(BZ)、健康國槐(JK)和感病國槐樹上的表象健康枝(BJ)樹皮的內源激素含量的測定,研究該病害發生期對國槐內源激素的影響。由圖1 可以看出,BZ、BJ和JK 間的Zeatin、GA、IAA 及ABA 含量變化趨勢基本一致。Zeatin 含量在8、9 月份達到高峰,之后呈下降趨勢,其中8 月份BZ 中的Zeatin 含量顯著高于BJ 和JK,BJ、JK 無差異。其它月份中三者間無顯著差異;GA 含量與Zeatin 含量年變化較為相似,在8 月份達到最高,之后處于下降趨勢。但其中8 月份BZ 的GA 含量顯著低于JK 和BJ,BJ 其次,BZ 中的GA 含量最低,其它月份三者間并無差異;而IAA 和ABA 的含量變化呈相反趨勢,IAA 除5 月份最高外,之后一直呈下降趨勢,并在11 月份達到最低,其中JK 含量最高,BJ 其次,BZ 中的IAA 含量最低。ABA 則是從5 月份開始一直呈上升趨勢,在11 月份達到最高,BZ 中的含量顯著高于JK 和BJ,BJ 與JK 的ABA 含量在5、8、9 月份中均無差異。
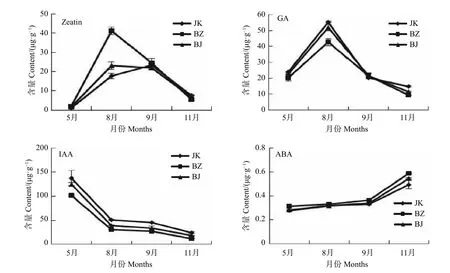
圖1 病健國槐內源激素含量Fig.1 Content of endogenous hormones in the diseased and heathy
2.2 轉錄組測序與組裝分析
通過對國槐感病樣本(BZ1、BZ2、BZ3)和健康樣本(JK1、JK2、JK3)進行高通量轉錄組測序,分別獲得26 499 450、22 444 628、33 026 072、37 221 613、28 464 069 和27 856 864 條Raw Reads,在去除含有接頭的Reads 以及低質量的Reads(包括去除N 的比例大于10%的Reads)后,最終得到20 427 416、17 402 101、25 556 038、28 443 464、21 878 012 和21 192 357 條Clean reads。其 中Q20堿基百分比分別是98.20%、98.16%、98.05%、97.95%、97.93%和97.99%,GC 堿基含量則均約達到44%(表1)。經過過濾后,最終得到可靠的轉錄本數據,并進行下一步生物學分析。
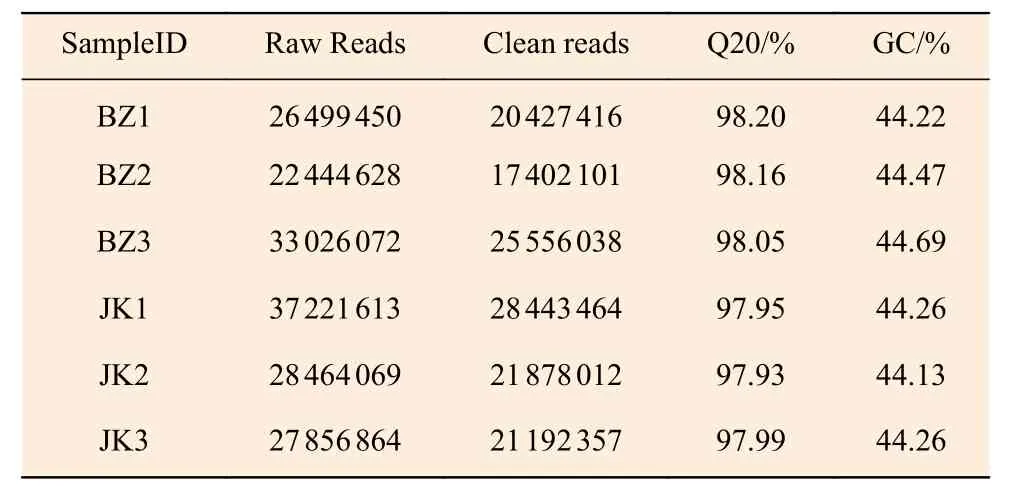
