小球藻(Chlorella vulgaris)耐受短期模擬酸雨及紫外輻射的光合生理特性研究
2022-03-11 02:32:40楊雨玲章鵬李亞鶴蔡飛畢淑峰李偉
生態(tài)毒理學(xué)報(bào) 2022年6期
關(guān)鍵詞:影響
楊雨玲,章鵬,李亞鶴,蔡飛,畢淑峰,李偉,*
1. 寧波大學(xué)海洋學(xué)院,寧波 315211 2. 黃山學(xué)院生命與環(huán)境科學(xué)學(xué)院,黃山 245041 3. 黃山學(xué)院新安江流域生態(tài)環(huán)境保護(hù)研究中心,黃山 245041 4. 黃山市屯溪區(qū)林業(yè)局,黃山 245041
浮游植物作為內(nèi)陸水域生態(tài)系統(tǒng)中初級(jí)生產(chǎn)力的重要組成,是構(gòu)成食物網(wǎng)的基礎(chǔ),在生物地球化學(xué)循環(huán)中扮演重要角色。環(huán)境因子如何影響、調(diào)節(jié)浮游植物的光合生理、初級(jí)生產(chǎn)、種群結(jié)構(gòu)及豐度是生態(tài)學(xué)研究的重要內(nèi)容。因空間位置(經(jīng)、緯度)、水體類型(湖泊、河流)、局域環(huán)境條件的不同,影響浮游植物光合生理、群落結(jié)構(gòu)的主要環(huán)境因子在不同流域存在一定差異。
黃山市是世界文明的生態(tài)旅游城市,同時(shí)也是我國(guó)受酸雨影響較大的城市之一(可參見(jiàn)歷年的《中國(guó)生態(tài)環(huán)境狀況公報(bào)》)。酸雨引起的水體酸化是影響新安江流域浮游植物的重要脅迫因子。水體酸化可影響浮游植物色素含量[1-2]、增加耐酸性基因表達(dá)[3]、影響光合生理過(guò)程、抑制生長(zhǎng),進(jìn)而影響群落結(jié)構(gòu)組成[4-5]。前期的研究表明,新安江流域浮游植物多樣性及豐度在短期酸脅迫下整體呈降低趨勢(shì),且不同門類浮游植物的敏感性不同[6],這可能與種間/株系間差異性有關(guān)[7]。
淡水水體因溶解有機(jī)質(zhì)含量、水體pH值等因素的影響,水體透光率存在較大差異,從而使到達(dá)水體的光質(zhì)及光強(qiáng)發(fā)生改變。對(duì)浮游植物光合作用及初級(jí)生產(chǎn)過(guò)程來(lái)說(shuō),陽(yáng)光光譜中可見(jiàn)光(P,400~700 nm)與紫外光(UVR,包括320~400 nm紫外線A(UVA)和280~320 nm紫外線B(UVB))是最為重要的部分。作為水質(zhì)全年總體較優(yōu)(Ⅰ類或Ⅱ類以上占比較高)的水系(濁度較低)[8-9],長(zhǎng)期酸雨背景下將進(jìn)一步提高水體透明度,從而增加可見(jiàn)光(P)及紫外輻射(UVR)在水中的強(qiáng)度與透射深度。UVR可單獨(dú)或與高光強(qiáng)耦合降低浮游植物的光合速率[10],抑制營(yíng)養(yǎng)鹽吸收[11-12],誘導(dǎo)細(xì)胞DNA、RNA突變以及活性氧自由基的產(chǎn)生[13-14],導(dǎo)致生產(chǎn)力的降低[12]。針對(duì)新安江流域(屯溪段)浮游植物群落結(jié)構(gòu)與環(huán)境因子的研究也表明,陽(yáng)光輻射尤其是UVR(UVA和UVB)為調(diào)控浮游植物豐度的重要因子[9]。
黃山市境內(nèi)新安江流域?yàn)槲覈?guó)首個(gè)跨省生態(tài)補(bǔ)償試點(diǎn)區(qū),具有重要生態(tài)意義與示范作用。在我國(guó)重要河流、湖泊水質(zhì)逐步好轉(zhuǎn)這一背景下,酸雨脅迫與UVR耦合對(duì)浮游植物生理、生態(tài)過(guò)程的影響,在未來(lái)將是我國(guó)淡水生態(tài)系統(tǒng)面臨的重要環(huán)境問(wèn)題之一。雖然在一定時(shí)期內(nèi)酸雨不會(huì)導(dǎo)致水體pH值降低至酸雨臨界點(diǎn)(pH值5.65)或更低的水平,然而,長(zhǎng)期、持續(xù)酸脅迫下,伴隨水體酸堿緩沖能力(acid-neutralizing capacity, ANC)的降低,流域水體在未來(lái)將面臨低pH的脅迫。因此,研究酸雨脅迫如何與UVR耦合影響新安江流域水體初級(jí)生產(chǎn)者,是亟需探討的重要科學(xué)問(wèn)題。前期的調(diào)查表明綠藻門為新安江流域主要優(yōu)勢(shì)門類(硅藻門、藍(lán)藻門次之)[6, 9],小球藻作為新安江流域重要優(yōu)勢(shì)種類,出現(xiàn)頻率高、豐度占比高[9]。本研究選取流域分離獲得的一株小球藻(Chlorellavulgaris)為對(duì)象,通過(guò)模擬實(shí)驗(yàn),研究了該種生長(zhǎng)及光合活性對(duì)短期模擬酸雨脅迫引起的水體pH值降低的響應(yīng),并探討了水體酸化與短期UVR的耦合效應(yīng)。
