煙草白粉病抗性基因型的多重PCR 檢測體系及其在回交育種中的應用
2022-03-25 13:26:36林世鋒王仁剛王自力任學良龍明錦史躍偉
煙草科技 2022年3期
林世鋒,王仁剛*,潘 飛,王自力,任學良,龍明錦,史躍偉
1. 貴州省煙草科學研究院 煙草行業(yè)煙草分子遺傳重點實驗室,貴陽市觀山湖區(qū)龍灘壩路29 號 550081
2. 上海農科種子種苗有限公司,上海市閔行區(qū)北翟路2901 號 201106
煙草白粉病是煙草生產中的主要病害之一,該病害發(fā)生普遍,給烤煙生產造成了嚴重的經濟損失[1]。目前,生產上對煙草白粉病的防治以化學防治為主,不僅需要相應的人力、物力投入,還會帶來煙葉農殘超標、環(huán)境污染等問題[2]。闡明煙草白粉病抗性機制,開展抗病育種,從遺傳本質上提高煙草對白粉病的抗性是防治煙草白粉病最經濟、最有效且對人畜安全的方法。
Mildew resistance locus O(MLO)蛋白是植物特有的一類蛋白家族,其中的部分成員是白粉病菌成功侵染寄主植物的必要因子[3]。一個或多個MLO基因功能的缺失突變可導致植物對白粉病菌廣譜且持久的抗性[4-7]。已知抗白粉病煙草品種Kutsaga E1和Tsukuba 1 的白粉病抗性來源于煙草品種Kokubu、Fujimura 等[8],通過對煙草品種 Kokubu 的白粉病抗性分子機制研究發(fā)現,NtMLO1 和NtMLO2 基因均為煙草白粉病隱性抗病基因(感白粉病基因),來源于Kokubu 的白粉病抗性是由NtMLO1 和NtMLO2 基因的隱性突變所致。
為開發(fā)相應的分子標記以輔助煙草白粉病抗病育種,張孝廉等[9-10]根據煙草白粉病抗感品種間NtMLO1 和NtMLO2 基因序列差異設計引物,構建了4 個單基因PCR 體系分別用于擴增野生型和突變型NtMLO1 和NtMLO2 基因。基于該研究,設計和組合引物并優(yōu)化PCR 反應體系和反應條件,旨在建立一種用于檢測NtMLO1 和NtMLO2 基因型的多重PCR方法,為煙草白粉病抗病育種回交后代基因型篩選提供可靠的技術手段。
1 材料與方法
1.1 試驗材料
抗白粉病烤煙品種Kutsaga E1和感白粉病烤煙品種K326、紅花大金元和云煙87均由貴州省煙草科學研究院新品種選育三組鑒定和提供。分別以K326、紅花大金元和云煙87 為母本,以Kutsaga E1作為父本進行雜交獲得各自的F1代,其中K326 和Kutsaga E1 的F1代再通過自交或回交(以K326 為父本)產生F2代及回交世代,用于抗譜鑒定和標記分析。
1.2 方法
1.2.1 白粉病菌人工接種