表1 每個樣品有效測序數據統計Table 1 Statistics of valid sequencing data of each sample
2.3 Unigne 功能基因注釋
采用BLAST[14]軟件對Unigene 序列與NR、Swiss-Prot[15]、GO[16]、COG/KOG[17]、KEGG[18]數據庫進行比對,共獲得Unigne 注釋功能信息有72 637條,占全部Unigene 的60%,其中NR 數據庫中注釋的信息最多,而COG 數據庫中被注釋信息最少。通過使用KOBAS2.0[15]獲得Unigene 在KEGG 中的KEGG Orthology 結果,在預測Unigene 的氨基酸序列之后使用HMMER[17]軟件與Pfam[19]數據庫比對,獲得Unigene 的注釋信息(表2)。
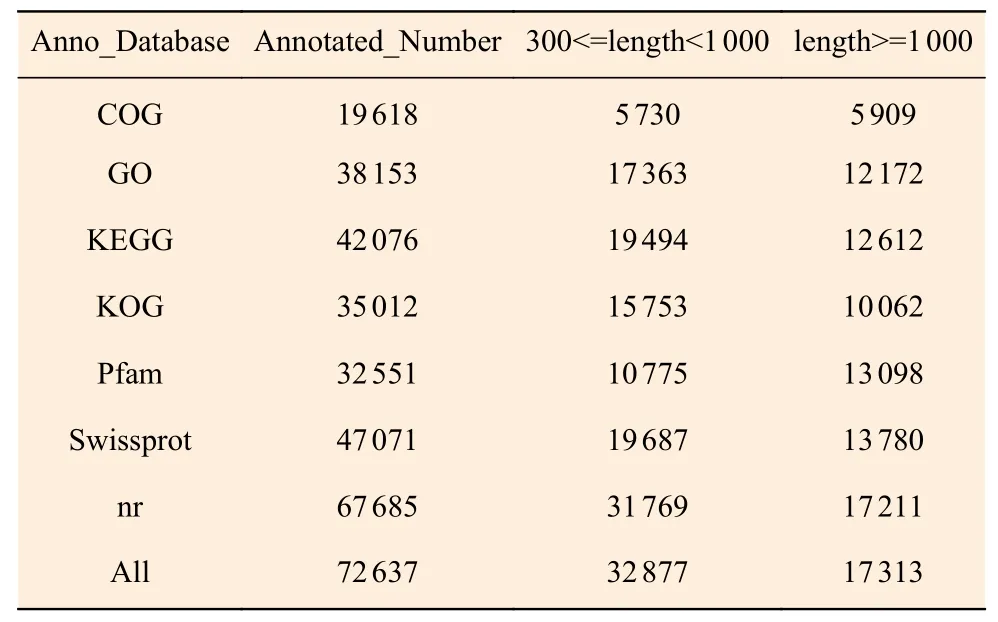
表2 Unigene 功能注釋統計Table 2 Unigene function annotation statistics
2.4 感病國槐與正常國槐基因表達差異分析
通過采用EBSeq[20]對所得數據進行差異分析,并采取Fold Change≥2 且FDR<0.01 作為篩選標準。如圖2 所示,綠點代表下調差異表達基因,紅點代表上調差異表達基因,藍色的點代表非差異表達基因。其中共得到差異表達基因數目4 714條,國槐帶化病樣品相對于健康國槐樣品上調基因有2 283 條,下調基因有2 431 條。這些結果表明,國槐在受到植原體侵染后其植物體內多數的基因表達受到抑制。
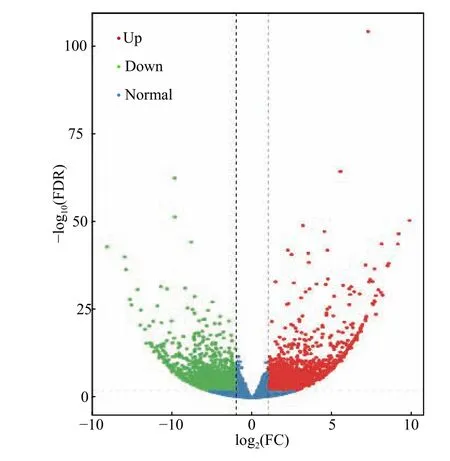
圖2 國槐帶化病株和健康株之間差異表達基因的火山圖Fig.2 Volcanic diagram of differentally expressed genes between locust striated and healthy plants
2.5 差異表達基因功能注釋和富集分析