1 材料與方法(Materials and methods)
1.1 藻種及培養(yǎng)條件
本實(shí)驗(yàn)所用小球藻(Chlorellavulgaris)于2013年7月分離自安徽省黃山市休寧縣汊口鄉(xiāng)一小型人工水池。藻種使用BG-11(+N)培養(yǎng)基進(jìn)行培養(yǎng),并長(zhǎng)期保種于光照培養(yǎng)箱內(nèi),溫度為20 ℃,光照強(qiáng)度為60 μmol·m-2·s-1,光暗比(L∶D)為12 h∶12 h。
1.2 實(shí)驗(yàn)方法
1.2.1 模擬酸雨母液制備及培養(yǎng)基pH值測(cè)定
人工調(diào)配質(zhì)量分?jǐn)?shù)為98%的濃硫酸(國(guó)藥集團(tuán)化學(xué)試劑有限公司)與質(zhì)量分?jǐn)?shù)為65%的濃硝酸(國(guó)藥集團(tuán)化學(xué)試劑有限公司),按物質(zhì)的量比為8∶1制備模擬酸雨母液(黃山地區(qū)以硫酸型酸雨為主)。通過(guò)向培養(yǎng)基中加入不同體積的模擬酸雨母液,分別獲得pH值為5.65和4.50的模擬酸雨培養(yǎng)基,正常培養(yǎng)基使用1 molL-1的HCl和NaOH調(diào)節(jié)pH值為7.10作為對(duì)照組。模擬酸雨制備過(guò)程中使用Mettler Toledo DL15 Titrator(瑞典)對(duì)pH值進(jìn)行測(cè)定,測(cè)定前使用National Bureau of Standards(NBS)緩沖液對(duì)pH計(jì)進(jìn)行校正。
1.2.2 模擬酸雨處理及細(xì)胞粒徑、比生長(zhǎng)速率(μ)的測(cè)定
通過(guò)向已調(diào)節(jié)好pH值(7.10、5.65和4.50)的培養(yǎng)基中加入指數(shù)生長(zhǎng)期的藻細(xì)胞,調(diào)節(jié)細(xì)胞濃度至約1.0×105個(gè)mL-1,于體積為1 L的聚碳酸酯瓶(PC)中培養(yǎng),每個(gè)處理下3個(gè)重復(fù)培養(yǎng)。將上述不同pH處理的小球藻藻體置于高(150 μmol·m-2·s-1)、低(60 μmol·m-2·s-1,即培養(yǎng)光強(qiáng))光強(qiáng)下進(jìn)行短期培養(yǎng)(24 h),光暗比(L∶D)為12 h∶12 h,培養(yǎng)溫度為20 ℃。使用顆粒計(jì)數(shù)儀(Z2, Beckman,美國(guó))測(cè)定上述條件下培養(yǎng)24 h后的細(xì)胞濃度及細(xì)胞粒徑,并對(duì)各處理下的pH值進(jìn)行測(cè)定獲得其變化情況。
比生長(zhǎng)速率(μ)的計(jì)算使用公式:
μ(h-1)=(lnNA-lnNB)/t
式中:NA為各處理下培養(yǎng)24 h(t)后的細(xì)胞濃度(個(gè)·mL-1),NB為初始接種的細(xì)胞濃度(個(gè)·mL-1)。
1.2.3 色素的提取和測(cè)定
各pH值處理24 h后,將一定體積的藻液經(jīng)GF/F濾膜(0.7 μm, Whatman)過(guò)濾,置于離心管內(nèi)并加入5 mL甲醇,4 ℃條件下過(guò)夜提取,于離心機(jī)內(nèi)5 000g下離心10 min,分光光度計(jì)(Beckman DU-800,美國(guó))下測(cè)定200~750 nm吸光值(Anm)。葉綠素a(Chla)、葉綠素b(Chlb)及類胡蘿卜素(carotenoid)的測(cè)定方法參照Wellburn[15],紫外吸收物質(zhì)(MAAs)的測(cè)定依據(jù)提取液上清吸收光譜在334 nm處的吸收峰高度(HOD)[16],以HOD與Chla的比值獲得,各種色素的具體計(jì)算方法如下:
Chla(μg·mL-1)=15.65×(A666-A750)-7.34×(A653-A750)
Chlb(μg·mL-1)=27.05×(A653-A750)-11.21×(A666-A750)
Carotenoid (μg·mL-1)=(1000×(A470-A750)-2.86×Chla-129.2×Chlb)/221
