不同鐵濃度對一種顆石藻Emiliania huxleyi的生長及光合作用的影響
2022-03-01 07:25:06任慶敏林燕妮王寅初
海洋科學 2022年12期
任慶敏, 林燕妮, 王寅初, 秦 松
不同鐵濃度對一種顆石藻的生長及光合作用的影響
任慶敏1, 2, 林燕妮4, 王寅初1, 3, 5, 秦 松1, 3, 5
(1. 中國科學院煙臺海岸帶研究所, 山東 煙臺 264003; 2. 中國科學院大學, 北京 100049; 3. 國家基礎學科公共科學數據中心, 北京 100190; 4. 煙臺大學, 山東 煙臺 264005; 5. 中國科學院海洋大科學研究中心, 山東 青島 266071)
為探討鐵對顆石藻生長及光合作用的影響, 以分離于秘魯上升流區域的兩株不同基因型的赫胥黎艾氏藻RCC6660與RCC6666為研究對象, 在調整5種鐵濃度(0、50、100、500、1 000 nmol/L)的Aquil培養基中培養, 比較其生長率、色素含量、營養鹽消耗、顆粒有機物含量及葉綠素熒光特性參數的差異。實驗結果表明: 鐵濃度未對兩株藻的生長產生顯著影響, 但0 nmol/L時RCC6660的比生長率顯著高于RCC6666(<0.05)。隨著鐵濃度的減少, 細胞粒徑和體積減小, 最大電子傳遞速率(rETRmax)和表觀光能利用效率降低。我們推測RCC6660為了保護細胞在低鐵時免受光損傷, 通過葉黃素循環以及高度鈣化的外殼實現光保護, 而RCC6666通過更多其他形式的非光化學熒光淬滅的手段。由于鈣化是個耗能過程, 低鈣化的RCC6666的光能利用效率在低鐵濃度下也更高。但RCC6660的rETRmax更大, 擁有較高的光合潛力, 這可能與其擁有的更高含量的色素有關。本研究揭示了不同基因型、不同鈣化程度的兩株對不同鐵濃度的差異響應, 及鐵限制條件下光保護機制和光能利用的不同, 為進一步研究光合和鈣化作用的偶聯機制提供了參考。
顆石藻; 鐵限制; 光合; 葉綠素熒光
顆石藻是一種單細胞的光合自養鈣化微藻, 屬于定鞭藻, 是海洋初級生產力的重要組成部分[1]。當今世界海洋中碳酸鈣的年產量約為50億噸, 從中生代開始, 顆石藻一直是世界海洋中主要的碳酸鈣生產者, 如今約占海洋碳酸鈣總產量的三分之一[2-3]。光照、溫度、pH、鹽度以及氮、磷營養鹽和鐵等環境因子是影響顆石藻的種類、密度和分布的主要原因[4]。鐵是海洋生態體系中有機體所必需的微量元素, 其對于藻類的新陳代謝、光合作用和呼吸過程中電子的轉移、硝酸鹽的還原、葉綠素的合成以及活性氧自由基的解毒來說都是必需的元素[5]。在寡營養大洋中, 鐵的可用性限制了浮游生物的生長, 從而大大降低了生物碳泵的效率。
國內外在鐵對浮游植物光合作用及其他生理生化特征的影響方面已經進行了較多的研究, 但多數實驗都圍繞藍藻和硅藻, 對于顆石藻的研究很少。鐵限制條件下, 硅藻的細胞體積會減小[6]或增大[7]。缺鐵會部分阻斷光系統II(PSII)和光系統I(PSI)之間的電子傳遞, 這是電子供體細胞色素c6濃度較低所致。這種受阻的電子傳遞可能導致質體醌池的過度減少, 引起PSII和PSI復合物的光化學效率顯著降低[8]。鐵限制導致細胞葉綠素濃度降低[9-10], 色素結合蛋白含量降低[11], 但體內葉綠素特異性光吸收截面增加[12]。
研究發現很多藻類在鐵限制的條件下長期進化而存在一些適應機制。比如硅藻擁有一系列應對鐵限制的策略, 使它們能夠在低鐵條件下存活, 并響應間歇性鐵輸入而迅速產生水華[13-14]。這些機制包括通過減少富含鐵的光合蛋白復合物的豐度來減少代謝鐵需求[15]; 用無鐵蛋白替代含鐵蛋白[9, 16]; 利用高親和力鐵攝取系統; 使用鐵儲存機制實現奢侈的鐵攝取量(即超過實現最大增長率所需的量); 最高的σPSII值和更大的細胞體積[17]等等。