接種鑒定病原菌為煙草白粉病致病菌二孢白粉菌(Erysiphe cichoracearum DC),由貴州省煙草科學研究院新品種選育三組分離和保存。苗期采用抖落法[11-12]接種白粉病菌。
1.2.2 煙草基因組DNA的提取
剪取約100 mg 煙葉置于2 mL 離心管中,于55 ℃干燥箱中干燥24 h。使用SPEX Geno/Grinder 2010 高通量動植物組織研磨機(美國SPEX SamplePrep 公司)將干燥后煙葉振蕩研磨成細粉狀后,一次性加入520.9 μL 新鮮配制的裂解緩沖液(350 μL PBS、0.9 μL RNase A、150 μL Buffer C-L、20 μL Proteinase K)裂解細胞。使用AxyPrep 基因組DNA小量制備試劑盒(美國Axygen公司)提取煙草品種基因組DNA。使用NanoDrop 1000超微量分光光度計(美國Thermo 公司)測定樣品DNA 濃度,將樣品DNA的濃度稀釋至80 ng/μL備用。
1.2.3 引物設計
分析煙草白粉病隱性抗病基因NtMLO1(M1)和NtMLO2(M2)的野生型和突變型基因序列(圖1)[8],相比感病品種,抗病品種NtMLO1 基因內含子7 的3′端2個堿基AG突變?yōu)門A,且抗病品種NtMLO2基因內含子6 的5′端2 個堿基AG 發(fā)生缺失。通過中國煙草基因組數據庫BLAST 工具進行相似性搜索與序列比對(Blastn),綜合考慮M1和M2基因突變位點及其上下游序列特征,設計多對備選引物。經實驗比對,篩選4 對特異性引物,組成2 個多重PCR 反應體系,分別用于擴增M1 和M2 野生型和突變型等位基因(表1)。
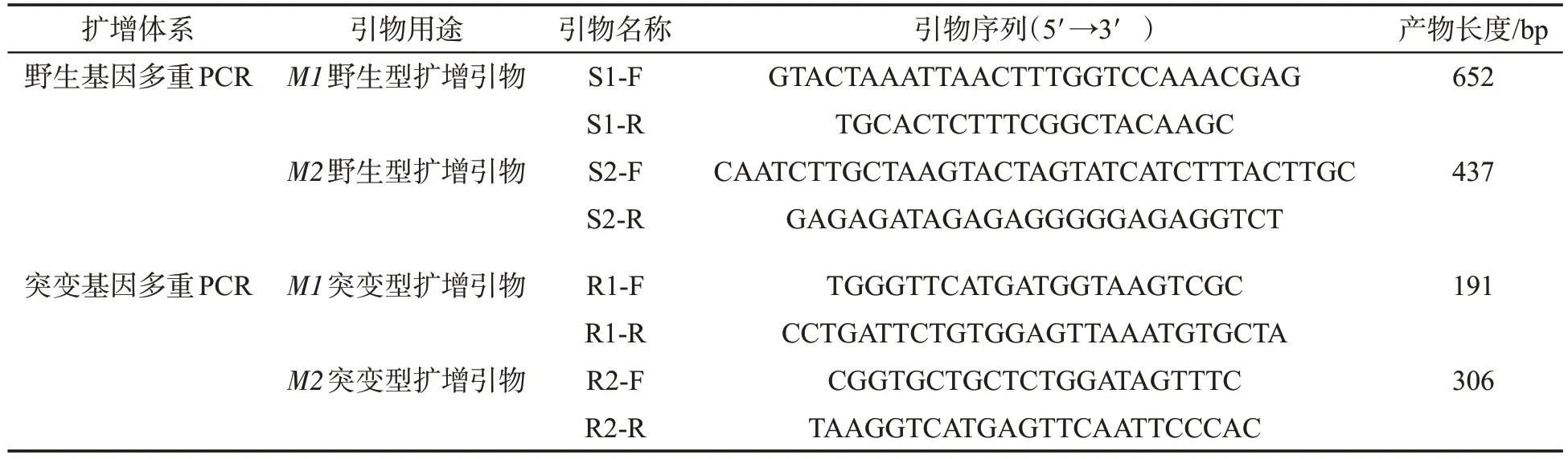
表1 多重PCR引物序列及擴增片段長度Tab.1 Primer sequences and lengths of amplified fragments for multiplex PCR
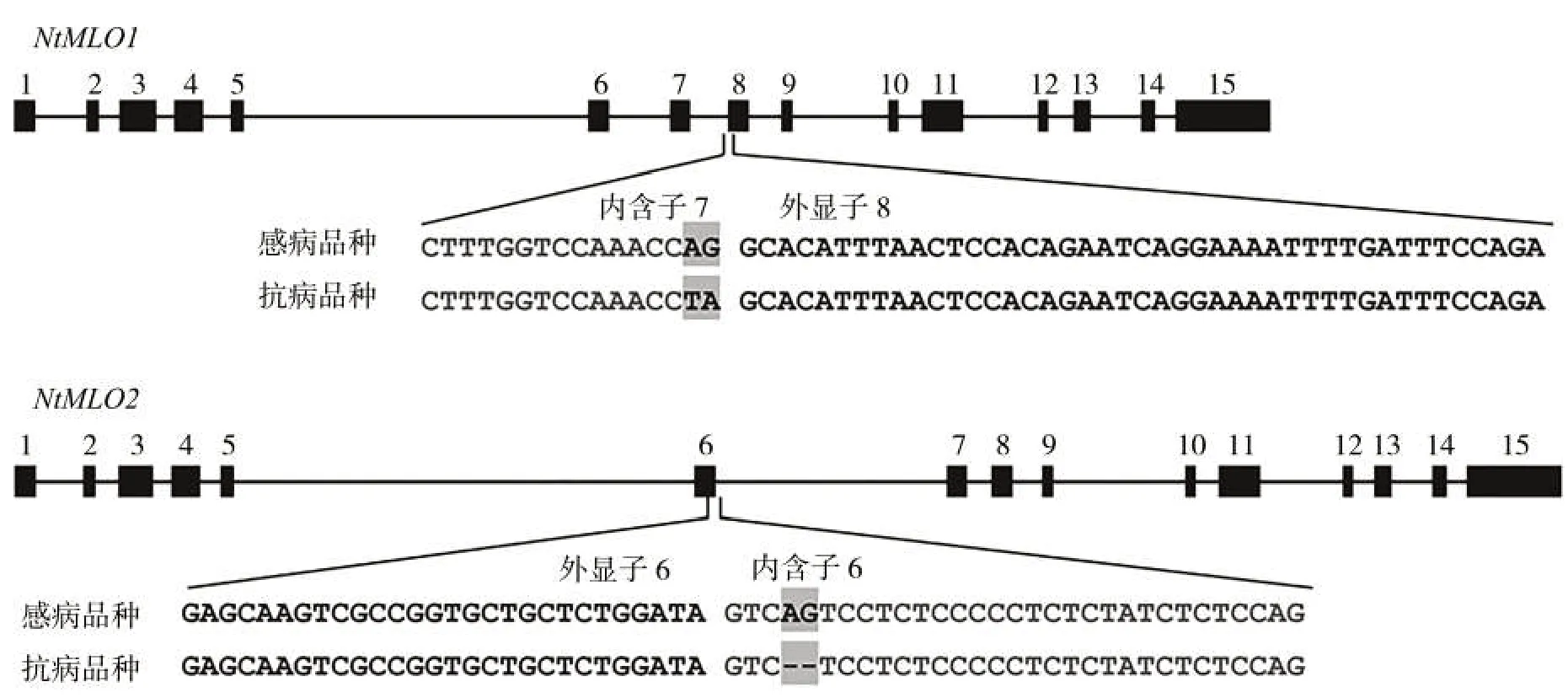
圖1 煙草白粉病隱性抗病基因NtMLO1和NtMLO2的野生型和突變型基因序列比對Fig.1 Alignment of wild-type and mutant sequences of recessive resistance genes NtMLO1 and NtMLO2
1.2.4 PCR擴增及產物檢測
單組引物 PCR 反應體系 20.0 μL:Premix TaqTM10.0 μL,正反向引物(10 μmol·L-1)各0.4 μL,gDNA模板(80 ng·μL-1)1.0 μL,ddH2O補足至20.0 μL。
