西南喀斯特石漠化環境適生植物細根構型及其與細根和根際土壤養分計量特征的相關性
2022-03-10 03:06:52盛茂銀肖海龍王霖嬌
生態學報 2022年2期
關鍵詞:植物
吳 靜, 盛茂銀, 肖海龍, 郭 超, 王霖嬌,*
1 貴州師范大學喀斯特研究院,貴陽 550001 2 國家喀斯特石漠化治理工程技術研究中心,貴陽 550001 3 貴州省喀斯特山地生態環境國家重點實驗室培育基地,貴陽 550001
細根(直徑小于2mm)具有復雜的分支系統,擁有較大的吸收表面積,是根系的重要組成部分。根系是植物與外界進行物質與能量交換的重要器官之一[1—3],根系構型體現根系在土壤中的分布與排列以及植物對土壤養分的利用策略[4],具有一定的可塑性。根系構型的研究主要著力于幾何形態和拓撲結構參數[5],幾何形態反映根系的形態和功能[6],拓撲結構參數反映根系在土壤空間中的分布特征[7]。在不同因素的影響下,根系構型的表達均有不同[8],存在幾何形態和拓撲結構參數的差異。根系構型在一定程度上反映了植物對環境所采取的適應策略。然而,由于不能直觀的了解根系在地下的生長情況及完整根系的獲取難度較大,以往較多的是通過生理指標反映粗根生長,而通過根系構型反映細根在土壤中的分支模式和適應策略較少。
Fitter等[5]提出了兩種分支模式:魚尾形分支模式和叉狀分支模式,比較了不同分支模式的營養競爭能力,發現魚尾形分支更能適應貧瘠的生境,叉狀分支更能適應養分豐富的生境[9- 10]。黃同麗等[11]對喀斯特地區3種灌木根系構型研究發現,根系均為典型的魚尾形分支模式。而蘇樑等[12]對喀斯特峰叢洼地(廣西環江毛南族自治縣的西北部)不同植被恢復階段的4個優勢種根系構型研究發現,根系均為典型的叉狀分支模式。同位于喀斯特地區出現不同的根系(粗根)分支模式,可能是遺傳和生境的影響[13]。細根是植物吸收水分和養分的主要器官,在植物與土壤的物質和能量交換中起著關鍵性的作用[14- 15]。近期的研究表明,細根養分特征受到土壤養分供應的限制[16]。Ladanai等[17]發現,植物細根和土壤間的養分特征具有緊密的相關性。馬玉珠等[18]綜合中國302個細根數據發現,土壤對細根N、P含量的影響較降水和溫度更高。陳曉萍等[19]研究發現,土壤C與細根N含量呈顯著正相關,而細根P的含量受土壤中P供應量的限制。細根養分與土壤養分關系的研究,可以幫助理解植物養分元素對土壤養分的適應策略,利于喀斯特石漠化的治理與恢復[16]。在喀斯特石漠化生態系統中植物養分特征的研究取得了顯著的成效[16]。王璐等[20]研究發現,核桃(Juglansregia)較刺梨(Rosaroxburghii)有更高的N∶P,對土壤養分的歸還能力較高。植株生長方面,劉立斌等認為在喀斯特地區,植物生長主要受P限制[21],而劉娜等認為主要受N限制[22],皮發劍等認為物種同時受N和P的限制[23]。植株養分含量及計量比和生長受到何種元素限制的研究,對反映根系的分支策略及植株適應策略具有重要意義,對于生長限制元素的研究可綜合植株種類、生境等因素進行綜合考慮。以上研究主要是對不同樹種間根系(粗根)構型和植株的養分特征分別對貧瘠土壤的適應策略研究[24],將細根構型和細根、根際土壤養分相結合的研究,特別是不同樹種在西南喀斯特石漠化環境中的細根拓撲結構和細根-根際土壤養分功能是否存在相關性,還不清楚。因此,將細根拓撲結構特征和養分特征綜合用于細根研究能更準確的反映細根構型特征和生態適應策略[25],對生態環境的恢復與治理起著關鍵作用。