顆石藻相比于硅藻, 似乎具有更少的鐵需求量, 但是鐵對于赫胥黎艾氏藻的DNA修復、ROS管理以及維持光合作用的最佳狀態是必不可少的, 間接影響碳循環和未來的生態系統功能。以時間和濃度依賴的方式攝取Fe(III)形式的鐵[18], 且其生長所需的鐵含量很低, 在含5.4 nmol/L鐵的培養基中即可維持較好的生長速率[19]。鐵限制條件下活性CO2和HCO3–的攝取率顯著降低, 導致生長速率顯著降低[20]。與鐵限制條件相比, 在高鐵條件下, 色素和光合參數增加, 活性氧(ROS)的積累減少, 由過量輻照度引起的DNA損傷最小化[21]。
秘魯海流系統具有全年持續的沿海上升流, 是最具生產力的海洋生態系統之一, 秘魯近海的缺氧和靜海水中, 沉積物來源的還原鐵[Fe(Ⅱ)]構成了溶解鐵的主要組成, 濃度高達200 nmol/L[22]。然而, 高營養的秘魯洋流系統中的浮游植物卻可能受到缺鐵的限制[23-24], 而且上升流的酸性環境可能加深了鐵限制的影響[25]。近岸上升流區域中的顆石藻在整個適應過程中可能同時經歷了鐵限制與富鐵環境, 其對鐵的適應范圍及對其光合作用的影響需要進一步探索。本文選取了秘魯近岸分離的RCC6660和RCC6666為研究對象, 它們屬于不同的基因型且有著不同的鈣化能力。通過比較不同鐵濃度對它們的生長及光合作用的影響, 探究不同藻株的差異響應表現及低鐵適應機制, 進一步揭示鈣化外殼在低鐵條件下可能對細胞的光保護作用。
1 材料與方法
1.1 實驗材料
1.1.1 藻種
本實驗所用的顆石藻為赫胥黎艾氏藻RCC6660和RCC6666, 由法國Roscoff藻種中心(http://roscoff-culture-collection.org/)提供。原藻種分離于秘魯上升流中心, 當地的pH最低為7.65。
1.1.2 鐵的控制
本實驗所用培養基是為研究微量金屬元素對藻類生理影響而設計的Aquil培養基, 培養基的配制及處理方法參照Price等[26]。其中人工海水及營養鹽(NaH2PO4·H2O、NaNO3和Na2SiO3·9H2O)分別使用Chelex 100(Bio-Rad, US)陽離子交換樹脂除去溶劑和試劑中引入的鐵污染。維生素及用100 μmol/L EDTA螯合的微量金屬元素母液使用酸洗的針頭過濾器過濾除菌。鐵使用EDTA螯合后添加, 培養基的鐵濃度分別設置為0、50、100、500、1 000 nmol/L。
實驗中所用的水為超純水(Milliphore, >18 MΩ·cm–1), 所有容器都經過酸洗, 然后用Q-H2O重復洗滌五次以上[27]。為防止高溫滅菌引入的鐵污染, 培養基使用0.22 μm孔徑的一次性無菌過濾器過濾除菌, 棄掉前200 mL濾液。
1.1.3 培養條件
培養基在使用之前, 將鹽度調整為35, 通過添加NaHCO3將溶解無機碳(DIC)調整為2 000 μmol·kg–1。藻株使用密閉蓋的175 cm2聚碳酸酯細胞培養瓶進行培養, 光照條件為100 μmol photons·m–2·s–1(14L︰10D), 溫度為20 ℃。將封閉的培養物每天搖動3次并隨機交換位置以確保細胞受光均勻。在正式實驗之前, 細胞在對應的鐵濃度下進行兩次適應性馴化, 即讓細胞從103cells/mL增殖至5×104cells/mL(不大于105cells/mL), 以適應實驗條件。在正式實驗時, 以103cells/mL的濃度接種, 當細胞濃度到達5× 104cells/mL(不大于105cells/mL)時測量光合生理并收樣測定其他參數。
