氮添加對黃河三角洲濱海濕地蘆葦養分再吸收效率的影響
2022-03-07 13:16:06陶寶先王晶東陳慶海袁海燕陳永金
生態學報 2022年3期
陶寶先,王晶東,陳慶海,馬 澍,袁海燕,陳永金,王 浩
聊城大學地理與環境學院,聊城 252059
植物可以采取各種不同的策略適應外界環境變化[1]。其中,再吸收作為一種養分保護機制,是指植物將養分從脫落的葉片轉移至根部的過程[2—4]。植物可以直接利用再吸收的養分,減少對外界氮、磷養分供應的需求[4—5]。通常,土壤養分可利用性增強會降低植物的養分再吸收效率[6—8],如外源氮添加降低植物對氮素的再吸收效率[9]。在溫帶濕地中植物生長常受氮素限制[10],外源氮添加可能緩解氮素對植物生長的限制,降低其對氮素的再吸收效率。此外,磷的再吸收效率對外源氮添加的響應目前仍不確定。前期研究發現,氮添加能顯著改變磷的再吸收效率[8],但其他研究也發現氮添加不會改變磷的再吸收效率[11]。因此,氮添加導致的土壤養分可利用性變化,可能改變溫帶濕地植物對養分的再吸收過程。
現有研究多關注植物葉片的養分再吸收過程,然而莖[12—13]、鞘[14]等器官中的養分也能被植物再吸收。由于莖對地上生物量中具有顯著貢獻[15],其在植物養分再吸收過程中占有不可忽略的地位[5, 13],且其養分再吸收過程也受外源養分添加的影響。早期研究發現,長期氮添加增加濕地凋落物氮含量,降低葉、莖等的養分再吸收效率[14]。然而,莖、鞘等器官的養分再吸收過程常被忽略[5, 14, 16]。此外,植物生長對不同形態氮素可能具有偏好性吸收。前期研究發現,水生植物吸收銨態氮較硝態氮消耗較少的能量[17—18],故水生植物偏好于吸收銨態氮[19]。甚至有研究發現,濱海濕地蘆葦能直接利用溶解性有機氮[20]。這暗示:植物對不同形態氮素的偏好性利用極可能影響其對養分的再吸收過程。各種形態氮素是否對植物的養分再吸收效率有不同的影響?目前的研究仍不充分。因此,研究植物葉、莖的養分再吸收效率及其對不同形態氮添加的響應,可深入理解土壤養分可利用性與植物內部養分保護之間的關系。
黃河三角洲是世界上較年輕的、典型的河口濕地之一[21]。蘆葦是黃河三角洲重要的植物物種,但由于氣候變化及人類活動的影響,過去幾十年蘆葦在該區的分布面積持續下降,影響了該區的植被恢復進程[22]。大氣氮沉降作為黃河三角洲濱海濕地的主要氮素來源之一,能夠調節濱海濕地諸多生態過程,然而氮沉降對黃河三角洲濱海濕地生態過程的影響研究仍不充分[23],尤其是氮沉降對黃河三角洲蘆葦的養分再吸收過程的影響研究仍不清楚。因此,本研究通過兩年的原位模擬氮添加實驗,研究氮添加(硝態氮、銨態氮)對黃河三角洲濱海濕地蘆葦養分再吸收效率(氮、磷)的影響,以期明確以下科學問題:(1)氮沉降是否影響黃河三角洲濱海濕地蘆葦葉、莖的養分再吸收效率?(2)蘆葦的養分再吸收效率是否對硝態氮、銨態氮添加具有不同的響應?(3)蘆葦葉與莖是否具有相同的養分再吸收效率?