西南喀斯特石漠化是我國西南山區一個獨特的生態環境系統,是我國典型的生態脆弱區;該區具有地表破碎、基巖裸露、土壤淺薄、分層復雜、碳酸鈣含量高等特點,給植被恢復帶來很大難度[26]。構樹(Broussonetiapapyrifera)、花椒(Zanthoxylumbungeanum)、刺梨、火棘(Pyracanthafortuneana)是西南喀斯特地區典型的適生植物;其中構樹是喀斯特地區常見的飼料作物,其韌皮纖維可作為造紙的高級材料,花椒、刺梨、火棘是經濟林樹種,通過平整土地,施加底肥進行人工廣泛種植,成為貴州省扶貧開發的特色產業植物。高經濟效益的適生植物是西南喀斯特石漠化生態恢復與治理的生態型樹種,但長期以來主要是針對粗根的研究,基于細根的研究甚少。鑒于此,本文從細根構型、細根養分、根際土壤養分三方面刻畫細根構型的特點及細根構型與養分之間的相關性,分析影響細根構型的主要因子,探討4種適生植物對喀斯特環境的生態適應性。該研究對揭示喀斯特地區植物共存機制具有科學意義,將為揭示細根構型與細根、根際土養分間的耦合關聯提供理論依據,并對該地區生態恢復與治理提供參考。
1 材料與方法
1.1 研究區概況
研究區位于貴州省關嶺-貞豐花江喀斯特高原峽谷石漠化生態建設綜合治理示范區(25°39′13″ N—25°41′00″ N,105°36′30″ E—105°46′30″ E),海拔450—1450m。氣候為南亞熱帶干熱河谷氣候,年均溫為18.4℃,最低氣溫6.6℃,最高氣溫32.4℃;年均降水量為1100mm,降水集中于夏季。土壤以黃壤、黃色石灰土為主,土壤質地粘重、缺乏團粒結構,土壤中鈣、鎂、鐵含量較高,pH值一般在6.5以上。花江示范區石漠化面積廣布,占示范區總面積的73.10%。經過“九五”和“十五”時期的治理,研究區內植被得到了較好的恢復,植被為亞熱帶常綠落葉針闊混交林,主要為馬尾松(Pinusmassoniana)、杉木(Cunninghamialanceolata)、油桐(Verniciafordii)、漆樹(Toxicodendronvernicifluum)、 構樹、花椒、刺梨、火棘等。
1.2 樣品采集與處理
2020年6月底,在示范區內選擇長勢相近且生長狀況良好的4種適生植物構樹、花椒、刺梨、火棘進行隨機取樣,并對地上部分指標(基徑、株高、植株年齡、林下成分)進行調查,統計為確定標準樣株各12株,4種適生植物的基徑:10—15mm、株高:80—100cm、植株年齡:1—1.5a,林下主要以斑茅(Saccharumarundinaceum)、狗尾巴草(Setariaviridis)等雜草為主。