MAAs (HOD·ng-1)=H/Chla
1.2.4 紫外輻射處理
將上述不同pH值、高低光強(qiáng)下培養(yǎng)24 h后的小球藻置于35 mL石英管內(nèi),通過(guò)在石英管外包裹不同的濾光膜獲得2種輻射處理,即只有可見(jiàn)光輻射(P)(Ultraphan 395濾膜,UV Opak,Digefra,德國(guó)),或者全波段輻射(PAB,可見(jiàn)光+紫外線A+紫外系B)(Ultraphan 295濾膜,UVOpak,Digefra,德國(guó))。本研究輻射處理使用太陽(yáng)模擬器(Sol 1200 W,A. G. H?nle,Martinsried,德國(guó))進(jìn)行,該光源可獲得穩(wěn)定的輻射處理,其中P的強(qiáng)度為87.5 W·m-2,UVA的強(qiáng)度為33.5 W·m-2,UVB的強(qiáng)度為1.91 W·m-2(參考我國(guó)南方地區(qū)夏季UVB輻射強(qiáng)度設(shè)置)。
1.2.5 葉綠素?zé)晒鈪?shù)測(cè)定
葉綠素?zé)晒鈪?shù)使用Xe-PAM(Walz,德國(guó))進(jìn)行測(cè)定。小球藻經(jīng)不同pH處理0、0.5、1、3、8、20和24 h后,對(duì)其有效光化學(xué)效率(Yield)進(jìn)行時(shí)間序列測(cè)定。轉(zhuǎn)移至陽(yáng)光模擬器下進(jìn)行短期輻射處理,測(cè)定經(jīng)P和PAB輻射處理1 h后的Yield和光響應(yīng)曲線。光響應(yīng)曲線共設(shè)置8個(gè)光強(qiáng)梯度,即0、156、226、337、533、781、1 077、1 593和2 130 μmol·m-2·s-1,其中飽和脈沖強(qiáng)度為5 000 μmol·m-2·s-1,持續(xù)時(shí)間為0.8 s。
Yield的計(jì)算公式為:
Yield=(F’m-F)/F’m
式中:Yield為光系統(tǒng)Ⅱ的有效光化學(xué)效率,F(xiàn)為光適應(yīng)下的實(shí)際熒光值,F(xiàn)’m為光適應(yīng)下的最大葉綠素?zé)晒狻?/p>
光響應(yīng)曲線相對(duì)電子傳遞速率(rETR)通過(guò)以下的公式計(jì)算:
rETR=Yield×0.5×PFD
式中:0.5代表光系統(tǒng)Ⅱ吸收的光量子占總量的50%,PFD為光化光的強(qiáng)度(μmol·m-2·s-1)。
快速光響應(yīng)曲線的擬合參照J(rèn)assby和Platt[17]的方法,使用如下公式進(jìn)行:
Y=rETRmax×tanh (α×x/rETRmax)
式中:x表示為光強(qiáng),Y為rETR。
通過(guò)擬合可獲得相對(duì)最大電子傳遞速率(rETRmax)、光能利用效率(α),以rETRmax與α的比值計(jì)算得到飽和光強(qiáng)(Ik)。
經(jīng)陽(yáng)光模擬器下處理后,相對(duì)于初始狀態(tài)的Yield,由P以及PAB導(dǎo)致的抑制率,計(jì)算方式如下:
式中:InhP、InhPAB表示在陽(yáng)光模擬器下經(jīng)P和PAB輻射處理1 h后相對(duì)于PInitial(初始Yield值)的抑制率,PP和PPAB表示P和PAB下1 h后的Yield值。
1.2.6 數(shù)據(jù)統(tǒng)計(jì)分析
不同pH值處理之間的顯著性使用one-way ANOVA(Turkey)分析;培養(yǎng)光強(qiáng)、pH值以及輻射處理兩兩之間以及三者之間的交互效應(yīng)使用Two/Three-way ANOVA分析(P<0.05)。數(shù)據(jù)處理及分析使用Prism 9.0和SPSS 26.0軟件進(jìn)行。
2 結(jié)果(Results)
2.1 培養(yǎng)體系pH值變動(dòng)情況
相對(duì)于初始pH值,經(jīng)24 h培養(yǎng)后,各處理下pH值均有一定升高,高光(HL)下pH值7.10、5.65和4.50處理,pH值分別升高了4.74%、9.20%和4.30%,低光(LL)下分別升高了3.19%、7.49%和2.59%(表1)。
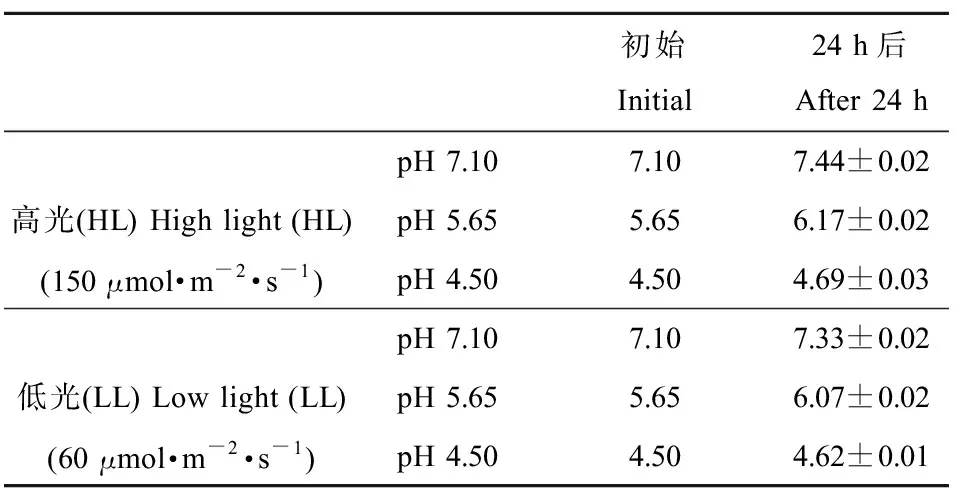
表1 不同處理下初始和培養(yǎng)24 h后的pH值Table 1 pH values at the beginning and after 24 h culture under different treatments
