木聚糖酶TgXyn2的酶學特性研究
2022-03-06 07:07:10葛雨杭陳美汐吳松娜
食品工業科技 2022年4期
葛雨杭,陳美汐,孫 博,吳松娜,萬 群,
(1.南京農業大學食品科技學院,江蘇南京 210095);2.南京農業大學理學院,江蘇南京 210095)
半纖維素是地球上第二豐富的可再生生物資源。木聚糖是半纖維素的主要骨架,由β-(1,4)糖苷鍵連接的木糖亞基組成,并在主鏈木糖2號和3號位存在不同的取代基[1]。半纖維素對維持植物細胞壁的完整性和細胞纖維的凝聚力起到重要的作用,廣泛分布于植物的細胞壁中,約占植物細胞干重的15%~35%[2]。木聚糖作為分子量大的高聚物,需要先降解成小分子的低聚木糖,才可以被有效地利用[3]。
木聚糖酶在自然界分布廣泛,可從動物、植物和微生物中獲得。木聚糖水解酶系(Xylanolytic enzyme systems)是一類降解木聚糖的酶系,包括內切-1,4-β-木聚糖酶、β-D-木糖苷酶、α-L-阿拉伯糖苷酶、α-D-葡糖苷酸酶、乙酰基木聚糖酶和酚酸酯酶[4],多應用在烘焙、釀造、飼料等食品工業中[5]。在烘焙食品生產中,木聚糖酶可以通過改變面筋網絡結構增大面團體積,使面團的彈性和延展性增加,干燥度、硬度及持水性降低,咀嚼性和黏性減少,從而改善面團的加工及穩定性能[6];在葡萄酒釀造工業中,木聚糖酶有利于葡萄皮浸漬和顏色的提取,易于澄清和過濾,改善葡萄酒質量和穩定性[7];在飼料行業中,木聚糖酶能夠降低谷物在動物腸道中的食糜黏度,提高飼料消化率[8]。
本文研究一種從貴州木霉菌株(Trichoderma guizhouense)中發現的內切-1,4-β-木聚糖酶TgXyn2(OPB43840.1,Endo-1,4-beta-xylanase 2),屬于第11家族的木聚糖酶[9],可以有效地降解木聚糖主鏈的β-1,4木糖苷鍵。本文通過大腸桿菌異源表達獲得TgXyn2,以木聚糖為底物,在不同的溫度、pH、底物來源、金屬離子及變性劑條件下研究其酶學特征,以期發掘一種新的木聚糖降解酶,為其在食品等輕工業生產中的實際應用提供理論依據。
1 材料與方法
1.1 材料與儀器
大腸桿菌E.coliBL21-Gold(DE3)菌株、表達質粒pCold-TF 實驗室保存;LB液體培養基:蛋白胨10 g/L、酵母提取物5 g/L、氯化鈉10 g/L;LB固體培養基:蛋白胨10 g/L,酵母提取物5 g/L,氯化鈉10 g/L,1.5%瓊脂;基本培養基:十二水合磷酸氫二鈉13.24 g/L、無水硫酸鎂0.12 g/L、磷酸二氫鉀1.6 g/L、硫酸銨7 g/L、檸檬酸氫二胺0.5 g/L、4%甘油。
G180TW高壓滅菌鍋 美國致微ZWALWAY公司;IS-RDS3疊加式恒溫搖床 美國CRYSTAL公司;ELITIST 22K-R立式高速冷凍離心機 湖南吉爾森科技發展股份有限公司;SW-CJ-IFD超凈工作臺 江蘇凈化設備有限公司;UH-03低溫高壓細胞破碎儀 永聯生物科技有限公司;PTT-A1000電子天平 福州華志科學儀器有限公司;B-500超微量核酸蛋白檢測儀 上海元析儀器有限公司;MDF-382E超低溫保存箱 松下冷鏈大連有限公司;VELOCITY 18R高速冷凍離心機 南京基天生物技術有限公司;JY300C電泳儀 君意電泳;NAS8000蛋白純化系統 蘇州利穗。