雙重引物 PCR 反應體系 20.0 μL:Premix TaqTM10.0 μL;M1正反向引物(10 μmol·L-1)各0.4 μL,M2正反向引物(10 μmol·L-1)各0.4 μL;gDNA 模板(80 ng·μL-1)1.0 μL;ddH2O補足至20.0 μL。
Premix TaqTM(TaKaRa TaqTMVersion 2.0 plus dye)購自寶生物工程(大連)有限公司。
PCR 反應程序:94 ℃預變性 3 min;94 ℃變性30 s,60 ℃退火30 s,72 ℃延伸30 s(M1野生型擴增引物對1-M1-F/1-M1-R的延伸時間除外,為40 s),30個循環(huán);72 ℃終延伸10 min。
凝膠電泳檢測:PCR 產物用加有GelRed 核酸染料的2%(質量分數)瓊脂糖在100 V電壓條件下電泳50 min,檢測結果在凝膠成像系統(tǒng)上顯示。
2 結果與分析
2.1 PCR分子標記體系特異性和穩(wěn)定性檢驗
如圖2所示,使用S1或S2引物對進行單重PCR擴增時,在含有M1 和M2 野生型等位基因的感病材料和抗感雜交組合(F1)材料中分別擴增出652 或437 bp的單一特異性條帶;進行雙重PCR擴增時,可以在上述材料中同時擴增出652 和437 bp的特異性條帶。使用R1 或R2 引物對進行單重PCR 擴增時,在含有M1和M2突變型等位基因的抗病材料和抗感雜交組合(F1)材料中分別擴增出191 或306 bp的單一特異性條帶;進行雙重PCR擴增時,可以在上述材料中同時擴增出191和306 bp的特異性條帶。結果表明,特異引物對S1、S2、R1 和R2 具有很好的特異性和穩(wěn)定性,S1和S2的雙重PCR擴增結果是各自單重 PCR 擴增結果的疊加,R1 和 R2 的雙重 PCR 擴增結果是各自單重PCR 擴增結果的疊加,且能根據2個雙重PCR 擴增結果區(qū)分供試材料的3 種基因型,達到預期的基因分型效果。
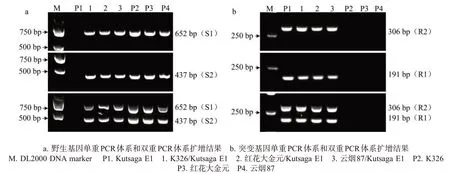
圖2 單重PCR體系和雙重PCR體系的擴增結果Fig.2 Amplification results of monoplex or duplex PCR system
2.2 F2代群體白粉病抗性鑒定
接種10 d 后調查Kutsaga E1 和K326 構建的F2群體(共500株)白粉病抗性,抗病單株為28株,感病單株為 472 株,符合 1 ∶15 的分離比率,證明 Kutsaga E1 的白粉病抗性受2 對獨立遺傳的等位基因(A、a和B、b)控制,抗病基因為隱性抗病基因。