2.2 細(xì)胞粒徑及比生長(zhǎng)速率(μ)
多因素方差分析表明,pH值處理對(duì)小球藻的μ產(chǎn)生顯著影響(P<0.05)。pH 5.65下,低光處理與對(duì)照組μ無(wú)顯著差異,高光處理μ顯著升高了79.6%(P<0.05)(圖1(a));較低的pH值(4.50)細(xì)胞出現(xiàn)負(fù)增長(zhǎng),相對(duì)于pH值7.10處理,高、低光強(qiáng)下分別降低了223%(P<0.05)和225%(P<0.05)(圖1(a))。

圖1 小球藻經(jīng)正常pH值(7.10)以及模擬酸雨脅迫(pH值5.65和4.50)處理,轉(zhuǎn)移至高光(150 μmol·m-2·s-1)和低光(60 μmol·m-2·s-1)下培養(yǎng)24 h后的比生長(zhǎng)速率(μ)(a)和平均細(xì)胞粒徑(b)(n=3)Fig. 1 Specific growth rate (μ) (a) and mean cell size (b) of Chlorella vulgaris cultured under high (150 μmol·m-2·s-1) and low (60 μmol·m-2·s-1) light conditions for 24 h in control pH (7.10) and acid stress treatments (pH 5.65 and 4.50) (n=3)
多因素方差分析表明,光強(qiáng)與pH值處理均單獨(dú)或交互對(duì)細(xì)胞粒徑產(chǎn)生顯著影響(P<0.05)。各pH值處理下高、低光強(qiáng)間細(xì)胞粒徑無(wú)顯著差異(P>0.05);相較于對(duì)照組(7.10),低pH值下(4.50)細(xì)胞粒徑在高光培養(yǎng)下顯著降低了5.5%(P<0.05),低光下各pH值處理間無(wú)顯著差異(P>0.05)(圖1(b))。
2.3 色素含量變化
多因素方差分析表明,光強(qiáng)而非pH值對(duì)光合色素(葉綠素a、b)及光保護(hù)色素(類胡蘿卜素)含量產(chǎn)生了顯著影響(均P<0.05);光強(qiáng)與pH值協(xié)同對(duì)葉綠素a、葉綠素b和類胡蘿卜素含量無(wú)顯著交互效應(yīng)(均P>0.05)(圖2)。不同pH值處理24 h后,高光強(qiáng)培養(yǎng)的藻體,單位細(xì)胞的葉綠素a(圖2(a))、葉綠素b(圖2(b))都顯著低于低光培養(yǎng)的藻體(P<0.05);類胡蘿卜素含量在高、低光強(qiáng)處理間無(wú)顯著差異(P>0.05)(圖2(c));高光強(qiáng)下,類胡蘿卜素與葉綠素a的比值顯著高于低光處理組(P<0.05)(圖2(d));MAAs在高、低光強(qiáng)下無(wú)顯著差異(P>0.05)(圖2(e))。高、低光強(qiáng)處理下,葉綠素a、葉綠素b、類胡蘿卜素、類胡蘿卜素與葉綠素a的比值以及MAAs在各pH值處理間無(wú)顯著差異(P>0.05)。
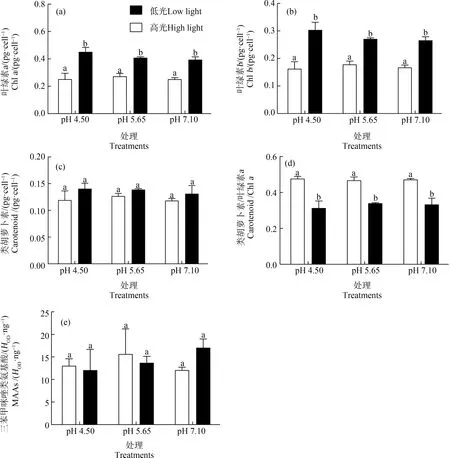
圖2 小球藻經(jīng)正常pH值(7.10)以及模擬酸雨脅迫(pH值5.65和4.50)處理,轉(zhuǎn)移至高光(150 μmol·m-2·s-1)和低光(60 μmol·m-2·s-1)下培養(yǎng)24 h后的葉綠素a(a)、葉綠素b(b)、類胡蘿卜素(c)、類胡蘿卜素與葉綠素a的比值(d)以及三苯甲咪唑類氨基酸(MAAs)(e)(n=3)注:MAAs的濃度以單位葉綠素a的質(zhì)量計(jì)。Fig. 2 Chlorophyll a (a), chlorophyll b (b), carotenoid (c), the ratio of carotenoid to chlorophyll a (d) and mycosporinie-like amino acids (MAAs) (e) of Chlorella vulgaris cultured under high (150 μmol·m-2·s-1) and low (60 μmol·m-2·s-1) light conditions for 24 h in control pH (7.10) and acid stress treatments (pH 5.65 and 4.50) (n=3)Note: The concentration of MAAs is expressed as mass per unit Chl a.