根系采用全挖法,在距樣株1.5m范圍內將凋落物、雜草等清理干凈,用鐵鏟、 鋤頭、 撬棍、毛刷等小心清理出所有直徑大于1mm的整株根系,然后選取直徑小于2mm的完整根系,即為細根,裝入有標簽的保鮮袋中,于當天放入2—3℃冰箱中冷藏。取樣過程中盡量避免末端低級根的損失,以保證細根構型的完整性[27]。根際土的采集參照抖土法[28],利用細根采集時余下的根際土壤,除去根系附近較大的石子和動植物殘體等,抖落根系表面的粒土和雜質,采集附著于根際表面4mm以內的根際土[29]。將每株植物基部土壤進行混合后,分別裝入貼有標簽的密封袋中帶回實驗室。
將取出的完整細根用去離子水洗去表面土壤,采用精度分別為0.01mm、1mm 的游標卡尺和卷尺,對各級根系分支前后的直徑、根系內部和外部連接數量、連接長度等進行測量。每級根系分支前后直徑測量3次,并根據Strahler等[30]方法確定根級。將測量后的同一植株的根系充分混合后分成3份,且4種樹種每一樹種各12棵,每棵樹的根際土取3份,細根樣品和根際土樣品各144個。將細根樣品于105℃殺青30min后調至60℃烘干至恒重,用LMP- 800A高速粉碎機粉碎,過0.149 mm篩。根際土自然晾干一個月,除去其中的動植物殘體以及石灰結核和石子等,用石磨棒研磨后過0.149 mm篩,用以化學分析[27]。
1.3 分析方法
1.3.1拓撲參數計算
(1)拓撲指數:采用Fitter[9]的拓撲指數計算方法:
TI=lgA/lgM
(1)
式中,M為根系所有外部連接的總數;A為最長根系通道內部連接的總數。TI=1時,根系為魚尾形分支,TI越接近0.5時,根系為叉狀分支(圖1)。
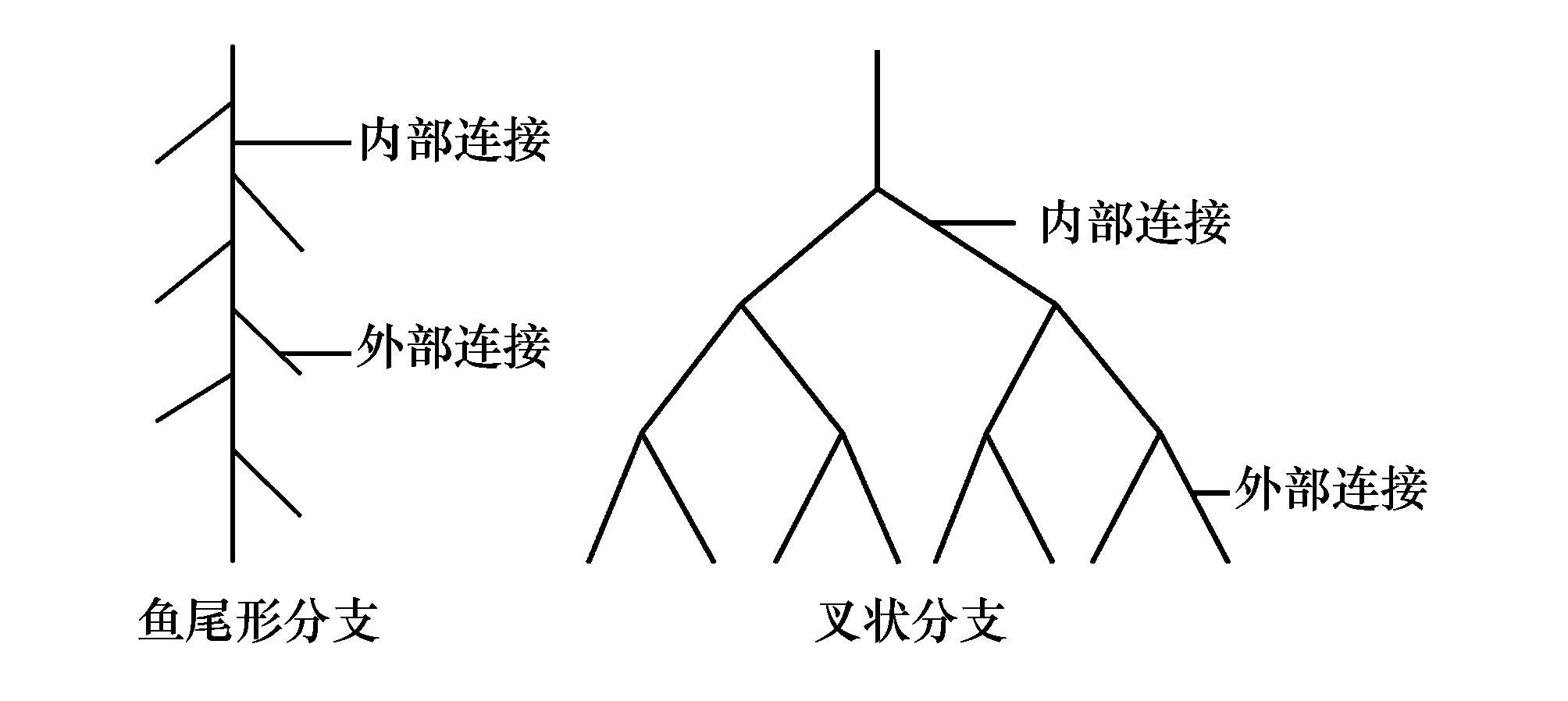
圖1 根系拓撲結構示意圖Fig.1 Schematic of root topological structures
Oppelt等[31]提出了新的修正拓撲參數計算方法:
(2)
式中,a(A)為拓撲長度(植物基部到根終端連接數量),b為平均拓撲長度,Ibv0=lnv0/ln2,b=Pe/v0,v0相當于(1)中的M,Pe為根系基部到根系終端通道的所有連接總數。修正后的拓撲參數qa、qb值的范圍在0—1之間,魚尾形分支qa=qb=1,叉狀分支qa=qb=0,兩種模式的過渡形式是qa、qb的值在0—1之間。
(2)根系分支率
根據Strahler[30]方法確定根級,既由外向內確定根級,最外層的所有根系為一級根,2個一級根相遇為二級根,2個二級根相遇為三級根,以此類推,不同級根系相遇,則以相遇根級中較高級根的級作為相遇后根的根級;由外向內統計不同根級(i)的數量Ni,以等級i為橫坐標,1gNi為縱坐標作圖,植株的總分支率(Rb)由回歸直線斜率的逆對數表示;逐步分支率(Ri:Ri+1)為相鄰兩級根系分支數之比,計算公式為Ri:Ri+1=Ni:Ni+1。
(3)根系分支前后橫截面積
Leonardo da Vinci法則認為植物分支前后的根系橫截面積相等,即分支前的根系橫截面積等于分支后的根系橫截面積總和,用方程式(3)表示:
(3)