2.5.1 差異表達基因GO 功能分析 通過對感病與健康國槐轉錄組差異表達基因進行GO 功能注釋富集分析,結果可分為3 大類:細胞組成(cellular component)、分子功能(molecular function)、生物學過程(biological process)。其中注釋到細胞組成有7 個,分子功能10 個,生物學過程3 個(圖3)。細胞組成中以膜的組成成分(783 個)和質膜(141)中差異表達基因最多,另外膜的組成成分、運動蛋白復合物等離子組分以及細胞壁等差異表達基因富集表達均達到顯著水平;分子功能中以蛋白絲氨酸蘇氨酸激酶(136 個)和DNA 結合轉錄(84 個)差異表達基因最多,而在木糖葡聚糖、木糖葡聚糖轉化和DNA 結合轉錄等富集基因中達到顯著水平;生物學過程中蛋白磷酸化中的差異表達基因居多并達到顯著水平。結果表明細胞組成和分子功能的基因富集更為顯著,這說明在國槐受到植原體侵染以后,細胞和分子功能方面均發生明顯的差異變化,并可能受到上述富集差異基因的調控。
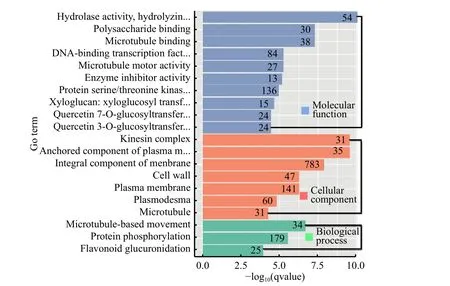
圖3 差異表達基因GO 富集分析Fig.3 GO enrichment analysis of differentially expressed genes
2.5.2 差異表達基因KEGG 注釋 在生物體內,不同的基因產物相互協調來行使生物學功能,對差異表達基因的通路(Pathway)注釋分析有助于進一步解讀基因的功能。通過對差異基因(2 539個)進行KEGG 功能注釋分析,發現差異基因被富集到48 條代謝通路,本研究挑選了富集最顯著的20 條pathway 條目在圖中進行展示(圖4)。其中代謝通路從低到高依次為:戊糖和葡萄糖醛酸互相轉化、卟啉和葉綠素代謝、AMPK 信號通路、癌變中的微型核糖核酸、多巴胺能受體、半光氨酸和蛋氨酸代謝、泛素介導的蛋白水解、MAPK 信號通路、溶酶體、卵母細胞減數分裂、RNA 降解、抗原加工和傳遞、苯丙氨酸合成、Ras 信號通路、內噬作用、植物激素信號轉導、植物-病原互作、淀粉和蔗糖代謝、Toll 受體信號轉導途徑和NF-KB信號傳導途徑。其中,匯集差異基因最多的Toll受體信號轉導途徑和NF-KB 信號傳導途徑均表現為顯著下調,另外,控制細胞內噬作用、減數分裂以及抗原加工與傳遞同樣表現為顯著下調。
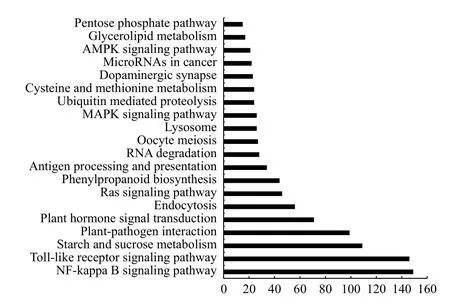
圖4 差異表達基因KEGG 富集Fig.4 Distribution of differentially expressed gene KEGG