1.2 實驗方法
1.2.1 細胞生長率與粒徑、體積
每天固定時間使用0.1 mL浮游生物計數框對細胞計數來確定細胞密度。比率生長速率()如下測定:=(ln2–ln1)/(2–1), 其中2和1分別代表2和1的細胞濃度。實驗結束后取樣, 使用流式細胞攝像系統FlowCAM(fluid imaging technologie, US)測量細胞粒徑及體積。
1.2.2 細胞色素
在GF/F濾膜(Millipore, US)上過濾100 mL培養物, 將膜折疊后放入離心管中, 加入4 mL 100%甲醇并置于4 ℃冰箱中提取過夜, 然后在4 ℃下離心10分鐘(6 700 r/min)。使用掃描分光光度計掃描上清液在200~800 nm的吸收[28]。使用下列公式計算葉綠素()[29]、葉綠素()[30]和類胡蘿卜素()[31]的濃度(μg·mL–1):
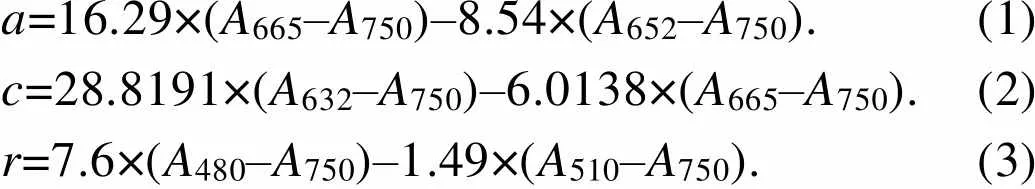
a=16.29×(A665–A750)–8.54×(A652–A750).(1) c=28.8191×(A632–A750)–6.0138×(A665–A750).(2) r=7.6×(A480–A750)–1.49×(A510–A750).(3)
1.2.3 碳酸鹽系統及營養分析
用玻璃纖維過濾器(Whatman GF/F)過濾所得的濾液進行DIC和營養鹽測定。DIC使用總有機碳分析儀(Shimadzu TOC-VCPH)測量; 使用連續流動分析儀(Seal-Branlubbe AA3, Seal Germany)測量硝酸鹽和磷酸鹽的濃度。
1.2.4 顆粒有機碳(POC)及顆粒有機氮(PON)含量測定
顆石藻細胞使用預燃燒(450 ℃, 12 h以上)的玻璃纖維濾膜通過抽濾收集, 每組收集100 mL。測量前添加230 μL的 0.1 mol/L鹽酸以除去膜上的無機碳, 用以測量POC和PON。濾膜在60 ℃烘箱中烘干2 h之后, 使用錫紙包裹并在元素分析儀中進行測定。將細胞的PON和POC含量乘以生長速率(), 以計算PON生產速率(PIN)和POC生產速率(POC)。
1.2.5 葉綠素熒光
藻細胞暗適應15 min之后, 使用葉綠素熒光儀(AquaPen-C, 捷克PSI)測量葉綠素熒光參數, 包括最大光化學效率(Fv/Fm)、有效光化學效率(YPSII)和非光化學淬滅(NPQ)。
快速光響應曲線設置6個光強梯度(10, 20, 50, 100, 300, 500 μmol photons·m–2·s–1), 每個光強60 s, 相對電子傳遞速率(rETR)的計算公式如下[32]:
=YPSII××0.5, (4)
式中,指代提供光化學光的光合有效輻射(PAR),為rETR。將rETR—PAR數據導入到SigmaPlot (v13.0)后, 使用雙指數衰減(double exponential decay)方程進行回歸擬合[33-34], 并得到各項特征參數, 包括最大相對電子傳遞速率(rETRmax)、飽和光強和表觀光能利用率。
1.3 數據處理
采用IBM SPSS Statistics 26.0數據統計軟件進行數據, 不同鐵濃度處理間的差異用方差分析、多重比較, 兩個藻種之間的差異用-檢驗分析, 以<0.05作為顯著性差異。
2 結果與分析
2.1 兩株顆石藻的進化分析