通過方程式(3),Van Noordwijk和Purnomosidhi[32]提出了能夠反映普遍根系分支規律的方程(4):
(4)
1.3.2樣品養分測定方法
本研究的實驗方法參考《土壤農化分析》[33],細根、根際土壤各項指標均3次重復測定。其中細根、根際土有機碳(SOC)采用重鉻酸鉀容量法-外加熱法測定,全氮(TN) 采用凱式定氮法測定,全磷(TP)采用H2SO4-H2O2消煮-鉬銻抗比色法。
1.4 數據處理
利用Excel 2016、Canoco 4.5和SPSS 22.0軟件對數據進行統計分析。采用單因素(one-way ANOVA)和LSD法進行方差分析和多重比較(α=0.05),利用Pearson Correlation相關分析檢驗細根構型分別與細根養分、根際土壤養分之間的相關關系;運用主成分分析找出影響細根構型的主要因子。
2 結果與分析
2.1 適生植物細根構型特征
由表1可知,4種適生植物的細根拓撲指數qa、qb、TI均不存在顯著差異,表明在喀斯特石漠化環境中4種適生植物的細根分支模式具有趨同性。其中TI均接近于1,為典型的魚尾形分支模式,該模式具有根系分支簡單,次級分支少,根系內部競爭小等特點。4種適生植物細根總根長較長且存在差異:花椒與火棘不存在顯著差異,其他之間均存在顯著差異。4種適生植物細根總分支率偏小且不存在顯著差異。4種適生植物細根分支前后橫截面積之比存在顯著差異:花椒與火棘存在顯著差異,其他之間均不存在顯著差異,表明構樹、刺梨細根分支前后橫截面積之比均接近于1,基本符合Leonardo da Vinci 法則,體現了構樹、刺梨細根分支前后的橫截面積基本相等。
由圖2可知,4種適生植物細根具有明顯的可塑性與不同的形態特征,可能與樹種和生境條件的不同有關。構樹、刺梨、火棘的細根呈人字形:構樹主根粗壯,側根較長且細,表皮呈土黃色;花椒主根粗短,側根長短差異較大,表皮呈土黃色;刺梨分枝密度不高,側根粗壯,表皮呈黃褐色;火棘主根粗短,側根細長,表皮呈黑色。4種適生植物通過增加根系的連接長度,提升空間拓展能力,以獲得充足的水分與養分供應。
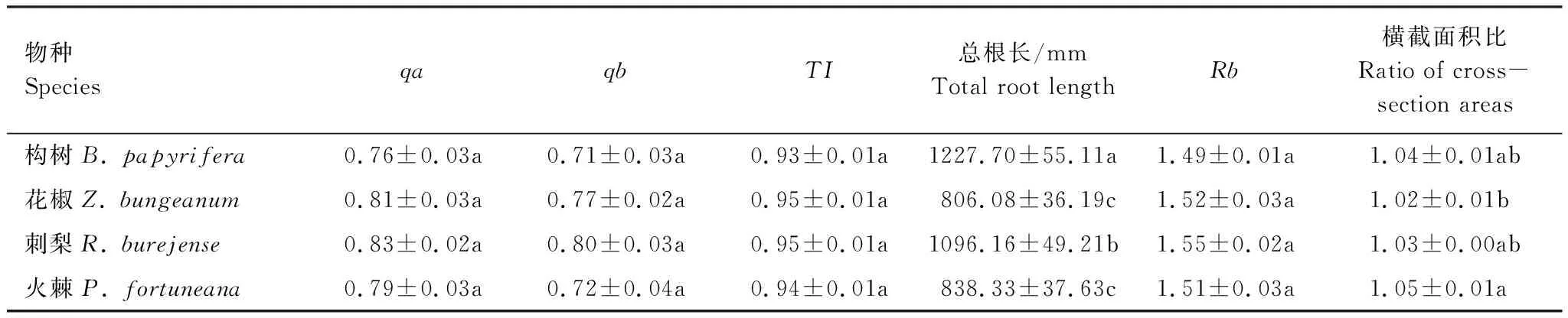
表1 4種適生植物細根構型特征
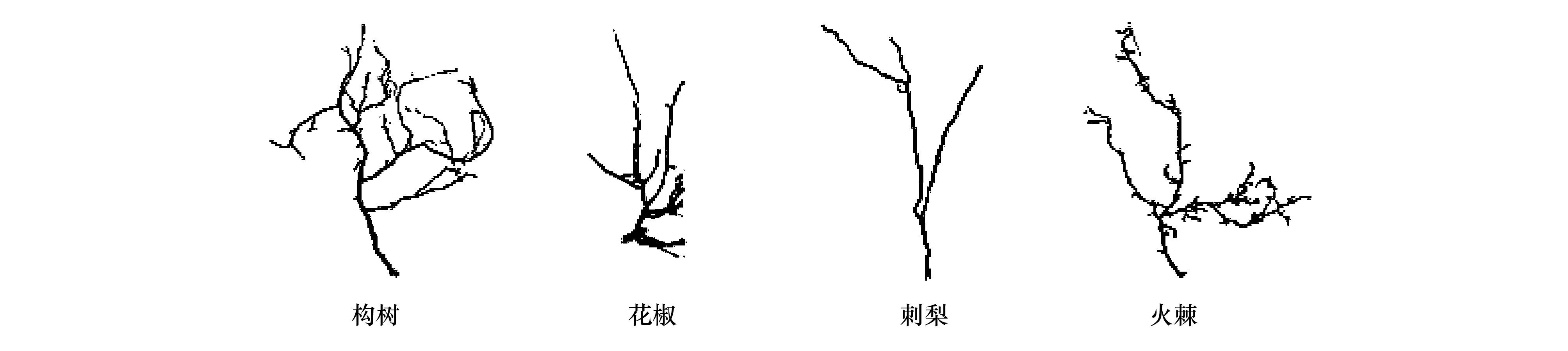
圖2 4種適生植物細根形態特征Fig.2 Fine root sharp of the four adaptive plants
2.2 適生植物細根構型特征與細根C、N、P化學計量的相關性
由表2可知,構樹細根P含量與細根總根長之間存在顯著負相關關系(P<0.05),細根C含量、N含量、C∶N、C∶P、N∶P與細根構型特征之間不存在顯著的相關關系(P>0.05)。花椒細根N∶P與細根總分支率之間存在顯著正相關關系(P<0.05),細根C含量、N含量、P含量、C∶N、C∶P 與細根構型特征之間不存在顯著的相關關系(P>0.05)。