2.4 有效光化學(xué)效率(Yield)及快速光響應(yīng)曲線
經(jīng)模擬酸雨處理后,對(duì)各處理下藻體Yield進(jìn)行時(shí)間序列測(cè)定。相對(duì)于初始Yield,高、低光強(qiáng)下藻體Yield隨處理時(shí)間的增加都呈增加趨勢(shì)(圖3)。高光培養(yǎng)下,相對(duì)于pH值7.10處理,低pH值處理(4.50)的藻體,Yield隨時(shí)間呈先升高后降低趨勢(shì),經(jīng)模擬酸雨處理8、20和24 h后,Yield相對(duì)于pH值7.10處理顯著降低了6.77%、4.69%和2.61%(P<0.05)(圖3(a));低光處理下,各pH處理間在一系列時(shí)間序列下都無(wú)顯著性差異(P>0.05)(圖3(b))。多因素方差分析顯示,不同培養(yǎng)光強(qiáng)對(duì)各pH值處理下藻體Yield具有顯著影響(P<0.05)。對(duì)比高、低光強(qiáng)處理各pH值下的Yield,整體上低光培養(yǎng)的藻體具有高的Yield(圖3(a)和圖3(b))。
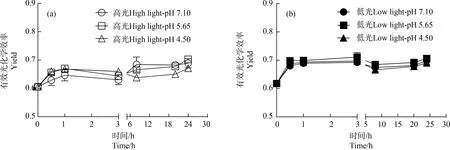
圖3 小球藻經(jīng)正常pH值(7.10)以及模擬酸雨脅迫(pH值5.65和4.50)處理,轉(zhuǎn)移至高光(150 μmol·m-2·s-1)(a)和低光(60 μmol·m-2·s-1)(b)下培養(yǎng)24 h過(guò)程中的有效光化學(xué)效率(Yield)(n=3)Fig. 3 The effective quantum yield (Yield) of control pH (7.10) and acid stress (pH 5.65 and 4.50) treated Chlorella vulgaris during the 24 h culture under high (150 μmol·m-2·s-1) and low (60 μmol·m-2·s-1) light intensity (n=3)
置于陽(yáng)光模擬器下輻射處理1 h后,與培養(yǎng)光強(qiáng)相比,Yield在各pH值下都顯著受到抑制,且抑制程度在PAB處理下更高(P<0.05)(圖4)。多因素方差分析表明,培養(yǎng)光強(qiáng)、輻射處理(P、PAB)以及pH處理都顯著影響了藻體的Yield(P<0.05),且培養(yǎng)光強(qiáng)、輻射處理及pH處理兩兩之間以及三者之間對(duì)Yield均產(chǎn)生了交互影響(P<0.05)。高光培養(yǎng)下的藻,相對(duì)于pH值7.10處理,在pH值4.50下經(jīng)P、PAB輻射處理后,Yield顯著降低了14.3%(P<0.05)和21.4%(P<0.05)(圖4(a));而低光下培養(yǎng)的藻,經(jīng)陽(yáng)光模擬器下輻射處理后(PAB),Yield在低pH值(5.65、4.50)下整體低于高光處理組,相較于pH值7.10處理,pH值4.50下Yield顯著降低了20.8%(P<0.05)(圖4(b))。高光強(qiáng)培養(yǎng)的藻體,UVR誘導(dǎo)的Yield抑制率在各pH處理間無(wú)顯著差異(P>0.05);而低光下培養(yǎng)的藻,pH值4.50下UVR誘導(dǎo)的抑制率在所有處理間最高,相較于pH值7.10處理升高了53.2%(P<0.05)(圖4(c))。
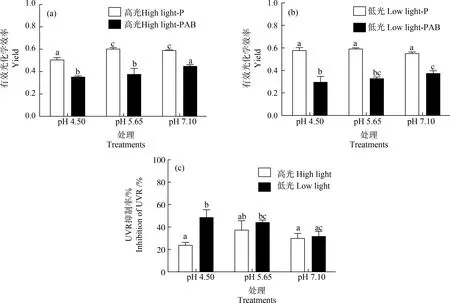