按照Bendif 根據核糖體系統發育定義的三個分支[35], 將RCC6660與RCC6666與其他藻株一起進行系統發育分析(圖1)。結果表明, RCC6660屬于α支, 而RCC6666屬于β支(圖1)。因此本文中, 為了行文簡潔, 赫胥黎艾氏藻RCC6660與RCC6666分別簡稱為α株與β株。
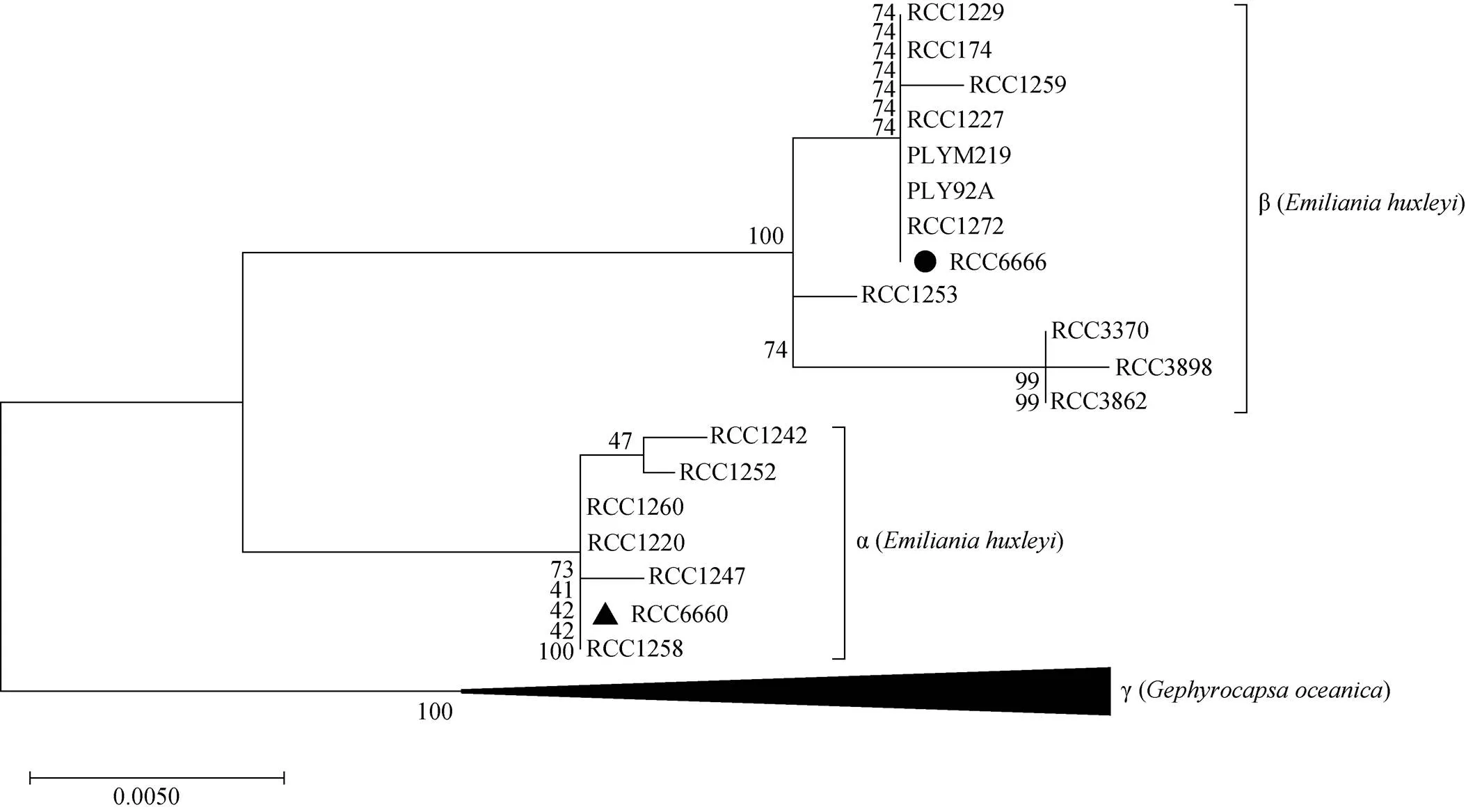
圖1 基于極大似然法利用線粒體cox3基因序列的艾氏藻系統發育樹66606666
注: 進化枝顯示100次重復的自展值支持。藻株RCC6660和RCC6666在赫胥黎艾氏藻兩個基因型分支(α和β)中的位置分別用黑色三角和黑色圓標記。外群(即γ枝)由大洋橋石藻()RCC1281、RCC1316、RCC1307、RCC1839、RCC1320、RCC1319、RCC1318、RCC1317、RCC1306、RCC1305、RCC1300、RCC1286、RCC1284、RCC1282、RCC1562、RCC1292組成
2.2 對顆石藻生長率的影響
如圖2所示, α株的比生長率隨著培養基中鐵濃度的增加表現出遞增的趨勢, 所以其在鐵濃度為1 000 nmol/L時比生長率大于其他組(>0.05), 而對于β株, 50 nmol/L才是其最適鐵濃度(0.05)。鐵濃度為0 nmol/L時, α株的比生長率顯著高于β株(< 0.05), 而在鐵濃度為50 nmol/L時, α株的比生長率顯著低于β株(<0.05)。
2.3 對細胞粒徑及體積的影響
如圖3所示, 兩藻株的細胞最小粒徑在鐵濃度較低時較小, 而在鐵濃度大于500 nmol/L時趨于穩定。在鐵濃度為0 nmol/L時細胞的細胞粒徑最大(>0.05), 高鐵濃度(>100 nmol/L)反而抑制了細胞的最大粒徑, 使其表現為減小的趨勢。在不同鐵濃度培養中, α株的最小粒徑均小于β株, 且在0 nmol/L和100 nmol/L時具有顯著性(<0.05), 而α株的最大粒徑均大于β株(>0.05)。兩藻株在鐵濃度為100 nmol/L時, 細胞平均粒徑和平均體積最大(>0.05)。且在各個鐵濃度中, α株的平均體積都更大(>0.05)。

圖2 五種實驗鐵濃度對RCC6660(α)和RCC6666(β)的比生長率的影響
注: 數值為平均值±標準差,=3。不同字母表示單一藻株不同鐵濃度之間具有顯著性差異(<0.05), 柱狀圖上的橫線表示兩個藻株之間有顯著性差異(<0.05)

圖3 五種實驗鐵濃度對RCC6660(α)和RCC6666(β)的細胞粒徑和細胞體積的影響
注: 數值為平均值±標準差,=3
2.4 對顆石藻色素含量的影響
2.4.1 不同鐵濃度對葉綠素的影響
不同鐵濃度對顆石藻葉綠素的含量的影響不大, 各個實驗組之間不具有顯著性差異(見圖4a)。兩者比較, 在較低鐵濃度(0、50、100、500 nmol/L)下, α株細胞中的含量更高, 而β株在1 000 nmol/L時的葉綠素含量更高。
2.4.2 不同鐵濃度對葉綠素的影響
如圖4b所示, 在0、50和100 nmol/L時, α株的葉綠素含量高于β株, 而在500和1 000 nmol/L時, α株的葉綠素含量顯著低于β株(<0.05)。且β株在1 000 nmol/L時的葉綠素含量顯著高于50 nmol/L (<0.05)。
2.4.3 不同鐵濃度對類胡蘿卜素的影響