由表2可知,刺梨細根P含量與qa、TI之間存在顯著負相關關系(P<0.05),細根C∶P與qa、qb、TI之間存在顯著正相關關系(P<0.05),細根C含量、N含量、C∶N、N∶P與細根構型特征之間不存在顯著的相關關系(P>0.05)。火棘細根N含量與細根總根長之間存在顯著負相關關系(P<0.05),細根C含量、P含量、C∶N、C∶P、N∶P與細根構型特征之間不存在顯著的相關關系(P>0.05)。
2.3 適生植物細根構型特征與根際土壤C、N、P化學計量的相關性
由表3可知,構樹根際土壤N、P、C∶P、N∶P與細根構型特征之間不存在顯著的相關關系(P>0.05)。根際土壤C含量與細根總分支率之間存在極顯著正相關關系(P<0.01)、與細根橫截面積比之間存在顯著負相關關系(P<0.05)。根際土壤C∶N與細根橫截面積比之間存在顯著負相關關系(P<0.05)。
由表3可知,花椒根際土壤C含量除與細根qa、qb、TI、橫截面積比之間存在顯著正相關關系(P<0.05),與細根總分支率存在顯著負相關關系(P<0.05)。根際土壤N含量與細根橫截面積比存在顯著正相關關系(P<0.05)。根際土壤C∶N與細根qa、qb、TI之間存在顯著正相關關系(P<0.05),根際土壤C∶P與細根總分支率存在顯著負相關關系、與細根橫截面積比存在顯著正相關關系(P<0.05)。根際土壤P含量、N∶P與細根構型特征之間不存在顯著的相關關系(P>0.05)。
由表3可知,刺梨根際土壤C含量與細根總分支率之間存在極顯著負相關關系(P<0.01),根際土壤P含量與細根總分支率之間存在顯著負相關關系(P<0.05),根際土壤C∶P與細根總根長之間存在顯著正相關關系(P<0.05)。根際土壤N含量、C∶N、N∶P與細根構型特征之間不存在顯著的相關關系(P>0.05)。火棘根際土壤養分與細根構型之間不存在顯著的相關關系(P>0.05)。
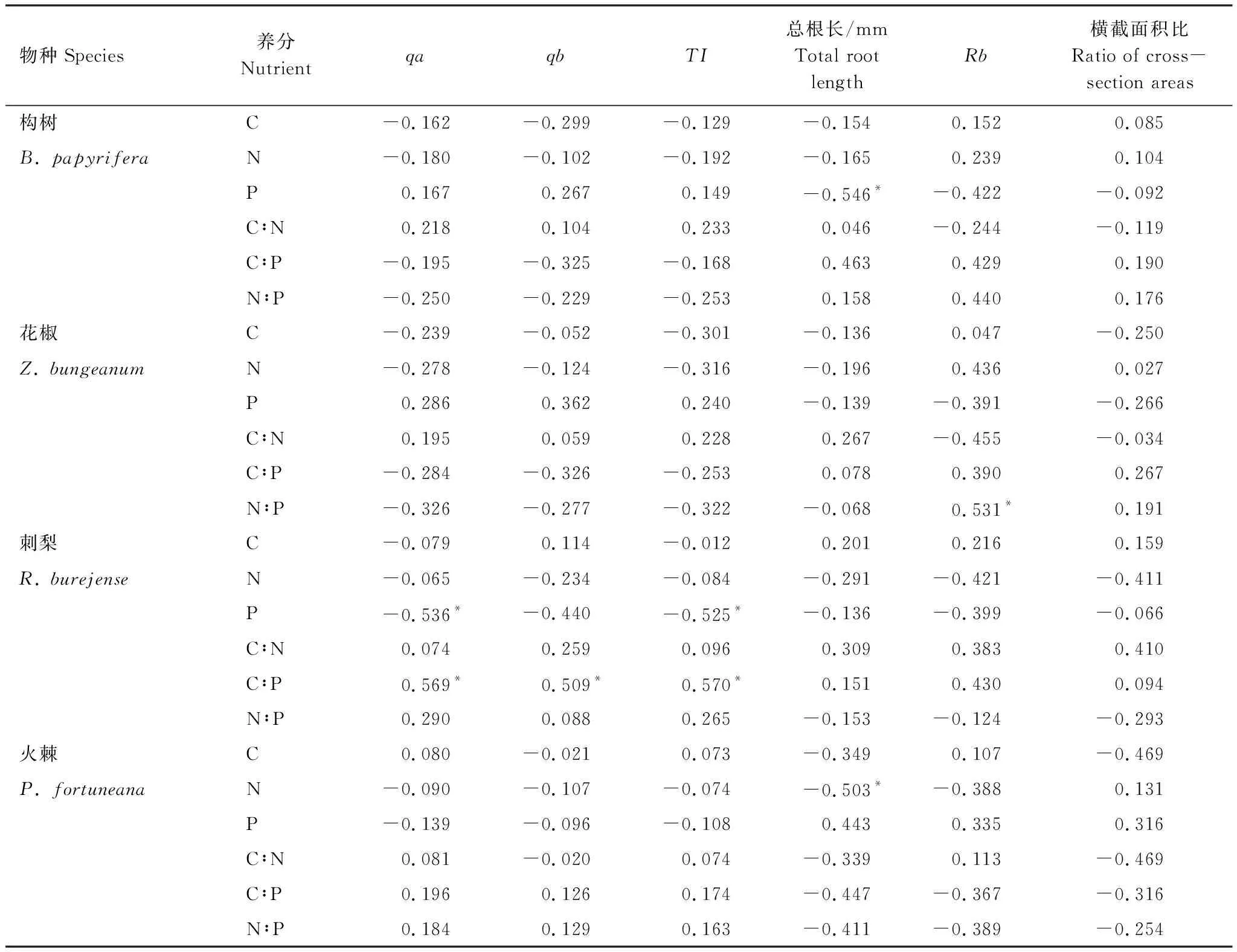
表2 4種適生植物細根構型與細根養分相關系數
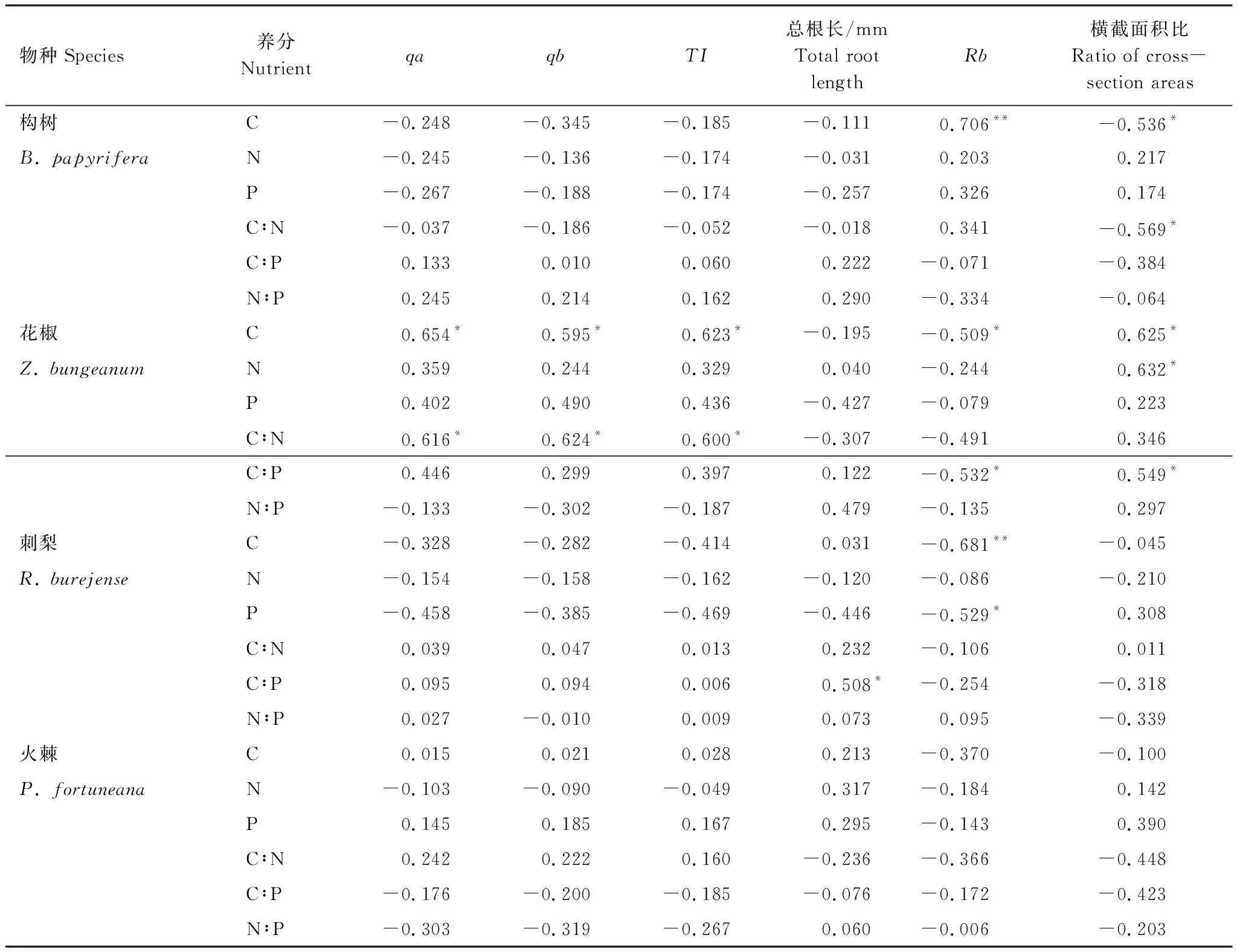
表3 4種適生植物細根構型與根際土壤養分相關系數
2.4 影響細根構型的細根和根際土壤C、N、P化學計量特征主成分分析

圖3 基于細根構型、養分特征的主成分分析Fig.3 Principal component analysis based on fine root architecture and nutrient characteristics rhizosphere soil nutrition 圖中主成分特征根>1;L為總根長;α為橫截面積比;C、N、P、C∶N、C∶P、N∶P分別為細根C、N、P、C∶N、C∶P、N∶P;C1、N1、P1、C∶N1、C∶P1、N∶P1分別為根際土壤C、N、P、C∶N、C∶P、N∶P
通過主成分分析發現(圖3),拓撲指數qa、qb、TI與細根C、N、P含量、根際土壤C∶N、C∶P呈正相關,總根長、橫截面積比與細根C、P含量、細根C∶N、根際土壤C、N含量、根際土壤C∶P、N∶P呈正相關,總分支率與細根C∶P、C∶N、N∶P、根際土壤C、N、P含量、根際土壤N∶P呈正相關。而細根N含量、細根C∶P、N∶P、根際土壤C∶P、N∶P的箭頭較長,表明其對細根構型的影響較大,特別是細根N∶P箭頭最長,說明其對細根構型的影響最大,且細根養分較根際土壤養分對細根構型的影響大。