2.5.3 國槐激素差異表達基因代謝通路 感病國槐和正常國槐激素代謝通路的差異基因被注釋到25 個DEGs(圖5)。由圖可知,其中有7 個下調基因(綠色標注),8 個上調基因(紅色標注)。其中,在色氨酸代謝通路上(生長素),AUX1 節點上有3 個差異表達基因下調,而在GH3 節點有3 個差異表達基因上調。而在玉米素信號傳導代謝通路上(細胞分裂素),CRE1 基因節點上涉及到7 個差異基因下調;AHP 節點上有3 個差異基因下調;ARR-A 節點處有3 個差異基因下調。二萜生物合成(赤霉素)代謝通路上關鍵調控基因TF 涉及13 個差異基因上調。生長素、玉米素合成關鍵基因的下調以及赤霉素合成基因的上調,可能是導致國槐枝條頂端生長停滯、發生橫向卷曲生長的主要原因。類胡蘿卜素生物合成(脫落酸)代謝途徑中,ABF 調控關鍵基因涉及10 個差異基因上調,產生這種原因可能與植原體的致病因子的基因干擾或植物體受到脅迫產生的自激反應有關。
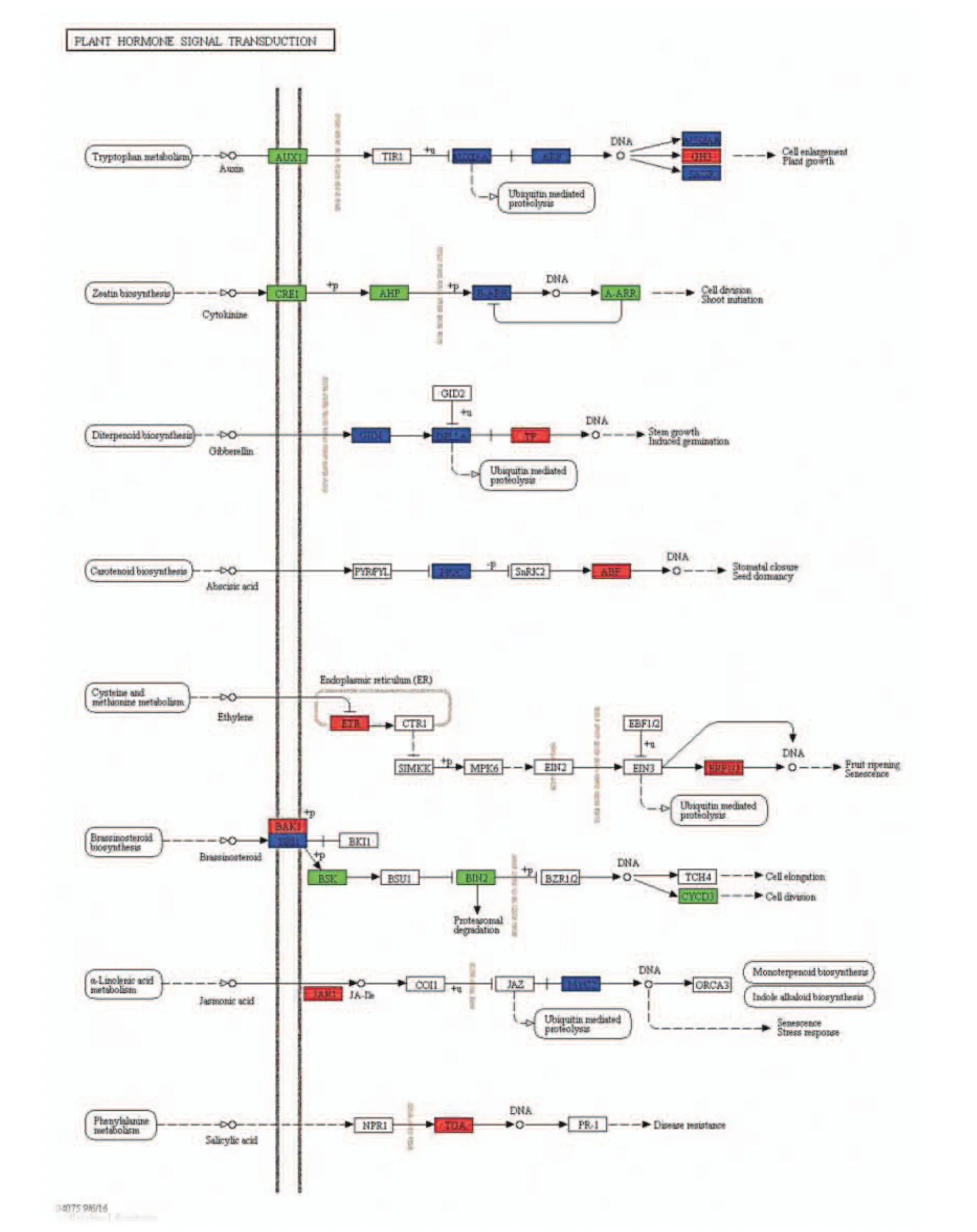
圖5 植物激素傳導(Ko04075)路徑Fig.5 Pathway map of phytohormone transmission (ko04075)
2.5.4 國槐激素差異表達基因qRT-PCR qRT-PCR驗證本研究通過對轉錄組差異基因數據進行篩選獲得與生長素、細胞分裂素、赤霉素和脫落酸代謝相關基因共6 個(表3),其中,與生長素代謝相關基因AUX 1 個(GHLAX5),與細胞分裂素信號傳導受體CRE1 相關基因3 個(GHAHK4、GHAHP4和GHARR8),調控赤霉素信號傳導的相關基因TF 1 個(GHTF),脫落酸信號轉導調控因子相關基因ABF 1 個(GHABF2)。通過對上述6 個激素代謝關鍵基因進行熒光定量表達驗證,如圖6 所示,雖然定量表達與轉錄組結果具有一定差異,但表達趨勢較為一致,生長素、細胞分裂素代謝關鍵基因表現為顯著下調,赤霉素和脫落酸代謝關鍵基因表現為顯著上調。
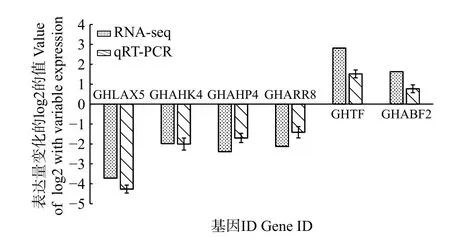
圖6 差異表達基因的qRT-PCR 驗證Fig.6 Validation of differentially expressed genes using qRT-PCR
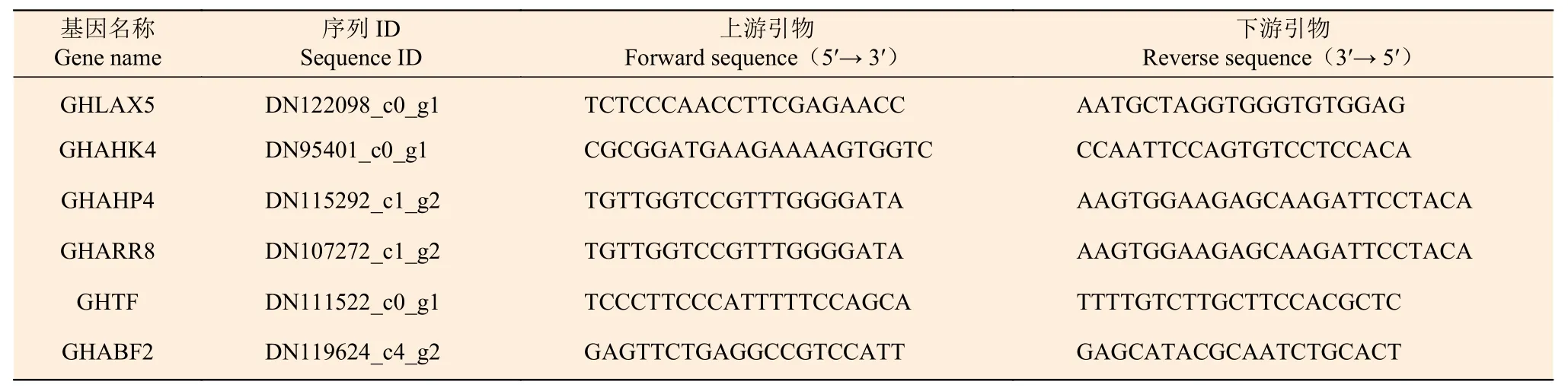