圖4 小球藻經(jīng)正常pH值(7.10)以及模擬酸雨脅迫(pH值5.65和4.50)處理,轉(zhuǎn)移至高光(150 μmol·m-2·s-1)(a)和低光(60 μmol·m-2·s-1)(b)下培養(yǎng)24 h后,置于陽(yáng)光模擬器下給予可見(jiàn)光(P)和可見(jiàn)光+紫外線A+紫外線B(PAB)輻射處理1 h的Yield和UVR誘導(dǎo)的抑制率(c) (n=3)Fig. 4 The Yield (a), (b) and its inhibition rate induced by UVR (c) of control pH (7.10) and acid stress (pH 5.65 and 4.50) treated Chlorella vulgaris that have been cultured under high (150 μmol·m-2·s-1) and low (60 μmol·m-2·s-1) light intensity for 24 h when exposure with P and PAB for 1 h (n=3)
對(duì)比高、低培養(yǎng)光強(qiáng)處理下UVR導(dǎo)致的抑制率,pH值4.50處理下,相對(duì)于高光培養(yǎng)的藻,UVR誘導(dǎo)的抑制率在低光強(qiáng)處理下增加了62.1%(P<0.05);其余pH值處理下,高、低光強(qiáng)間無(wú)顯著差異(P>0.05)(圖4(c))。多因素方差分析表明,培養(yǎng)光強(qiáng)以及pH值處理都可顯著影響藻體對(duì)UVR的響應(yīng)(抑制率),且二者具有交互影響(P<0.05)。
總體上,pH值處理未對(duì)藻體rETRmax、α和IK產(chǎn)生影響(均P>0.05,three-Way ANOVA);而紫外輻射顯著影響了藻體的rETRmax、α和IK(均P<0.05,three-Way ANOVA)(圖5和表2)。相較于P處理,PAB處理下rETRmax、α顯著降低,IK呈升高趨勢(shì);培養(yǎng)光強(qiáng)與紫外輻射耦合顯著影響了α(P<0.05)和IK(P<0.05)(表2)。高光培養(yǎng)下的藻體經(jīng)PAB輻射處理后,rETRmax、α隨pH值降低呈降低趨勢(shì);低光培養(yǎng)下的藻體,經(jīng)PAB輻射處理后,rETRmax、α和IK在各pH值間無(wú)顯著差異(P>0.05),然而UVR誘導(dǎo)的rETRmax和α的抑制率在低光處理下整體高于高光培養(yǎng)的藻體(表2)。

圖5 小球藻在正常pH值(7.10)以及模擬酸雨脅迫(pH值5.65和4.50)處理后轉(zhuǎn)移至高光(150 μmol·m-2·s-1)(a)和低光(60 μmol·m-2·s-1)(b)下培養(yǎng)24 h后,置于陽(yáng)光模擬器下給予可見(jiàn)光(P)和可見(jiàn)光+紫外線A+紫外線B(PAB)輻射處理1 h的快速光響應(yīng)曲線(n=3)Fig. 5 The rapid light curve of control pH (7.10) and acid stress (pH 5.65 and 4.50) treated Chlorella vulgaris that have been cultured under high (150 μmol·m-2·s-1) (a) and low (60 μmol·m-2·s-1) (b) light intensity for 24 h, exposure with P and PAB for 1 h (n=3)
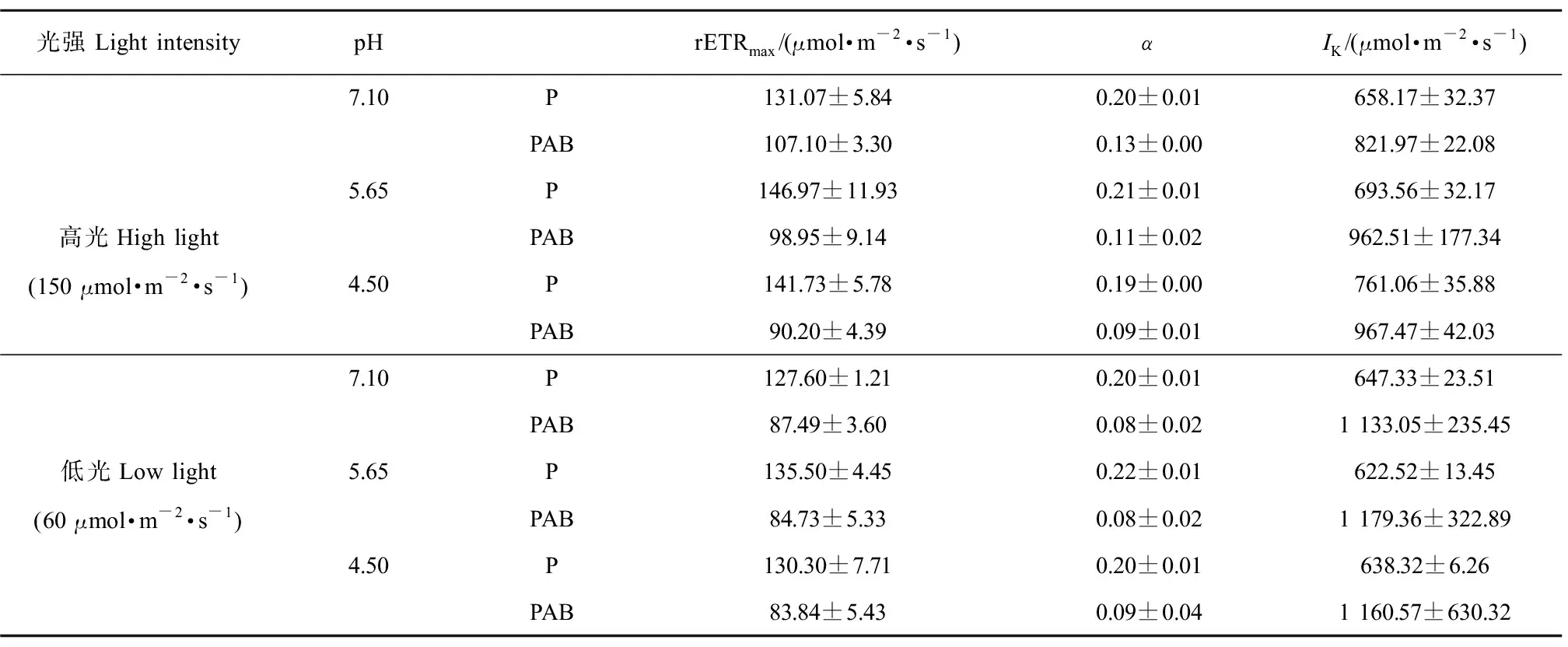
表2 根據(jù)圖5快速光響應(yīng)曲線計(jì)算得到的不同光強(qiáng)水平、pH值下培養(yǎng)的小球藻經(jīng)可見(jiàn)光(P)和可見(jiàn)光+紫外線A+紫外線B(PAB)輻射處理1 h的最大相對(duì)電子傳遞速率(rETRmax)、光能利用效率(α)和飽和光強(qiáng)(IK)(n=3)Table 2 The calculated maximum relative electron transport rate (rETRmax), light using efficiency (α) and saturation light intensity (IK) of Chlorella vulgaris that acclimated under high and low light intensity with different pH treatments, when exposure with P and PAB for 1 h, the calculation was according to figure 5 (n=3)