1.2 實驗方法
1.2.1 構建質粒與轉化菌種 木聚糖酶TgXyn2的氨基酸序列由南京農業大學理學院萬群課題組提供,來自貴州。TgXyn2氨基酸序列總長度為185,由通用生物系統(安徽)有限公司合成并克隆至pCold-TF表達載體上。上游引物:ACGCCATATCGCCGA AAGG;下游引物:GGCAGGGATCTTAGATTCTG;過程:以第一鏈cDNA為模板,用上游引物和下游引物進行PCR擴增。擴增程序如下:95 ℃,3 min;95 ℃,30 s,60 ℃,30 s,72 ℃,1 min,30個循環;72 ℃延伸7 min,即可獲得PCR擴增產物。利用堿裂解法大量提取和純化表達質粒,再用熱休克的方法,將質粒轉入表達菌株,具體過程如下:取50 μL表達感受態Rosetta冰上解凍后,取2 μL重組質粒緩慢加入并冰置25 min,42 ℃水浴熱激45 s,冰浴2 min;加入500 μL無抗LB溶液,37 ℃ 200 r/min下培養1 h。取適量菌液涂布到含有1 μL/mL氨芐青霉素的LB固體培養皿中,倒置在37 ℃保溫箱里培養12 h。
1.2.2 酶的表達與純化 挑取固體培養皿上生長出來的單菌落接種到含50 mg/L氨芐青霉素的LB液體培養基中,在37 ℃,200 r/min培養至OD600為0.6,降溫到15 ℃,加入誘導劑0.5 mmol/L IPTG,繼續培養24 h。通過離心收集細胞,用裂解液溶解沉淀細胞,低溫高壓細胞破碎儀破碎后,離心取上清,通過Ni親和層析柱純化TgXyn2,獲得粗酶液然后加入1%(質量比)的TEV;混合物透析過夜后,再次純化透析,在280 nm的吸光度下測量確定蛋白質濃度,分裝保存。
采用SDS-PAGE電泳分析方法[10],配制10%分離膠與4 %濃縮膠,將蛋白按照1:1的比例加入到含有5%的β-巰基乙醇樣品緩沖溶液中,95 ℃加熱4 min,12000×g離心1 min上清液電泳,上樣跑膠,煮沸水洗染色后進行膠圖分析,鑒定蛋白表達情況。
1.2.3 TgXyn2酶學特性
1.2.3.1 酶活測定 木聚糖酶能夠將木聚糖降解為木糖等還原性糖,因此可以通過比色法DNS法[11]測定產生的還原糖,進而確定木聚糖酶的酶活[12]。本文以0.2%(m/v)木聚糖為底物,每組處理3個重復,測定時每個試管加入100 μL的0.5 mg/mL TgXyn2酶液進行反應,反應時間10 min。酶活力(U)單位定義:在一定條件下,1 mg木聚糖在1 min內催化底物產生1 μmol還原糖為1個酶活力單位(U)。通過使用Origin 9.0(OriginLab,USA)Hill函數,當n=1時可以模擬米氏方程,擬合得到Vmax和Km值。