表3 差異表達基因qRT-PCR 引物Table 3 Primers of differentially expressed genes QRT-PCR
3 討論
多數研究認為,植原體在侵入植物體后,產生的某種產物會導致植物體內激素失衡,進而導致植物體表型生長發生改變,出現小葉、叢生等癥狀[21-22]。因而通過研究病株機體內源激素含量進而解釋植原體病害的致病機理逐漸成為植原體病害研究的主要手段。趙錦等[23]研究發現棗瘋病株在1 年中的發育旺盛期內玉米素含量表現為異常顯著升高,而且在生長后期內玉米素含量也顯著高于健株。Kesumawati[24]的研究結果也表明在八仙花受到植原體感染的組織中含有較高含量的玉米素。另外,有研究認為玉米素和生長素的比例變化失調可能是導致植物體發生小葉、叢枝等病狀的主要因素[23,25-26]。本研究發現國槐在受到植原體侵染以后,其生長旺盛的8 月份期間,病枝內玉米素顯著高于健樹和病樹上的無癥狀枝,而在其他月份并未產生較大差異;但在旺盛生長期內病枝中的赤霉素、生長素含量則均顯著低于健樹和病樹上的無癥狀枝,在旺盛生長期內的玉米素含量與生長素比例的嚴重失衡可能是導致國槐感病枝條發生小葉和叢枝的主要原因,與前人得出的玉米素含量的顯著增加是導致器官分化方向改變,進而引起叢枝的結果較為一致[23]。而且生長素、赤霉素的顯著降低也可能是導致國槐病枝向內卷曲和停止伸長生長的主要原因[27]。
植原體的功能基因組較小,在侵染寄主后,其基因組片段會整合到植物的細胞基因組中,研究發現植原體基因組缺乏脂肪酸和氨基酸合成等基本的生化途徑[23],因而可能主要通過基因表達調控幫助其完成寄生,并會對寄主植物產生影響。有研究發現,植原體的簡單基因組中存在編碼致病效應因子蛋白的基因,部分致病效應因子通過與標靶蛋白相結合干擾寄主植物的正常代謝以達到干擾寄主植物防御功能的目的[4],翠菊黃化植原體叢枝株系所產生的致病效應蛋白SAP11AYWB 能夠通過靶標作用寄主的TCP 轉錄因子,進而影響寄主植物體內的抗體活性甚至使得該類轉錄因子失活,導致叢枝癥狀的產生[28]。本研究通過轉錄組測序發現國槐帶化病枝條內的多數基因表達受到抑制,細胞組分與分子功能富集差異基因最多,并且差異基因多數注釋到植物的免疫應答機制、植物-病原互作和激素信號轉導等方面,其中Toll 受體信號轉導途徑和NF-KB 信號傳導途徑匯集差異基因最多,且均表現為顯著下調,有研究指出Toll 受體在動物體內主要通過識別來自細菌和病毒的侵入信號及細胞損傷狀況,進而對免疫應答起到關鍵調控作用[29],而NF-KB 信號通路作為Toll 受體信號傳導激活的下游通路,常與Toll 受體在免疫機制中形成聯控作用[30],這說明國槐枝條在受到植原體侵染以后,機體調控免疫應激的能力顯著降低,細胞衰亡程度加快。