3 討論(Discussion)
對(duì)浮游植物光合生理過(guò)程以及群落結(jié)構(gòu)組成產(chǎn)生調(diào)節(jié)效應(yīng)的諸多生態(tài)因子中,水體pH值和光照(光質(zhì)、光強(qiáng))是最為主要的調(diào)節(jié)因子。小球藻是綠藻門中分布較為廣泛(淡、咸水水體)的世界性普生種類[18],適應(yīng)與耐受環(huán)境脅迫能力較強(qiáng),也是新安江流域的重要優(yōu)勢(shì)種類[9],探討流域重要環(huán)境脅迫(酸雨與UVR)對(duì)其光合生理的影響,具有重要生態(tài)學(xué)意義。本研究結(jié)果表明小球藻對(duì)模擬酸雨引起的短期pH值降低有一定耐受能力,生長(zhǎng)及色素對(duì)pH值降低的響應(yīng)相較于培養(yǎng)光強(qiáng)影響的響應(yīng)要低,低光、較低pH值(4.50)下培養(yǎng)的藻體光合過(guò)程對(duì)高光強(qiáng)、紫外輻射的敏感性增加。
酸雨的生態(tài)學(xué)效應(yīng)在陸生植物中已開(kāi)展了較多的研究[19-20],酸雨引起的水體酸化對(duì)水生藻類的影響也有了一定的認(rèn)識(shí)[21-23]。以往的研究表明,酸雨脅迫引起的水體短期或長(zhǎng)期酸化可改變浮游[6, 24]或底棲生物群落結(jié)構(gòu)組成[21]。酸雨引起的水體酸化其效應(yīng)主要包括水體酸堿緩沖能力降低(pH值下降)和無(wú)機(jī)碳濃度、形式改變[25],因不同門類、屬種的浮游植物的敏感程度(無(wú)機(jī)碳親和力、酸堿耐受能力)不同[26],響應(yīng)水體酸化的光合生理過(guò)程也存在顯著的種間或株系間差異[6-7]。本研究短期模擬酸雨臨界點(diǎn)pH值(5.65)對(duì)小球藻生長(zhǎng)、細(xì)胞粒徑大小及色素含量均未產(chǎn)生顯著影響,相較于藍(lán)藻門銅綠微囊藻產(chǎn)毒株系905以及不產(chǎn)毒株系469[7],小球藻對(duì)模擬酸雨引起的水體酸化耐受性較高、敏感性較低。這種響應(yīng)模擬酸雨脅迫引起水體pH降低的種間差異性表現(xiàn),在酸雨控制區(qū)內(nèi)可能成為水體浮游植物群落結(jié)構(gòu)組成的主要調(diào)節(jié)因素。浮游植物光合色素含量可受光強(qiáng)變動(dòng)的調(diào)節(jié)[27-28],本研究也表明光合色素(Chla、Chlb)受培養(yǎng)光強(qiáng)的影響較大、響應(yīng)較快,而光合及光保護(hù)色素(類胡蘿卜素、MAAs)對(duì)短期pH值變化的響應(yīng)不敏感,整體上未受光強(qiáng)或pH值影響。光保護(hù)色素未受培養(yǎng)光強(qiáng)升高的影響,表明本研究設(shè)置的高光培養(yǎng)(150 μmol·m-2·s-1)尚未達(dá)到其飽和光強(qiáng),無(wú)需通過(guò)光保護(hù)色素的合成應(yīng)對(duì)光脅迫。