1.2.3.2 溫度對酶活性的影響 酶反應的溫度梯度設定為25、30、35、40、45、50和55 ℃,各組pH保持在5.0。
1.2.3.3 pH對酶活性的影響 各組反應體系溫度為35 ℃,考察pH2.0、3.0、4.0、5.0、6.0、7.0、8.0的影響,pH條件2.0和3.0使用的是50 mmol/L甘氨酸-鹽酸緩沖液,pH條件4.0、5.0、6.0使用的是50 mmol/L乙酸-乙酸鈉緩沖液,pH7.0、8.0條件使用的是50 mmol/L檸檬酸-磷酸氫二鈉緩沖液。
1.2.3.4 酶動力學參數 在最適條件下,底物梯度濃度分別設置0.1~5.0 mg mL-1(從第二組開始,每0.2 mg mL-1設置一組),并以此計算酶動力學參數。
對測得的反應速度與相應底物濃度進行回歸分析,求得Km值。計算公式為米氏方程(Michaelis-Menten equation):v=Vmax×[S]/(Km+[S]),v代表反應初速度,Vmax代表最大反應速度,[S]代表底物濃度。此運算可同時得到Vmax。同理,根據公式Vmax=Kcat×[E],計算Kcat。
1.2.3.5 不同化學試劑對酶活性的影響 分別在酶反應體系中加入0.1 mmol/L的鹽酸胍、EDTA、DTT、SDS等化學試劑,在最適條件下與0.2%(m/v)的木聚糖底物進行反應。以未加化學試劑的情況下測得的TgXyn2活性為100%,比較不同化學試劑對酶活性的影響。
1.2.3.6 金屬離子對酶活性的影響 分別在酶反應體系中加入0.1 mmol/L的Na+、K+、Ca2+、Fe3+、Cu2+、Mg2+、Zn2+、Co2+、Cd2+、Mn2+、Li+等金屬離子,在最適條件下與底物進行反應。以未加金屬離子的情況下測得的TgXyn2活性為100%,比較不同金屬離子對酶活性的影響。
1.2.3.7 酶的溫度耐受性 將TgXyn2在20~55 ℃(每隔5 ℃設置一組)條件下分別保溫1 h后,在最適條件下與底物進行反應。以未保溫處理的TgXyn2活性為100%,比較經過不同溫度處理后TgXyn2的酶活。
1.2.3.8 酶的酸堿耐受性 選用pH范圍為2.0~8.0的緩沖液稀釋酶液,并將處理后的酶液于4 ℃條件下存放1 h后,在最適條件下與底物進行反應。以未經緩沖液處理的TgXyn2活性為100%,比較經過不同酸堿度處理后TgXyn2的酶活。
1.2.3.9 酶分解不同底物的能力 分別以(甘蔗渣來源)木聚糖、(小麥麩皮來源)木聚糖、(燕麥漿來源)木聚糖、微晶纖維素(MCC)作為酶反應體系中的底物,底物濃度為0.2%,在最適條件下與木聚糖酶TgXyn2進行反應。以(甘蔗渣來源)木聚糖為底物時的TgXyn2活性為100%,測定反應后體系的旋光度,比較TgXyn2對不同底物的分解能力。
1.2.4 同源結構建模 將TgXyn2的氨基酸序列上傳至I-TASSER在線網站(https://zhanglab.ccmb.med.umich.edu/I-TASSER/)進行同源三維結構建模[13],利用PyMOL軟件畫出蛋白三維結構圖[14]。