3 討論
3.1 喀斯特石漠化生境適生植物細根構型
本研究中4種適生植物均為典型的魚尾形分支模式,與黃同麗等[11]對喀斯特地區3種灌木根系研究結果一致。在投入相同的碳時,魚尾形分支利用自身優勢,有效避免根系內部競爭從而迅速占領冗余空間[10],達到吸收養分的目的,因此魚尾形分支更適應于喀斯特獨特的生境[10,34],并能針對生境的變化快速做出適應性的改變[34]。而蘇樑等[12]對喀斯特峰叢洼地不同植被恢復階段的4個優勢種根系構型研究發現均為典型的叉狀分支模式。喀斯特地區出現兩種不同的分支模式,可能與樹種、生境、遺傳有關[13],而分支模式的具體決定因素尚需深入研究。
增加根系的連接長度是植物應對環境變化、擴大分布范圍和提高養分利用效率的重要策略,且養分吸收效率和空間拓展能力與細根連接長度呈正相關[31]。本研究中4種適生植物的細根均為連接長度較長的魚尾形分支,根系通過增加連接長度、降低根系分支率,提高養分吸收效率和空間拓展能力,保證植物的正常生長;這與Fitter[10]和Fitter等[35]發現的根系較高吸收效率與魚尾形分支和較長連接長度相關的結果一致。較長的細根連接長度可能與獨特的生境有關,喀斯特地區純碳酸鹽巖大面積出露,巖溶作用強烈,導致土層淺薄不連續且持水能力低,植物出現“根包石”現象,根系需沿著巖石向下擴張到土壤小生境中,以獲得水分和養分供應。
本研究中4種適生植物的細根總分支率較小,表明細根的生態適應性是大致相同的,即利用較小的分支率來適應喀斯特獨特的生境;這與蘇樑等[12]研究結果一致。根系分支率是植物對環境適應能力的體現,有研究表明根系分支率能較好地反映根系構型的可塑性變化[36];因此加強喀斯特地區細根構型的研究,以充分了解細根構型的可塑性變化,助力于石漠化的治理與恢復。
Leonardo da Vinci 法則認為,根系分支前的橫截面積等于根系分支后的橫截面積之和[32];這與徐立清等[24]對張廣才嶺不同生境胡桃楸(Juglansmandshurica)的研究,及蘇樑等[12]對喀斯特峰叢洼地不同植被恢復階段的4個優勢種根系構型的研究相同;本研究發現構樹、刺梨的細根分支前后的橫截面積之比不存在顯著差異且分別為1.04、1.03,均接近于1,符合Leonardo da Vinci 法則,表明構樹、刺梨的細根分支前后的橫截面積相等,但花椒、火棘不符合Leonardo da Vinci 法則,可能與樹種、遺傳、生境、及前人對橫截面積比的研究是基于粗根的研究等有關。
4種適生植物的細根形態多為人字形,根系分支簡單,連接長度較長,與魚尾形分支相似,更好的體現了細根形態與細根構型之間的相似性,兩者均是對生境適應的重要策略。
3.2 喀斯特石漠化生境細根和根際土壤C、N、P計量特征對細根構型的影響