如圖4c所示, 在0、50和100 nmol/L時, α株的類胡蘿卜素含量高于β株, 而在500和1 000 nmol/L時, α株的類胡蘿卜素含量低于β株, 與葉綠素含量變化一致, 且在1 000 nmol/L時存在顯著性差異(<0.05)。α株在100 nmol/L鐵濃度時的類胡蘿卜素含量顯著高于500 nmol/L和1 000 nmol/L(<0.05)。β株在0和50 nmol/L時的類胡蘿卜素含量顯著低于其他濃度的含量(<0.05)。

圖4 五種實驗鐵濃度對RCC6660(α)和RCC6666(β)中的葉綠素a、葉綠素c及類胡蘿卜素的影響
注: 數值為平均值±標準差,=3。不同字母表示單一藻株不同鐵濃度之間具有顯著性差異(<0.05), 柱狀圖上的橫線表示兩個藻株之間有顯著性差異(<0.05)
2.5 對顆石藻氮磷消耗的影響
如表1所示, 培養基中鐵濃度為50 nmol/L時, 培養結束后培養基中剩余的磷含量最高, 之后隨著鐵濃度的增加磷消耗加劇(>0.05)。但是在鐵濃度0 nmol/L的培養基中培養的細胞竟然消耗了比高濃度鐵環境中更多的磷。α株比β株在同樣的條件下消耗的磷多。根據氮鹽的剩余濃度可知, 硝酸鹽的消耗呈現先增后減再增的狀態。
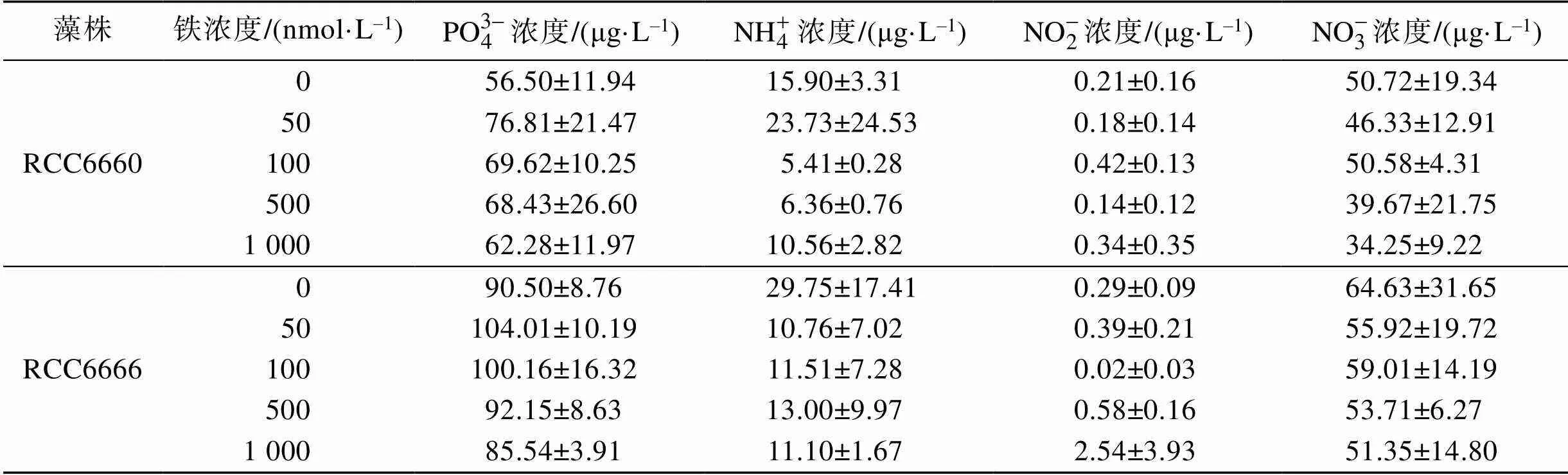
表1 五種實驗鐵濃度對RCC6660(α)和RCC6666(β)的氮磷消耗的影響
注: 數據為實驗結束后培養基中的氮(稀釋50倍)與磷(稀釋2倍)含量, 起始值相同。表中數據為三次重復的平均值±標準差,=3
2.6 對顆石藻光合作用的影響
不同鐵濃度條件下培養的α株藻細胞PSII活性沒有顯著性差異, 但是β株在鐵濃度為100 nmol/L時的活性顯著低于500 nmol/L時(<0.05)。而且在鐵限制條件(0 nmol/L)下, α株的v/m顯著低于β株(<0.05)(圖5a)。

圖5 五種實驗鐵濃度對RCC6660(α)和RCC6666(β)的最大光化學效率、有效光化學效率和非光化學淬滅的影響
注: 數值為平均值±標準差,=3。不同字母表示單一藻株不同鐵濃度之間具有顯著性差異(<0.05), 柱狀圖上的橫線表示兩個藻株之間有顯著性差異(<0.05)
從圖5b可以看出, 在五種實驗鐵濃度下, β株的NPQ都大于α株, 且在鐵濃度較低的培養條件(0、50和100 nmol/L)下, β株的NPQ顯著大于α株(<0.05)且呈較高水平。α株的NPQ隨鐵濃度增加表現為遞增趨勢。
如圖5c所示, 兩個藻株相比, β株在實驗鐵濃度下的PSII有效光化學效率更大, 且在50和100 nmol/L時是顯著的(<0.05)。β株在500 nmol/L時的有效光化學效率顯著高于在0 nmol/L和1 000 nmol/L的條件下的(<0.05)。
從圖6可以看出, 在5種實驗鐵濃度下, β株的表觀光合效率均高于α株, 且最大電子傳遞速率在較高鐵濃度(100、500和1 000 nmol/L)時也較高, 但是低濃度時反之。兩者表觀光合效率和最大電子傳遞速率均無顯著性差異。總體來講, 鐵濃度高時, 表觀光合效率和最大電子傳遞速率也呈較高的水平。

圖6 五種實驗鐵濃度對RCC6660(α)和RCC6666(β)的表觀光合效率、最大電子傳遞速率的影響及快速光響應曲線
注: 數值為平均值±標準差,=3
2.7 對顆石藻顆粒有機物生產的影響
如圖7所示, α株的PON含量以及POC與PON的生產率在五種實驗鐵濃度下均大于β株, 且在500 nmol/L時POC、PON含量和產率達到最大值。在五種實驗鐵濃度下, β株的POC、PON含量和產率都呈現先減后增再減的趨勢。兩株藻的POC/PON值也表現出這個趨勢且在50 nmol/L和500 nmol/L時具有顯著性差異(<0.05)。
3 討論
3.1 鐵濃度與細胞粒徑