2.3 利用分子標記鑒定F2代群體單株基因型
使用2個雙重PCR擴增體系對F2代群體內不同單株的DNA 進行PCR 擴增,擴增結果見圖3。F2群體中共有9 種基因型,分別為AABB、AABb、AAbb、AaBB、AaBb、Aabb、aaBB、aaBb、aabb,其中僅 aabb基因型(與親本1 一致)抗病,在突變基因雙重PCR體系中擴增出2條特異性條帶,在野生基因雙重PCR體系中無條帶,2個白粉病隱性抗病基因同時發(fā)生純合突變,該后代植株具有白粉病抗性。其他基因型均感病,在野生基因多重PCR 體系中可擴增出1 條或2條特異性條帶。
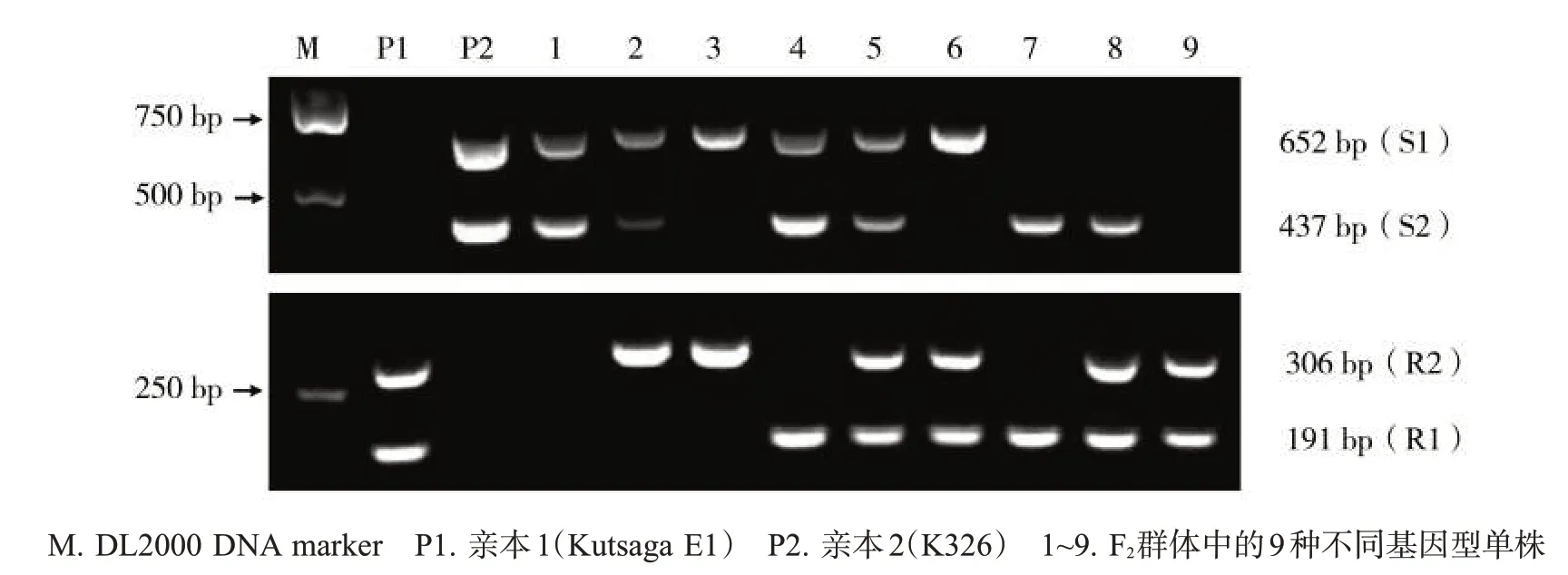
圖3 基于野生基因雙重PCR體系和突變基因雙重PCR體系的F2群體及其親本的檢測結果Fig.3 Detection results of parents and their F2 population via wild-type and mutant allele-specific multiplex PCR systems
2.4 分子標記在煙草白粉病抗性回交改良中的應用
使用野生基因雙重PCR 體系和突變基因雙重PCR 體系對K326 和Kutsaga E1 的回交后代分離群體進行PCR 擴增,結果見圖4。不同于F2代分離群體,該群體有4 種基因型:AABB 基因型在突變基因多重PCR體系中擴增時無條帶;AaBB基因型在突變基因多重PCR體系中只擴增出一條191 bp的特異性條帶;AABb 基因型在突變基因多重PCR 體系中擴增出一條306 bp的特異性條帶;AaBb基因型可在突變基因多重PCR體系中同時擴增出191 bp和306 bp的 2 條特異性條帶,分別見圖 4 中單株 4、6、9、17 和21。篩選出攜帶2個隱性抗白粉病基因(AaBb)的單株,選擇其中表型盡可能接近輪回親本的單株繼續(xù)與輪回親本雜交或進行自交獲得隱性純合子(aabb)。自交子代純合子篩選的方法同2.3節(jié),使用2 個多重PCR 體系對BCnF2代群體內不同單株的DNA進行PCR擴增,選擇基因型為aabb的子代單株。

圖4 2個多重PCR體系在親本及回交后代部分單株中的檢測結果Fig.4 Detection results of parents and some of their backcross progenies via two duplex PCR systems