1.3 數據處理
采用GraphPad Prism7處理軟件進行數據處理和統計分析,根據實驗數據繪制折線圖和柱狀圖,總結現象趨勢,分析實驗結論以評價模型的統計意義。
2 結果與分析
2.1 內切-1,4-β-木聚糖酶的表達、純化與電泳鑒定
利用ExPASy在線網站預測并結合已知基因序列,通過氨基酸序列計算得出TgXyn2的分子量大約是20.19 kDa。根據上述的實驗操作,用堿裂解法大量提取和純化表達質粒,用熱休克法將質粒轉入E.coliBL21-Gold(DE3)表達宿主細胞,對TgXyn2成功誘導表達,SDS-PAGE膠圖結果如圖1所示。
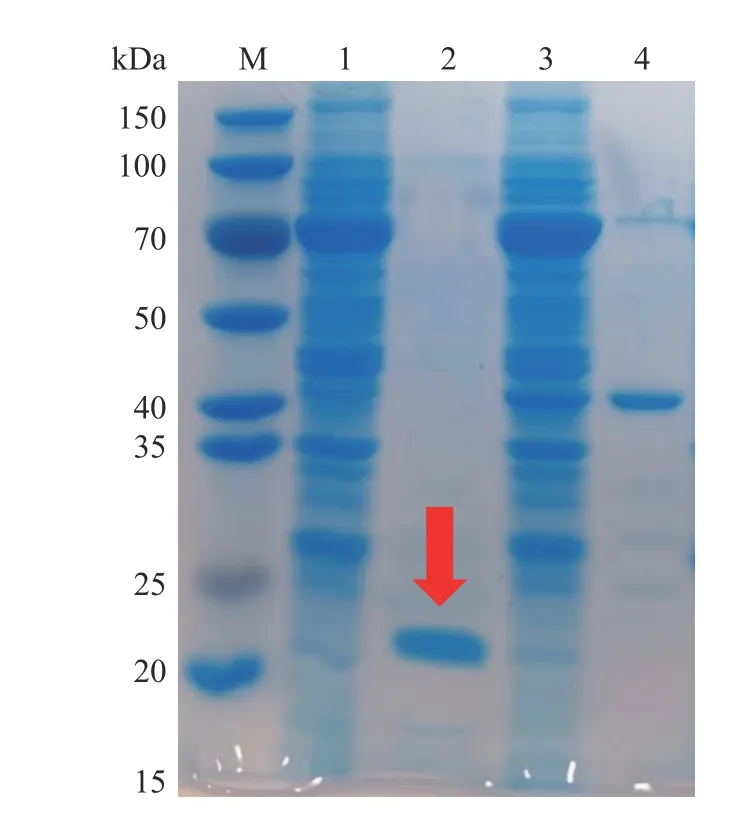
圖1 TgXyn2的SDS-PAGE分析Fig.1 SDS-PAGE analysis of TgXyn2
在標準蛋白M條帶的相對分子質量20 kDa的位置附近出現清晰可見的一條較濃的條帶,與預測的木聚糖酶TgXyn2分子量20.19 kDa十分接近,說明TgXyn2得到了有效表達和純化。上樣剩余條帶1與上樣流出條帶3,均在70 kDa附近存在濃度較高的蛋白,經推斷可知為含有融合蛋白的目的蛋白;雜蛋白條帶4主要集中在40~50 kDa范圍內,經推斷可知此為酶切后的融合蛋白。
2.2 內切-1,4-β-木聚糖酶酶學性質分析
2.2.1 溫度對酶活性的影響 TgXyn2在35 ℃時酶活性達到最大,而在低溫下(25 ℃左右)依舊能保持60%以上的活性,溫度達到50 ℃以上時酶活性明顯降低,溫度繼續升高后酶活性降低至20%以下(圖2A),原因可能是在高溫條件下蛋白質結構發生改變,不能形成與底物結合的空間結構,從而失去降解底物的能力。
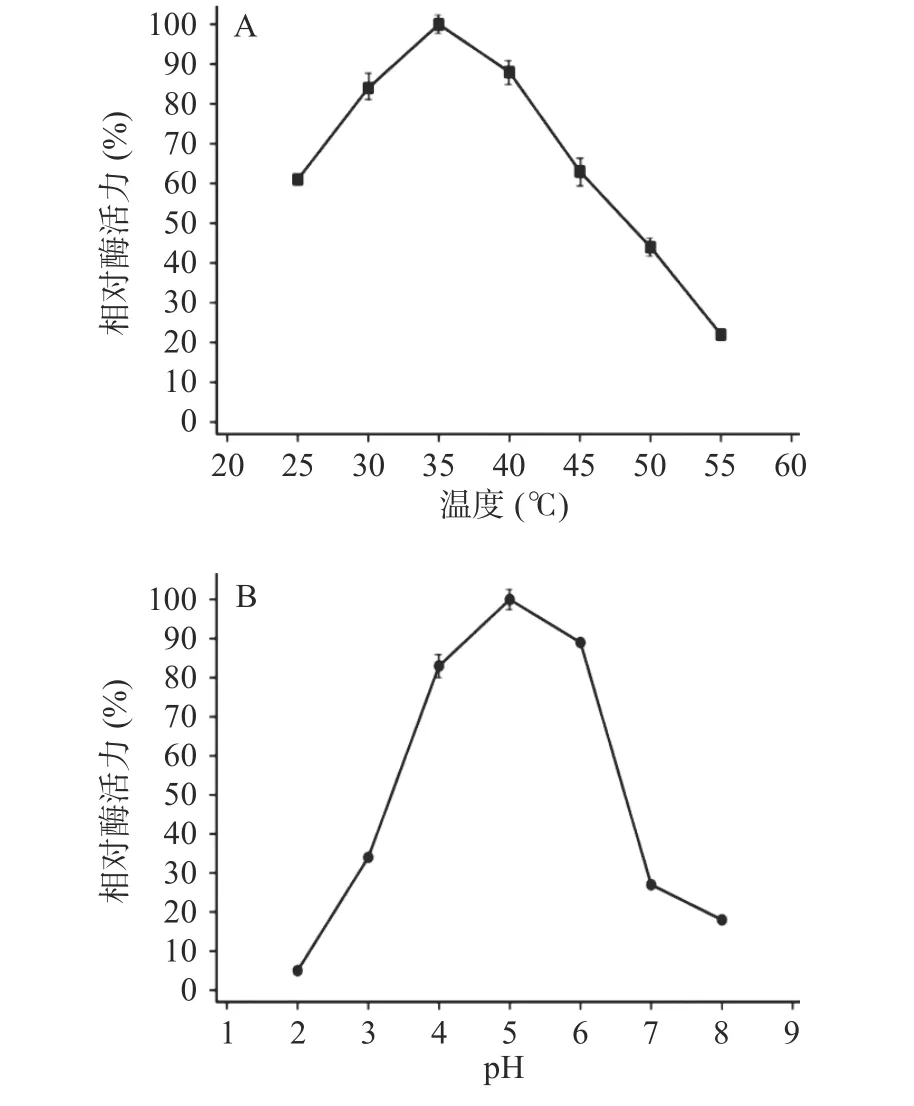
圖2 TgXyn2活性在不同溫度和pH條件下的變化Fig.2 TgXyn2 activity under different temperature and pH
2.2.2 pH對酶活性的影響 TgXyn2活性在pH5.0時達到最大值,在pH6.0~7.0過程中活性明顯下降,pH8.0時活性低于20%;而在pH3.0時仍保留了35%左右的酶活,可以推斷出TgXyn2更適應偏酸性環境(圖2B)。因此,TgXyn2酶在最適pH范圍4.0~6.0之間表現出較高活性,大于或小于最適pH,都會降低酶活性。主要是因為過酸、過堿影響酶的穩定性,進而使酶遭受不可逆破壞。此外也存在改變底物分子和酶分子的帶電狀態和天然構象,從而影響酶和底物的結合的情況。
2.2.3 酶動力學參數的計算 結合圖2A、圖2B實驗數據,通過使用Origin 9.0(OriginLab,USA)Hill函數,當n=1時可以模擬米氏方程,擬合計算出TgXyn2酶催化(甘蔗渣來源)木聚糖底物的Km、Vmax值。得到在pH5.0、溫度35 ℃的最適條件下TgXyn2的比活力為84.47 U/mg,Km=1.287 mg/mL,Vmax=2.083 μmol·min-1·mg-1;由Vmax=Kcat×[E0]得Kcat=57.47 s-1。