南美洲秘魯沿岸的近海洋流所經海域是一個廣闊的、高硝酸鹽的但鐵有限的葉綠素區域, 該地區底層水中有極高濃度的溶解鐵(>50 nmol/L), 在秘魯南海岸沿岸上升流水域和秘魯洋流近岸海域中, 溶解鐵濃度很低(<0.1 nmol/L)[36]。顆石藻可能在這片水域逐漸適應, 因而在廣范圍的鐵濃度下的生長率都趨于穩定。在鐵濃度較低(0、50、100 nmol/L)時, 兩藻株的細胞粒徑和體積隨著鐵濃度的降低而減小, 細胞的比表面積增大, 進而促進了細胞對鐵的吸收。此外, 細胞體積小對于生活在低鐵環境中的細胞特別有利, 因為它降低了細胞對于鐵、氮和光合產物的需求[37], 而且降低了下沉率。總體來看, 在所有鐵濃度下, β株的細胞粒徑和體積低于α株, 這暗示它可能對鐵有更高的要求, 或者對鐵濃度的變化反應更敏感。后者也可以從其在色素和顆粒有機物等參數隨鐵濃度的增加呈現“波浪式”變化的表現可以看出。

圖7 五種實驗鐵濃度對RCC6660(α)和RCC6666(β)的顆粒有機碳與顆粒有機氮含量及產率、兩者比值的影響
注: 數值為平均值±標準差,=3。不同字母表示單一藻株不同鐵濃度之間具有顯著性差異(<0.05), 柱狀圖上的橫線表示兩個藻株之間有顯著性差異(<0.05)
3.2 氮磷營養的消耗
細胞減少鐵需求的另一種方法是使用還原態的氮代替氧化態的氮, 因為硝酸鹽的還原需要含鐵酶, 如硝酸鹽和亞硝酸鹽還原酶[14, 38]。本實驗中采用硝酸鹽作為氮源, 所以鐵需求量可能會相對增大[39]。隨著鐵濃度變化, 兩株藻對于氮磷的消耗趨勢一致。在0 nmol/L的鐵限制環境中, 細胞對于硝酸鹽的消耗比1 000 nmol/L的富鐵條件下低, 但卻消耗了與富鐵條件下差不多的磷。在五種實驗鐵濃度下, α株都比β株消耗的氮磷要多。這表明, α株比β株對氮磷的需求更多, 鐵限制條件會降低細胞對于硝酸鹽的利用但是不影響磷的利用。鐵限制條件下, 細胞可能是部分氮脅迫的。
50 nmol/L下兩株藻細胞對硝酸鹽的吸收表現出了一個小的峰值, 而對磷的需求卻達到了最小值, 此時消耗的N︰P較大。β株在50 nmol/L鐵濃度下細胞的PON值顯著降低, 表明細胞不能維持其最佳的氮配額。而且盡管C︰N比增加了, POC含量也減少了, 這與之前在硝酸鹽中生長的細胞在鐵脅迫下表現得一致[37, 40]。然而α株不具備此特征, 其在鐵濃度為500 nmol/L時細胞含有較高的POC及PON含量但是 C︰N仍顯著小于β株。鐵會促進碳和氮同化成葉綠素和儲能化合物等成分[9, 38, 41], 其機制尚不清楚, 凈結果是促進了有機質的積累。
3.3 色素與光保護
受鐵限制的硅藻降低了葉綠素含量、光合效率[42-43], 所有這些都導致了生長速度的降低。然而判斷鐵限制的重要指標v/m[44]以及比生長率[9, 45-46]都沒有顯著的變化, 或許可以說明顆石藻對鐵的極低需求度[19]。兩個藻株在不同鐵濃度下的細胞色素含量呈現一定的規律。α株在鐵濃度為100 nmol/L 時葉綠素、葉綠素和類胡蘿卜素的總含量是最多的, 而β株在鐵濃度為1 000 nmol/L 時總含量是最多的。這也在一定程度上證明了兩株藻鐵需求的不同。
細胞中光合色素含量下降似乎是鐵脅迫下的普遍行為[44, 47-49], 但在我們的實驗中, 葉綠素含量并沒有顯著降低。色素濃度的這種相對不敏感是鐵限制的謎團之一, 尤其是在大量營養素水平較高時[45-46]。但是類胡蘿卜素含量變化很大。β株在鐵限制條件下類胡蘿卜素含量顯著降低, 然而α株正好相反, 在富鐵環境中細胞的類胡蘿卜素含量較低。為什么會產生這樣的差異?