1 材料與方法
1.1 研究區概況
研究區位于山東省東營市中國科學院黃河三角洲濱海濕地生態試驗站(37°45′50″N, 118°59′24″E)。該區屬于暖溫帶大陸性季節氣候,年均溫11.7—12.8℃,無霜期196 d,年蒸發量190—2400 mm,年降水量552—630 mm,且近70%的降水集中在7、8、9月份,雨熱同期,四季分明。在7、8月份大雨后研究區地表常有暫時性積水。該區土壤類型以鹽土和潮土為主[24],平均土壤鹽度約為0.69%[21]。植被以耐鹽物種為主,主要有檉柳(Tamarischinensis)、蘆葦及鹽地堿蓬(Suaedasalsa)[25]。在本研究的試驗點,主要植被類型為蘆葦。
1.2 樣地布置
2016年2月,在研究區設置對照、硝態氮(NaNO3)、銨態氮(NH4Cl)3個處理,每個處理設置3個樣方,共9個樣方。每個樣方(長2 m ×寬2 m)用4塊不銹鋼板焊接成的不銹鋼筒(長2 m ×寬2 m ×高0.8 m)與外界隔離,為防止樣方內添加氮素的流失,將不銹鋼筒插入地下約0.4 m深,地表以上約0.4 m高。相鄰樣方間留2 m的緩沖區。早期研究發現,研究區年平均氮沉降量為2.8 g/m2[26],且未來有增加趨勢[27]。因此,本研究設置的年氮沉降量為8 g/m2,每年分4次輸入(3、6、9、12月各一次),每次的輸入量為2 g/m2。氮素用2 L樣地附近溝塘的地表水溶解,用噴壺均勻地噴灑于樣地地表,對照處理僅噴灑等量的地表水。
1.3 樣品采集與分析
2017年5月,在每個樣方內選擇長勢均勻的10株蘆葦并做標記。2017年6—9月,每月中旬于每個樣方內齊地剪除標記的蘆葦2株,剪下完全展開的葉片及其對應部位的莖,用于分析全氮、全磷含量。10月中旬起每周觀察各樣方內植物的生長狀況,至各樣方內標記蘆葦完全變黃時采集地上部分。每月采樣后,將每個樣方2株蘆葦的葉或莖樣品分別混合成一個混合樣,再用去離子清洗干凈,于70℃烘干至恒重,粉碎、過0.1 mm篩。樣品采用濃硫酸-H2O2消煮,全氮采用靛酚藍比色法測定,全磷采用鉬銻抗比色法測定,有機碳采用高溫外加熱重鉻酸鉀氧化法測定[28]。
1.4 數據處理與統計分析
養分再吸收效率=(Cm-Cs)/Cm。其中,Cm代表成熟器官的養分含量,Cs代表脫落器官的養分含量。6—9月樣品中全氮或全磷含量最高值代表成熟器官的養分含量,葉、莖成熟器官氮含量及葉成熟器官磷含量均采用6月份數據,莖成熟器官磷含量采用8月份數據。10月份蘆葦完全變黃時各器官的全氮或全磷含量代表脫落器官的養分含量。
采用單因素方差分析(Tukey法)檢驗各指標在不同處理間的差異,顯著性水平為P<0.05。采用SPSS 25.0與Excel 2016進行數據統計和計算,采用Origin 2016作圖,圖中誤差線為標準差。
2 結果
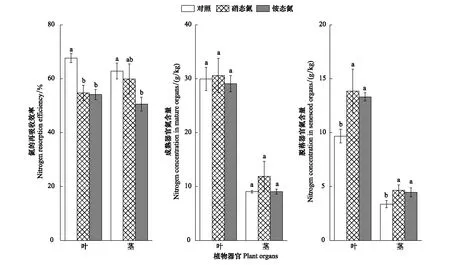