對比細根養分、根際土壤養分與細根構型的相關性發現,細根養分與細根構型的相關性較小,但主要與細根N、P有關。4種適生植物中刺梨的細根養分對細根構型的影響較大,這可能與刺梨的P(0.64 mg/g)含量不高有關,且研究區石漠化程度高,植被覆蓋率低導致風化和淋溶作用較強,使得P易受損失進入水體[37-38],植物的生長受到P的限制[27]。構樹、刺梨的細根P含量分別與細根總根長、細根拓撲結構呈顯著負相關,火棘N含量與細根總根長呈顯著負相關;而花椒N∶P與細根總分支率呈顯著正相關,刺梨C∶P與細根拓撲結構呈顯著正相關。這可能是因為細根P含量、N含量較為充分且能滿足植物生長需要時,細根不在通過改變拓撲結構和延長總根長來滿足其正常生長;當N∶P、C∶P較為缺乏時,細根通過改變拓撲結構和總分支率來獲取更多的C、N來滿足其正常生長,同時也反映出P含量對計量比的影響。細根拓撲結構的大小與細根吸收水分、養分的效率有關,本研究中魚尾形分支是理想的細根構型,擁有更高的水分、養分吸收效率和適應喀斯特貧瘠生境的能力;較長的細根連接長度利于細根在土壤中伸展,增加其在土層中的分布,提高對水分和養分的吸收效率[39-40]。
根際土壤中的C含量主要來源于細根的死亡、分泌及呼吸等過程;N含量主要來源于有機質的分解、生物量的多少和植被的固氮作用,有機質的多少直接決定了根際土壤中N含量的高低;P含量主要來源于土壤的母質和成土作用。根際土壤養分與細根構型的相關性分析表明,根際土壤的C含量、P含量及C∶P與細根總分支率呈顯著負相關,根際土壤C含量、C∶N與細根橫截面積比呈顯著負相關。這可能因為根際土壤養分較為充足時,細根不在通過改變總分支率、橫截面積比來滿足其生長。根際土壤養分與細根構型之間的相關性較細根養分與細根構型之間的相關性更為顯著、關聯性更多,這可能與根際土壤是細根養分最主要、最直接的來源[27],及細根的功能是吸收根際土壤中的水分和養分有關[41]。
通過主成分分析可從多個變量中找出影響細根構型的主要因子,本研究中細根N含量及細根、根際土壤中與P相關的計量比均是影響細根構型的主要因子,特別是細根養分有更為直接的影響,這可能與細根和細根構型特征有更為直接的聯系有關。喀斯特地區植物的生長受到N、P元素的限制[16,23],養分元素的多少直接影響細根構型的發展,為更好的讓4種植物適應生境可通過對N、P元素的調節從而促進細根構型的改變,使4種植物更能適應喀斯特石漠化環境。通過養分元素的調節以改變細根構型特征是調節植物生態適應性的有效方式。然而未見有細根構型與細根養分和根際土壤養分相結合的研究,大多是單一的根系構型特征的研究。李思詩等[42]研究發現根系平均直徑是影響根系構型的主要因子;唐子欽等[43]研究發現根系總根長是影響根系構型的主要因子;單立山等[44]發現根系拓撲指數、根系連接長度、根系分支率、根系橫截面積比是影響根系構型的主要因子。本研究的細根構型特征顯示,4種植物在細根總根長、細根橫截面積比中存在顯著差異,其可能是影響細根構型的主要因子。構樹與刺梨總根長較長且存在差異可能與其本身對水分、養分的需求較高且采樣處石子較多、土層淺薄等有關,為滿足生長需要,構樹、刺梨必通過增加細根連接長度以獲得充足的水分、養分;同時也體現了喀斯特石漠化對構樹、刺梨的生長影響較大,但兩者密不可分,構樹、刺梨的生長對喀斯特石漠化的恢復與治理具有一定的促進作用。