鐵限制條件下, 真核藻類主要通過葉黃素循環相關的NPQ來耗散葉綠素吸收的過多的過剩光能, 在高光低鐵條件下, 非光化學淬滅對光系統的保護作用更加明顯且重要[10, 47-49]。類胡蘿卜素能通過葉黃素循環調節天線的能量傳遞, 還在防御光合結構的光氧化破壞和捕光復合體的裝配方面發揮重要作用。鐵限制條件下, β株的NPQ要顯著大于α株。這可能是由于α株是一種高度鈣化的細胞[50], 顆石粒組成的外骨骼可以抵擋一部分的紫外光[51], 在鐵限制條件下, 高含量的類胡蘿卜素及厚的碳酸鈣外殼都為細胞在高強度PAR/UVR下提供了多重保護。而β株可能更多地選擇增強NPQ的能力以增加熱耗散。而這種NPQ可能不是來自葉黃素, 而是捕光天線復合體在PSI與PSII之間的移動或PSII反應中心的淬滅[52]。β株在低鐵時趨于生產更少量的類胡蘿卜素, 這似乎也是一種光保護手段, 因為捕光色素的增加抑制PSII的修復速率, 加劇光抑制[53]。
3.4 光合電子傳遞
類囊體是鐵主要存在的部位, 類囊體電子傳遞的幾乎每一個方面都依賴于鐵。鐵由于其高還原-氧化電位而在許多金屬蛋白中作為電子載體[39]。在浮游植物中, 高比例的鐵包含在光合作用裝置和線粒體電子傳遞鏈的組件中[54-55]。鐵限制導致最大電子傳遞速率和表觀光能利用效率都隨鐵濃度的降低而降低[56-57]。本文的結果也表現出了這個趨勢, 但是鐵濃度為1 000 nmol/L時顆石藻最大電子傳遞速率下降, 可能是此濃度超過了光合作用的最適濃度甚至有抑制作用。鐵脅迫下細胞捕獲和利用光能的能力較弱, 因此缺鐵的細胞可能比富鐵的細胞更容易受到光化學損傷[5], 這與本實驗中光飽和參數也在低鐵時降低一致, 細胞在0 nmol/L下耐受強光的能力較弱。
β株在任何鐵濃度時都保持更高的光能的利用效率, 這可能也與其鈣化作用弱于α株相關, 畢竟鈣化是個耗能過程[58]。在鐵限制條件(0 nmol/L, 50 nmol/L)下, α株的最大相對電子傳遞速率rETRmax更大, 擁有較高的光合潛力, 當高鐵濃度時β株才占據優勢。這可能是由于其低鐵時更高的葉綠素含量, α株可能通過過度生產光合作用天線, 增加光子聚焦來克服短期的電子傳輸效率低下, 從而彌補對效率較低的無鐵蛋白質的持續依賴。
4 結論
海洋浮游生物的鐵供應變化會通過改變碳固存率對大氣中二氧化碳的濃度產生重大影響[59], 而顆石藻作為主要的鈣化浮游生物, 了解鐵限制對其的影響是很重要的。本研究通過比較不同鐵濃度對兩株不同鈣化程度且不同基因型的顆石藻生長及光合作用的影響, 證明了顆石藻通過降低細胞粒徑等策略來適應缺鐵條件。同時缺鐵影響了色素合成及電子傳遞, 顆石藻為了應對鐵限制產生了一系列光保護機制。因為兩株藻鈣化程度的差異造成了鐵限制條件下的差異響應, 因此在之后的研究中, 需要進一步證實是不是鈣化作用的差異導致了光合方面的差異表現。
[1] HAY W W, MOHLER H P, ROTH P H, et al. Calcareous nannoplankton zonation of the cenozoic of the gulf coast and caribbean-antillean area, and transoceanic correlation[J]. Protein Science A Publication of the Protein Society, 1967, 19(9): 1649-1661.
[2] IGLESIAS-RODRIGUEZ M D, HALLORAN P R, RICKABY R E M, et al. Phytoplankton calcification in a high-CO2world[J]. Science, 2008, 320(5874): 336-340.
[3] MILLIMAN J D. Production and accumulation of calcium-carbonate in the ocean: budget of a nonsteady state[J]. Global Biogeochemical Cycles, 1993, 7(4): 927-957.
[4] YANG T N, WEI K Y, GONG G C. Distribution of coccolithophorids and coccoliths in surface ocean off northeastern Taiwan[J]. Botanical Bulletin of Academia Sinica, 2001, 42(4): 287-302.
[5] GEIDER R J, LAROCHE J. The role of iron in phytoplankton photosynthesis, and the potential for iron- limitation of primary productivity in the sea[J]. Photosynthesis Research, 1994, 39(3): 275-301.
[6] LEYNAERT A, BUCCIARELLI E, CLAQUIN P, et al. Effect of iron deficiency on diatom cell size and silicic acid uptake kinetics[J]. Limnology and Oceanography, 2004, 49(4): 1134-1143.
[7] TAKEDA S, OBATA H. Response of equatorial pacific phytoplankton to subnanomolar fe enrichment[J]. Marine Chemistry, 1995, 50(1/4): 219-227.
[8] RONCEL M, GONZáLEZ-RODRíGUEZ A A, NARANJO B, et al. Iron deficiency induces a partial inhibition of the photosynthetic electron transport and a high sensitivity to light in the diatom[J]. Frontiers in Plant Science, 2016, 7: 1050.
[9] ALLEN A E, LAROCHE J, MAHESWARI U, et al. Whole-cell response of the pennate diatomto iron starvation[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(30): 10438-10443.
[10] VAN DE POLL W H, VAN LEEUWE M A, ROGGEVELD J, et al. Nutrient limitation and high irradiance acclimation reduce PAR and UV-induced viability loss in the Antarctic diatom(Bacillariophyceae)[J]. Journal of Phycology, 2005, 41(4): 840-850.
[11] RUETER J G, UNSWORTH N L. Response of marine(cyanophyceae) cultures to iron nutrition[J]. Journal of Phycology, 1991, 27(2): 173-178.
[12] GREENE R M, GEIDER R J, KOLBER Z, et al. Iron- Induced changes in light harvesting and photochemical energy conversion processes in eukaryotic marine al-gae[J]. Plant Physiology, 1992, 100(2): 565-575.
[13] SUNDA W G, HUNTSMAN S A. Iron uptake and gro-wth limitation in oceanic and coastal phytoplankton[J]. Marine Chemistry, 1995, 50(1/4): 189-206.
[14] MARCHETTI A, SCHRUTH D M, DURKIN C A, et al. Comparative metatranscriptomics identifies molecular bases for the physiological responses of phytoplankton to varying iron availability[J]. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(6): E317-E25.
[15] STRZEPEK R F, HARRISON P J. Photosynthetic architecture differs in coastal and oceanic diatoms[J]. Nature, 2004, 431(7009): 689-692.
[16] PEERS G, PRICE N M. Copper-containing plastocyanin used for electron transport by an oceanic diatom[J]. Nature, 2006, 441(7091): 341-344.