圖1 氮輸入對氮的養分再吸收效率及其成熟、脫落器官氮含量的影響Fig.1 Effects of nitrogen additions on nitrogen resorption efficiency and concentrations of nitrogen in mature and senesced organs 小寫字母代表不同處理間有顯著差異(P<0.05)
硝態氮、銨態氮添加均顯著增加脫落器官(葉與莖)的氮、磷含量(P<0.001),其中,脫落葉片氮含量增加43.22%(硝態氮)、37.60%(銨態氮),脫落莖氮含量增加38.28%(硝態氮)、32.74%(銨態氮),脫落葉片磷含量增加30.91%(硝態氮)、36.36%(銨態氮),脫落莖磷含量增加33.33%(硝態氮)、36.51%(銨態氮);但同一脫落器官的氮或磷含量在兩類氮輸入處理間無顯著差異。兩類氮添加對成熟器官氮、磷含量無顯著影響(圖1、2)。成熟葉片N/P比值為16.33(對照)、18.25(硝態氮)、16.46(銨態氮),莖N/P比值為6.39(對照)、9.53(硝態氮)、6.92(銨態氮;圖3)。兩類氮添加顯著降低蘆葦凋落物(即脫落器官)的C/N、C/P(P<0.01),葉凋落物的C/N均值為48.02(對照)、33.91(硝態氮)、34.28(銨態氮),莖凋落物的C/N均值為138.74(對照)、101.55(硝態氮)、104.75(銨態氮),葉凋落物的C/P均值為2175.13(對照)、1681.52(硝態氮)、1611.43(銨態氮),莖凋落物的C/N均值為859.39(對照)、655.68(硝態氮)、624.12(銨態氮;圖4)。
葉片氮的再吸收效率為51.83%—69.51%,均值為67.66%(對照)、54.76%(硝態氮)、54.14%(銨態氮);莖氮的再吸收效率為47.64%—64.76%,均值為62.85%(對照)、59.95%(硝態氮)、50.60%(銨態氮)。葉片磷的再吸收效率為51.30%—75.68%,均值為70.38%(對照)、56.80%(硝態氮)、57.63%(銨態氮);莖磷的再吸收效率為75.56%—85.91%,均值為84.95%(對照)、77.43%(硝態氮)、78.33%(銨態氮)。兩類氮添加顯著降低葉、莖的養分再吸收效率(P<0.001),其中,葉片氮的再吸收效率降幅為19.06%(硝態氮)、19.98%(銨態氮),莖氮的再吸收效率降幅為4.61%(硝態氮)、19.48%(銨態氮),葉片磷的再吸收效率降幅為19.30%(硝態氮)、18.12%(銨態氮),莖氮的再吸收效率降幅為8.85%(硝態氮)、7.79%(銨態氮)。針對同一器官,氮或磷的再吸收效率在兩類氮輸入處理間均無差異。相同氮輸入處理或對照處理下,葉、莖的氮素再吸收效率無顯著差異,但莖磷的再吸收效率顯著高于葉片(P<0.01;圖1、2)。