在常規(guī)回交育種過程中(圖5a,以Kutsaga E1和K326為例),當目標性狀受隱性基因控制時,不能依據表型在回交后代(BCnF1)中選擇出具有目標性狀的植株,每次回交之前均需自交一次,然后從分離群體(BCnF2)中選擇具有隱性目標性狀的植株進行下一輪的回交。這樣一代回交,一代自交,交替進行,防止目標性狀丟失。不同于常規(guī)回交育種,分子標記輔助選擇育種(圖5b,以Kutsaga E1和K326為例)可利用分子標記直接對BCnF1單株進行基因型鑒定,選擇攜帶2個隱性抗白粉病基因的單株,不需自交一代,且無需進行自交后代單株白粉病菌的接種鑒定工作。除可顯著縮短育種時間外,利用分子標記鑒定抗性還具有以下優(yōu)點:①無需根據表型進行人工判斷,檢測準確率達100%;②減少人為接種病害對其他實驗材料的污染;③不需考慮人工接種病害時對溫度條件的需求;④提取的后代單株的DNA還可用于其他性狀的分子標記輔助選擇。

圖5 常規(guī)回交育種(a)與分子標記輔助選擇育種(b)定向改良煙草白粉病抗性過程比較(以Kutsaga E1和K326為例)Fig.5 Processes of conventional backcross breeding (a) and molecular marker-assisted selection breeding (b) for resistance to powdery mildew in tobacco ( an example with Kutsaga E1 as the donor and K326 as the acceptor)
3 討論
通過比較M1和M2的基因組序列,發(fā)現M1基因在突變位點處的堿基序列與M2 基因相應位置的堿基序列一致性較高。其中,M1 野生型基因與M2 野生型或突變型基因的一致性更高(圖6a)。因此,在M1 基因突變位點設計的野生型基因特異性引物可能與M2 相應位置的堿基序列發(fā)生結合,反之亦然。如不充分考慮該情況,設計的引物可能會在同時擴增M1和M2野生型等位基因的多重PCR體系中發(fā)生交錯組合(圖6b、圖6c、圖6d),從而影響擴增效果,這一推論也在之前的預實驗中得到證實。根據上述序列比較結果,在M1突變位點處設計野生型基因的正向特異引物(圖6e),在M2突變位點處設計野生型基因的反向特異引物(圖6e),并在上述引物3′端倒數第3位引入1個錯配堿基,以提高PCR延伸反應的特異性。

圖6 NtMLO1和NtMLO2基因突變位點區(qū)域的DNA序列比較及引物設計策略分析Fig.6 Multiple alignments and primer design strategies of DNA sequences of mutation site regions in NtMLO1 and NtMLO2 genes