另外,轉錄組結果表明在激素代謝通路中調控生長素代謝的響應因子AUX1 的表達顯著下調,這與前人得到的在表達植原體致病蛋白“天狗蛋白”的植物中生長素響應因子顯著下調,且生長素含量顯著降低的結果較為一致[31]。同樣,Jennifer研究表明擬南芥組氨酸激酶(CRE)作為一種玉米素受體,可能在玉米素功能信號傳導中發揮著關鍵作用[32],而Riefler 等人研究結果也指出植物體內的玉米素含量可能受到CRE 受體的反饋抑制,在CRE 受體突變的植株中積累了更高含量的細胞分裂素[33],本研究結果表明在玉米素代謝通路中,CRE1 受體基因表達顯著下調,另外負調控細胞分裂發育的A 型ARRs 基因[32]表達也顯著下調,這與前人報道稱泡桐叢枝病枝發病后,病枝內玉米素代謝通路基因表達顯著上調[34]的研究結果相反,但這些結果說明國槐帶化病枝內積累的較高含量細胞分裂素以及細胞增生進而發生橫向生長可能與CRE1 和A 型ARRs 等基因的顯著下調表達有關,具體原因則需要對這兩個基因進行進一步功能驗證。另外本研究通過對轉錄組篩選得到的生長素、玉米素、赤霉素和脫落酸代謝通路中的關鍵基因進行熒光定量表達驗證,結果表明表達趨勢與轉錄組數據較為一致,這說明本研究所進行的轉錄組驗證準確性較高。綜上,可能正是由于國槐病株內源激素代謝關鍵基因表達顯著上調或下調,進而導致生長素、玉米素、赤霉素和脫落酸的含量積累發生變化,這與前人在對篩選得到的關鍵基因進行定量表達驗證后[10,34]所得到的結果一致。
4 結論
本研究通過探究國槐帶化病株內激素含量與基因表達之間的變化聯系,認為國槐植株在受到病原侵染以后,其病枝內玉米素、生長素和赤霉素的代謝關鍵調控基因表達顯著上調或下調,進而導致病枝內的玉米素和生長素比例失衡,以及赤霉素顯著降低,這可能是導致國槐病枝表現叢生、小葉及枝條帶化生長的主要原因。本工作為進一步深入研究國槐帶化病及其防治工作提供了理論基礎。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