當酶有多個可利用的底物時,其對不同底物的催化效率可能差別很大。可以用Kcat/Km來確定酶的最適底物,當利用蛋白質工程對酶進行改造時,不同突變型的Kcat/Km也是需要要測定的參數,用來表征酶催化效率的變化情況。
2.2.4 不同化學試劑對酶活性的影響 據方差分析,1 mmol/L的DTT對TgXyn2的酶活性具有明顯提高作用,能夠達到未經處理的TgXyn2活性的137.31%;1 mmol/L的鹽酸胍對酶活性的影響效果不明顯,只達到103.69%;而1 mmol/L的EDTA、SDS對TgXyn2具有明顯抑制作用,加入酶反應體系后,酶活分別下降到未經處理的TgXyn2活性的57.11%和45.12%(圖3)。綜上所述,DTT對TgXyn2的活性具有促進作用,EDTA和SDS對酶活均有較大抑制作用。

圖3 不同化學試劑對 TgXyn2 酶活性的影響Fig.3 Effects of different chemical reagents on TgXyn2 enzyme activity
2.2.5 金屬離子對酶活性的影響 1 mmol/L的Co2+可以明顯提高TgXyn2的酶活性,能夠達到未經處理的TgXyn2活性的127.13%;而1 mmol/L的Cu2+、Cd2+、Mn2+和Fe3+的環境下TgXyn2活性下降明顯,分別下降到未經處理的TgXyn2活性的67.22%、73.64%、79.73%和89.70%(圖4)。綜上所述,Co2+對TgXyn2活性的促進最明顯,Cu2+等對酶活有較大抑制作用。馬騰飛等[15]研究表明,在畢赤酵母中表達的木聚糖酶XynB,同樣是Co2+對木聚糖酶活性有較明顯的促進作用, Cu2+對其有抑制作用,不同的是Fe2+對該酶的活性有微弱的促進作用。由于金屬離子可以直接與酶的過渡態功能基團配位,穩定過渡態的幾何結構和電荷,或者通過長程的靜電作用穩定過渡態電荷,也可以通過引起pKa的改變,從而活化堿基或糖基,因此金屬離子對于酶的活性具有一定的促進或抑制作用。

圖4 金屬離子對TgXyn2酶活性的影響Fig.4 Effects of metal ions on TgXyn2 enzyme activity
2.2.6 酶的溫度耐受性 木聚糖酶TgXyn2在20~35 ℃溫度范圍內處理后的殘余酶活較為穩定,能保持在原酶活性的55%以上。超過40 ℃時,處理后的殘余酶活顯著 (P<0.05)降低至5%以下,說明溫度對木聚糖酶TgXyn2有明顯的影響(圖5),超過40 ℃時酶活較低的原因,主要是酶的構象被破壞,底物不能和活性中心有效結合。該酶的溫度耐受性不高,熱穩定性具有很大的改造空間[16]。相比于從高寒草甸土壤樣品中篩選出低溫木聚糖酶FY8[17],其最適反應溫度為40 ℃,20~70 ℃時酶活力均隨溫度升高先增后降,在40 ℃時酶活力為最大值,與本文的TgXyn2最適溫度接近,所以也屬于低溫酶,但其溫度耐受范圍更廣,熱穩定性更好,耐酸性則稍差。