花椒、火棘的橫截面積比存在顯著差異,且不符合Leonardo da Vinci 法則,可能與樹種、遺傳、生境、及前人對橫截面積比的研究是基于粗根的研究等有關,對細根橫截面積比的研究應擴大其研究深度,豐富研究成果,但細根橫截面積比存在差異的決定性因素尚需深入研究。綜上所述,由于樹種、根系類型、樹種年限、生境等不同,影響根系構型的主要因子存在差異[45]。
3.3 喀斯特石漠化生境適生植物細根適應策略
在貧瘠的喀斯特生境中4種植物均通過降低細根分支率、增加細根的連接長度以獲得充足的水分和養分,細根吸收效率與細根分支、連接長度具有一定相關性[31],較少的細根分支和更長的細根連接長度可能會增加細根的比重,促使細根的吸收能力增加;且細根比重的增加可提高對土壤的固結能力,有效的防止水土流失;同時魚尾形分支具有較強的空間拓展能力和避免根系內部競爭從而快速占領冗余空間,呈現出一種特殊的生長模式—根包石。喀斯特地區養分缺乏[16,27],但土壤養分是細根養分的主要來源之一[46]。本研究中根際土壤養分、細根養分均較低,加之獨特的喀斯特生境,植物為適應生境,通過魚尾形分支模式、較長的細根連接長度等在貧瘠的生境中獲得充足的養分,達到一種哪有較充足的養分就往哪兒伸展的生長機制。4種植物對喀斯特生境的適應性可能與細根N含量影響細根壽命及周轉有關,細根N含量增加、細根周轉變快、非結構性碳水化合物降低等將會導致細根壽命減短,同時細根N含量也會影響細根周轉速率的變化,從而對細根的適應性產生一定的影響[47];當生境發生變化時,4種植物可能通過改變細根N含量以影響細根壽命和周轉,同時轉變細根構型特征及對養分的吸收利用效率等方式以適應生境的變化。細根構型特征、細根養分、根際土壤養分綜合用于根系的研究能精準的反映細根的生態適應策略,可助力于喀斯特生態系統保護與恢復。
4 結論
(1)喀斯特石漠化適生植物構樹、花椒、刺梨、火棘細根均呈魚尾形分支模式、較長的細根連接長度、較小的細根分支率,其生態適應性大致相同,獨特的生境中“根包石”現象突出,植物具有較強的貧瘠生境適應能力。
(2)研究區的構樹、刺梨細根分支前后的橫截面積比符合Leonardo da Vinci 法則,花椒、火棘細根分支前后的橫截面積比存在差異,不符合Leonardo da Vinci 法則,且細根分支前后的橫截面積比是否符合Leonardo da Vinci 法則可能因樹種而異。
(3)相關性分析表明,細根N、P含量及與P相關的計量比和根際土壤C、N、P含量及與C相關的計量比分別與細根構型特征的相關性顯著;主成分分析表明,細根養分、根際土壤養分對細根構型特征有一定的影響,其中細根N含量及細根、根際土壤中與P相關的計量比對細根構型特征的影響較大。
(4)研究區養分含量低且對植物的生長有雙重影響,魚尾形分支降低了根系內部的水分、養分競爭,植被不斷的調整細根構型特征以適應喀斯特貧瘠的生境。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