圖2 氮輸入對磷的養分再吸收效率及其成熟、脫落器官磷含量的影響Fig.2 Effects of nitrogen additions on phosphorus resorption efficiency and concentrations of phosphorus in mature and senesced organs 小寫字母代表不同處理間有顯著差異(P<0.05)

圖3 氮輸入對成熟葉、莖N/P比的影響Fig.3 Effects of nitrogen additions on the ratio of N/P in mature leaf and culm 小寫字母代表不同處理間有顯著差異(P<0.05)
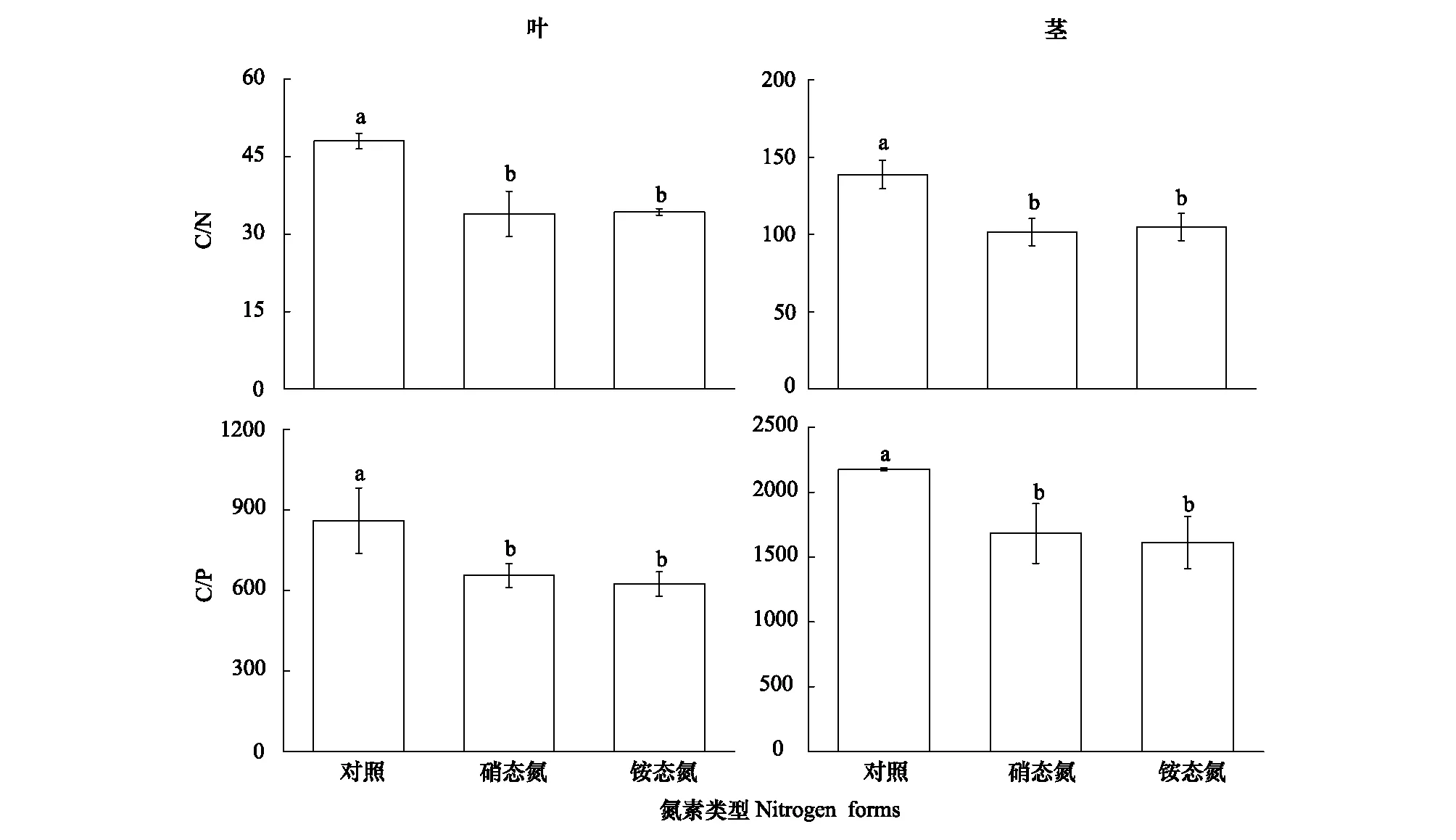
圖4 氮輸入對葉、莖凋落物C/N、C/P比的影響Fig.4 Effects of nitrogen additions on the ratios of C/N and C/P in senesced leaf and culm小寫字母代表不同處理間有顯著差異(P<0.05)
3 討論
植物葉片養分含量增加通常會降低養分的再吸收效率[7]。然而,早期研究也發現,氮添加僅能降低溫帶草原部分物種葉片氮的再吸收效率[29]。本研究發現,硝態氮、銨態氮添加均增加了脫落器官的氮含量,但降低了氮的再吸收效率(圖1),這與其他草原[6, 8]及濕地[14]生態系統的研究結論相似,即外源氮添加增加土壤氮的可利用性,促進植物對氮的吸收,降低氮的再吸收效率[6, 8]。溫帶濕地植物生長通常受氮素限制[10],這意味著外源氮添加可能促進植物吸收氮素,增加脫落器官氮含量,進而降低氮的再吸收效率。近期研究也發現,外源氮添加能增加土壤硝態氮、銨態氮含量,促進黃河三角洲蘆葦的生長[21]。生態化學計量比是反映植物生長狀況的重要指標,早期研究曾將N/P<14、N/P>16、14
早期研究發現,水生植物對不同形態氮素有選擇性吸收[17—18, 20]。這暗示:硝態氮、銨態氮可能對植物的養分再吸收有不同的影響。然而,本研究發現,兩類無機氮添加對養分再吸收效率產生相似的作用(圖1、2),這與Kou等[11]的研究結論相似。其原因可能是:(1)黃河三角洲濱海濕地土壤氮素含量明顯低于其他濱海濕地,如長江口[31]。這可能加劇氮素對植物生長的限制,增加蘆葦生長對氮素的需求,蘆葦有可能吸收各種形態的氮素來滿足自身生長對氮素的需求。(2)早期研究發現,濱海濕地蘆葦能夠吸收無機及有機形態的氮素[20],這表明蘆葦對不同形態氮素可能無差別吸收。這可能導致兩種形態氮素雖能降低養分的再吸收效率,但兩者的效果無差別。