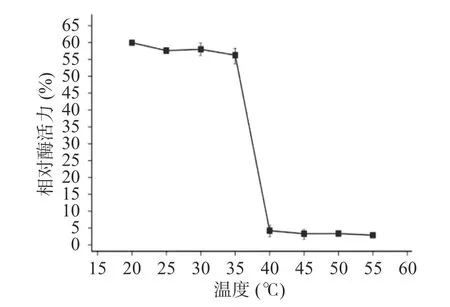
圖5 TgXyn2的溫度耐受性Fig.5 Temperature tolerance of TgXyn2
2.2.7 酶的酸堿耐受性 木聚糖酶TgXyn2在pH3.0~7.0范圍內處理后的殘余酶活較為穩定,能保持在45%以上,在pH4左右酶活殘留最高,超過55%;在pH低于2.0時殘余酶活略有下降,在30%左右;而pH高于7.0時,處理后的殘余酶活降低至5%左右,由此可見該酶更適應酸性環境[18](圖6)。

圖6 TgXyn2的酸堿耐受性Fig.6 Acid base tolerance of TgXyn2
2.2.8 酶降解不同底物的能力 在測定的4種底物中,TgXyn2只能降解木聚糖類底物,水解甘蔗渣來源的木聚糖能力最強,以水解甘蔗渣來源的木聚糖時測得的TgXyn2酶活性為100%,水解小麥麩質來源和燕麥漿來源的木聚糖分別達到86.92%和59.66%;TgXyn2對微晶纖維素MCC這類非木聚糖底物基本沒有表現出水解活性(圖7)。類似的還有馬騰飛等[15]研究的在畢赤酵母中表達的木聚糖酶XynB,該木聚糖酶同樣特異性較強,對可溶性淀粉、微晶纖維素、魔芋苷聚糖、低聚果糖完全不能水解。說明這兩類木聚糖酶在水解方面有很強的特異性,其活性部位的構象只與特定的底物發生互補結合,能夠專一地水解木聚糖。

圖7 不同底物對應酶活大小Fig.7 The enzyme activity corresponding to different substrates
2.3 TgXyn2的同源蛋白結構預測及序列分析
通過在NCBI網站(https://blast.ncbi.nlm.nih.gov/Blast.cgi)上將TgXyn2的序列進行序列對比,我們發現TgXyn2與哈茨木霉CBS 226.95的序列相似度為100%,目前并沒有關于哈茨木霉CBS 226.95的相關研究報道。TgXyn2的結構域和同源蛋白結構分析結果(圖8)和TgXyn2的序列比對分析結果(圖9)表明,TgXyn2與GH11家族關系密切,分析TgXyn2中所有可能作為催化殘基的保守氨基酸,只有Glu 172和Glu 81位于酶與底物作用的活性中心[19],這些位點可能為該蛋白的活性中心。
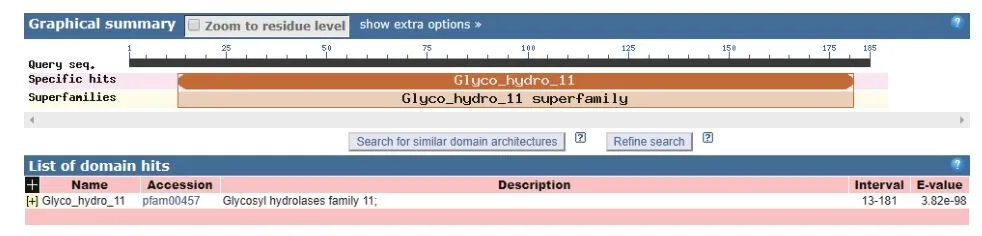
圖8 TgXyn2一級結構Fig.8 Primary structure of TgXyn2

圖9 TgXyn2序列比對示意圖Fig.9 Sequence alignment of TgXyn2
對TgXyn2的三維結構進行建模,由三維結構(圖10)可知,木聚糖酶TgXyn2整個酶分子呈右手型結構,由1個α-螺旋和2個反向平行的β-折疊構成。GH11家族木聚糖酶的催化機制是經典的酸堿催化機制[20],由圖可知該酶的活性中心Glu 172和Glu 81在反應隧道內,因此具有極大的改造空間,例如可通過導入二硫鍵等突變手段使得反應隧道打開,增大活性中心的接觸面,提高該酶的活性。