水體酸化引起的H+濃度升高,酸堿耐受能力不同的藻類(胞內(nèi)酸堿平衡調(diào)控),其能量代謝過(guò)程(光合放氧、固碳、光呼吸及暗呼吸等過(guò)程)也將受到不同程度的影響,進(jìn)而引起差異性的光合特性與生長(zhǎng)表現(xiàn)[29]。水體H+增加(CO2濃度升高引起)與高光強(qiáng)耦合可顯著抑制浮游植物的光合效率、誘導(dǎo)高的非光化學(xué)淬滅(NPQ)從而抑制藻類生長(zhǎng)[30-31]。本研究中也發(fā)現(xiàn)低pH/高H+與高的培養(yǎng)光強(qiáng)耦合,對(duì)藻體光化學(xué)效率產(chǎn)生協(xié)同抑制效應(yīng),表明較低的pH下藻體對(duì)光強(qiáng)的敏感性增加、光能利用效率降低。
以往研究表明UVR可通過(guò)損傷光系統(tǒng)光合元件(例如,光系統(tǒng)II關(guān)鍵蛋白D1、PsbA和PsbD等)[32-34]、誘導(dǎo)活性氧自由基等[35]降低藻體光合活性。此外,UVR與酸雨脅迫耦合可進(jìn)一步降低浮游植物的光合能力,且這一影響存在種間/株系間差異[7]。本研究中UVR對(duì)Yield、最大相對(duì)電子傳遞速率(rETRmax)和光能利用效率(α)的抑制率在低pH值(4.50)、低光培養(yǎng)下的藻體中更為顯著,藻體對(duì)高光及紫外輻射的敏感性增加,這可能與低光培養(yǎng)下藻體相對(duì)較低的carotenoid/Chla以及光系統(tǒng)Ⅱ高的損傷、低的修復(fù)速率有關(guān)[34]。這一結(jié)果表明藻細(xì)胞經(jīng)歷的光環(huán)境(高、低光強(qiáng))可顯著影響水體酸化與UVR的耦合效應(yīng)。
根據(jù)《中國(guó)生態(tài)環(huán)境狀況公報(bào)》公布的近30年的觀測(cè)數(shù)據(jù),我國(guó)受酸雨影響的城市依然處于較高的比例(例如,2020年度發(fā)生酸雨的城市占所有監(jiān)測(cè)城市的34.0%),酸雨脅迫及其生態(tài)學(xué)效應(yīng)在未來(lái)很長(zhǎng)一段時(shí)間仍將是我國(guó)面臨的重要環(huán)境問(wèn)題之一。淡水水體,尤其是河流生態(tài)系統(tǒng),水體光環(huán)境(光強(qiáng)、光質(zhì))受水深、流速、濁度以及降雨量等因素影響,并存在季節(jié)變化特征。酸雨控制區(qū)不同門類、屬種的浮游植物如何響應(yīng)與適應(yīng)pH值與光環(huán)境的變化將對(duì)河流浮游植物的群落結(jié)構(gòu)組成與變動(dòng)起到重要調(diào)節(jié)作用。小球藻作為新安江流域主要優(yōu)勢(shì)種類,對(duì)短期水體酸化有一定耐受能力,然而,氣候變化背景下其光合生理過(guò)程如何響應(yīng)與適應(yīng)未來(lái)長(zhǎng)期、多重環(huán)境脅迫的影響,將決定其在水生態(tài)系統(tǒng)中的競(jìng)爭(zhēng)優(yōu)勢(shì)與結(jié)果。本研究是基于短期模擬酸雨脅迫與UVR耦合效應(yīng)的研究,未來(lái)的研究將通過(guò)開(kāi)展長(zhǎng)期脅迫下的響應(yīng)與適應(yīng)機(jī)制探討,以及通過(guò)對(duì)流域其他門類(藍(lán)藻門、硅藻門等)、類群(例如非優(yōu)勢(shì)種類)進(jìn)行比較生理學(xué)分析,以系統(tǒng)了解酸雨控制區(qū)光強(qiáng)、光質(zhì)如何影響水體浮游植物群落多樣性及演替規(guī)律。
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(shù)(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
知識(shí)經(jīng)濟(jì)·中國(guó)直銷(2016年3期)2016-02-27 16:15:49
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學(xué)低年級(jí)版)(2011年3期)2011-01-01 00:00:00