目前,磷的再吸收效率與土壤氮素可利用性之間的關系仍不確定。前期研究發現,外源氮添加能顯著增加[8, 32]、降低[6]植物葉片磷的再吸收效率,或對磷的再吸收效率無顯著影響[33]。外源氮添加促進植物生長,這極有可能產生植物生長的稀釋作用,使植物生長轉為磷限制[34]。本研究的N/P比也表明(圖3),蘆葦葉片生長受磷限制。這意味著:氮添加可能增加植物生長對磷的需求,促進植物對磷的再吸收。然而,本研究卻發現,外源氮添加顯著降低磷的再吸收效率,這與Lü 等[6]的研究結論相似:外源氮添加可能促進土壤磷的周轉,增加土壤磷的可利用性,進而降低植物生長對磷的需求及其再吸收效率。近期研究發現,硝態氮、銨態氮添加刺激了黃河三角洲濱海蘆葦濕地土壤磷酸酶活性[21],其他淡水濕地的研究也發現相似結論[35],這表明外源氮添加可能通過刺激磷酸酶活性增加土壤磷的可利用性,這極有可能緩解植物生長對磷的需求,進而降低植物對磷的再吸收效率。
前期研究多關注葉片的養分再吸收效率,而莖、鞘等器官的養分再吸收過程常被忽略。早期研究已發現,葉片的養分再吸收效率與莖相似[12],表明除了葉片之外的其他器官也對植物的養分再吸收有著不可忽略的貢獻。本研究發現,蘆葦的葉(58.86%)、莖(57.80%)具有相似的氮的再吸收效率,且與全球平均水平相近(62.1%)[7]。蘆葦葉片磷的再吸收效率均值為61.60%,與全球陸地生態系統植物對磷的再吸收效率均值相近(64.9%),但蘆葦莖磷的再吸收效率(均值80.24%)卻顯著高于全球平均水平[7],這與Aerts等[36]的結果(91%)相近。雖然蘆葦莖的氮、磷含量低于葉(圖2),但莖的生物量是葉的2.9倍[37],且莖磷的再吸收效率也要高于葉,這意味著蘆葦莖養分的再吸收量要明顯高于葉,即黃河三角洲濱海濕地蘆葦的莖較葉片對養分的再吸收具有更大的貢獻,尤其是磷。同時,本研究也建議,后續研究應該注重對非葉器官(如莖、鞘等)養分再吸收過程的研究。
本研究發現,外源氮添加增加了脫落器官氮、磷含量,降低了凋落物C/N、C/P(圖4),改變凋落物質量。也有研究發現,長期氮添加減少濕地挺水植物葉片易分解碳含量,促進莖中難分解碳組分的累積[38],降低了葉、莖凋落物質量。而凋落物生態化學計量比(如C/N、C/P、N/P等)及有機碳質量[39—40]的變化能進一步影響凋落物分解過程[41—42],甚至改變濕地碳循環過程。因此,后續應進一步加強植物養分再吸收效率變化對凋落物分解的研究。
4 結論
(1)氮添加增加了蘆葦脫落葉、莖的氮、磷含量,降低了蘆葦葉、莖的氮素與磷的再吸效率。同一脫落器官的養分(氮或磷)含量及同一器官的養分(氮或磷)再吸收效率在兩類氮添加處理間均無顯著差異。
(2)蘆葦葉、莖具有相似的氮素再吸收效率,但莖磷的再吸收效率顯著高于葉,表明:蘆葦莖對養分再吸收過程具有不可忽略的貢獻。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
甘肅教育(2020年14期)2020-09-11 07:57:42
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
時代英語·高二(2015年1期)2015-03-16 00:08:11