本研究中最初試圖通過優(yōu)化反應條件,獲得一個同時檢測M1和M2野生型和突變型的四重PCR體系,擴增結果如圖7所示。由于引物間的干擾,造成四重PCR 體系擴增效率較低,需使用價格較高的熱啟動Taq 酶(Premix TaqTMHot Start Version)進行40個PCR循環(huán),且該體系重復性較差,故無法應用于分子標記輔助育種。因此,分開構建2 個雙重PCR 體系,包括擴增M1 和M2 野生型等位基因的雙重PCR體系和擴增M1和M2突變型等位基因的雙重PCR體系,同時通過對PCR反應體系和反應條件的優(yōu)化,確保了擴增結果的重復性和可靠性(圖2、圖3、圖4)。
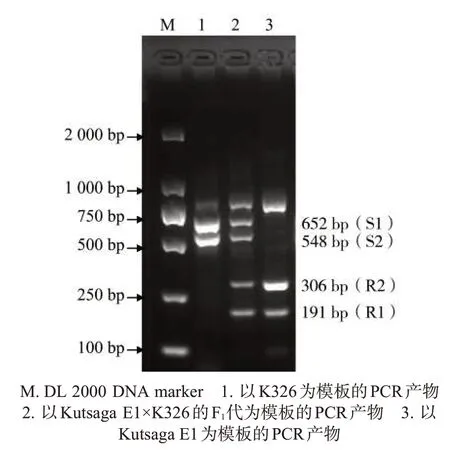
圖7 同時檢測NtMLO1和NtMLO2野生型和突變型等位基因的四重PCR擴增結果Fig.7 Amplification results of wild-type and mutant alleles of NtMLO1 and NtMLO2 genes via four pairs of primers in one PCR system
4 結論
根據煙草白粉病感病基因M1和M2野生型和突變型等位基因序列差異設計引物,建立了2 個雙重PCR 體系,包括擴增M1 和M2 野生型等位基因的雙重PCR體系和擴增M1和M2突變型等位基因的雙重PCR 體系,與單重PCR 體系相比,縮短了鑒定時間、提高了檢出效率,為煙草白粉病抗病育種回交后代基因型篩選提供了一種客觀且可靠的技術手段。
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
新世紀智能(數學備考)(2020年11期)2021-01-04 00:38:16
中國外匯(2019年17期)2019-11-16 09:31:14
浙江中西醫(yī)結合雜志(2017年2期)2017-01-12 18:23:59
當代化工研究(2016年9期)2016-03-20 16:22:08
現代企業(yè)(2015年1期)2015-02-28 18:43:18
新高考·高一物理(2014年1期)2014-09-18 01:26:07
聲屏世界(2014年6期)2014-02-28 15:18:09
中國煙草學報(2012年5期)2012-04-12 06:21:18
中國煙草學報(2012年2期)2012-04-09 06:44:56