圖10 TgXyn2三級結構示意圖Fig.10 Schematic diagram of TgXyn2 tertiary structure
3 討論與結論
當前關于木聚糖酶的研究頗多,但由于木聚糖酶的多樣性和復雜性,以及自然界發現的木聚糖酶多為堿性木聚糖酶等因素,使得木聚糖酶在食品等行業中的應用有待提高。對木聚糖酶的熱穩定性和酸堿穩定性等方面進行研究和改造,不僅對理解蛋白結構與功能的關系有重要意義,而且能為生產應用帶來高效性和經濟效益。例如目前木聚糖酶領域的一項研究熱點:運用蛋白質工程技術手段對木聚糖酶進行分子改造[21],讓位于N末端“手掌”區域的取代殘基和突變體中新生成的氫鍵來幫助提高酶的熱穩定性。穩定性的顯著提高將為木聚糖酶在不同生產領域的應用鋪平道路[22]。
本文所研究的TgXyn2是從貴州木霉菌株(Trichoderma guizhouense)中發現的低溫耐酸性木聚糖酶,分子量為20.19 kDa,可以有效降解木聚糖主鏈的β-1,4木糖苷鍵。TgXyn2的酶促反應最適作用溫度與宋文芳等[23]從若爾蓋沼澤的草甸淤泥中分離篩選出能同步產生低溫纖維素酶和低溫木聚糖酶的低溫厭氧纖維素菌CD-2所產木聚糖酶接近,均為35 ℃;最適pH與鄭亞倫等[24]對重組菌株BA-TB-1進行誘導表達獲得的木聚糖酶接近;與已報道[25-26]的木霉(Trichoderma)及黑曲霉(Aspergillus niger)所產木聚糖酶相比,本實驗所研究的木聚糖酶TgXyn2的最適pH更接近中性。
在pH3~7的范圍內、尤其是在pH4~6的范圍內TgXyn2能夠保持較高的酶活力,即使是在雞的肌胃即pH2.5~3的酸性環境內,該酶的酶活力下降依舊不顯著,說明家禽的消化道內酸堿度環境對酶TgXyn2的活性影響較小[27-29]。然而木聚糖酶TgXyn2的最適溫度在35 ℃,在超過40 ℃的環境中,酶活力下降很快,因此不能完全適應家禽的胃腸道溫度。如將酶TgXyn2用作肉雞的基礎飼糧食品等行業,還需對TgXyn2進行必要的熱穩定性改造。
木聚糖酶TgXyn2具備一定的耐酸性,但其熱穩定性還需進一步改良。綜合來看,酶TgXyn2在食品、造紙、紡織等行業有一定的應用價值。例如Liu等[30]研究了巴氏擬桿菌(Paenibacillus barengoltzii)產的新型木聚糖酶的生化特性,并以此指導其在玉米芯和甘蔗渣生產低聚木糖中的應用,本研究發現TgXyn2對于甘蔗渣來源的木聚糖分解能力較強,在食品加工活動中有發揮作用的潛力。木聚糖酶家族資源豐富,應用廣泛,未來發展潛力巨大,除上述食品飼料等行業外,它還非常廣泛地應用于環境保護[31]和農產品加工等諸多行業中。隨著木聚糖酶理論基礎的繼續發展,應用研究也越來越受到重視,木聚糖酶的研究已成為糖化學催化領域中十分重要的分支,在食品、農業、新能源、醫藥等領域[32]具有優秀的研究潛力和廣闊的應用前景。