[17] STRZEPEK R F, BOYD P W, SUNDA W G. Photosynthetic adaptation to low iron, light, and temperature in Southern Ocean phytoplankton[J]. Proceedings of the National Academy of Sciences, 2019, 116(10): 4388- 4393.
[18] HARTNETT A, BOTTGER L H, MATZANKE B F, et al. Iron transport and storage in the coccolithophore:[J]. Metallomics, 2012, 4(11): 1160- 1166.
[19] BOYE M, VAN DEN BERG C M G. Iron availability and the release of iron-complexing ligands by Emilia-nia huxleyi[J]. Marine Chemistry, 2000, 70(4): 277- 287.
[20] SCHULZ K G, ROST B, BURKHARDT S, et al. The effect of iron availability on the regulation of inorganic carbon acquisition in the coccolithophore Emiliania huxleyi and the significance of cellular compartmentation for stable carbon isotope fractionation[J]. Geochimica et Cosmochimica Acta, 2007, 71(22): 5301- 5312.
[21] SEGOVIA M, LORENZO M R, INIGUEZ C, et al. Physiological stress response associated with elevated CO2and dissolved iron in a phytoplankton community dominated by the coccolithophore[J]. Marine Ecology Progress Series, 2018, 586: 73-89.
[22] SCHLOSSER C, STREU P, FRANK M, et al. H2S events in the Peruvian oxygen minimum zone facilitate enhanced dissolved Fe concentrations[J]. Scientific Reports, 2018, 8(1): 12642-12650.
[23] HUTCHINS D A, HARE C E, WEAVER R S, et al. Phytoplankton iron limitation in the humboldt current and peru upwelling[J]. Limnology and Oceanography, 2002, 47(4): 997-1011.
[24] MOORE J K, DONEY S C, GLOVER D M, et al. Iron cycling and nutrient-limitation patterns in surface waters of the world ocean[J]. Deep-Sea Research Part II- Topical Studies in Oceanography, 2001, 49(1/3): 463- 507.
[25] SHI D L, XU Y, HOPKINSON B M, et al. Effect of ocean acidification on iron availability to marine phy-toplankton[J]. Science, 2010, 327(5966): 676-679.
[26] PRICE N M, HARRISON G I, HERING J G, et al. Preparation and chemistry of the artificial algal culture medium aquil[J]. Biological Oceanography, 1989, 6(5/6): 443-461.
[27] MALDONADO M T, PRICE N M. Influence of N substrate on Fe requirements of marine centric diatoms[J]. Marine Ecology Progress Series, 1996, 141(1/3): 161- 172.
[28] 繆杭斌. 鈣化與非鈣化顆石藻對海水酸化與UV輻射的生理學響應[D]. 廈門: 廈門大學, 2017.
MIAO Hangbin. Physiological responses of calcifying and non-calcifying coccolithophore strains to seawater acidification and UV radiation[D]. Xiamen: Xiamen University, 2017.
[29] PORRA R J. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b[J]. Photosynthesis Research, 2002, 73(1/3): 149-156.
[30] RITCHIE R J. Consistent sets of spectrophotometric chlorophyll equations for acetone, methanol and ethanol solvents[J]. Photosynthesis Research, 2006, 89(1): 27-41.
[31] STRICKLAND J, PARSONS T. A practical handbook of seawater analysis[M]. Ottawa: Fisheries Research Board of Canada, 1972.
[32] DUARTE B, SANTOS D, MARQUES J C, et al. Biophysical probing of Spartina maritima photo-system II changes during prolonged tidal submersion periods[J]. Plant Physiology and Biochemistry, 2014, 77: 122-132.
[33] EILERS P H C, PEETERS J C H. A model for the relationship between light-intensity and the rate of photosynthesis in phytoplankton[J]. Ecological Modelling, 1988, 42(3/4): 199-215.
[34] RALPH P J, GADEMANN R. Rapid light curves: A po-werful tool to assess photosynthetic activity[J]. Aquatic Botany, 2005, 82(3): 222-237.
[35] BENDIF E, PROBERT I, DIAZ-ROSAS F, et al. Recent reticulate evolution in the ecologically dominant lineage of coccolithophores[J]. Frontiers in Microbiology, 2016, 7: 784.
[36] BRULAND K W, RUE E L, SMITH G J, et al. Iron, macronutrients and diatom blooms in the Peru upwelling regime: brown and blue waters of Peru[J]. Marine Chemistry, 2005, 93(2/4): 81-103.
[37] MUGGLI D L, HARRISON P J. Effects of nitrogen source on the physiology and metal nutrition ofgrown under different iron and light conditions[J]. Marine Ecology Progress Series, 1996, 130(1/3): 255-267.
[38] MILLIGAN A J, HARRISON P J. Effects of non- steady-state iron limitation on nitrogen assimilatory en-zymes in the marine diatom(Bacillariophyceae)[J]. Journal of Phycology, 2000, 36(1): 78-86.
[39] RAVEN J A. The iron and molybdenum use efficiencies of plant-growth with different energy, carbon and nitrogen-sources[J]. New Phytologist, 1988, 109(3): 279- 287.
[40] MUGGLI D L, LECOURT M, HARRISON P J. Effects of iron and nitrogen source on the sinking rate, physiology and metal composition of an oceanic diatom from the subarctic Pacific[J]. Marine Ecology Progress Series, 1996, 132(1/3): 215-227.
[41] TIMMERMANS K R, STOLTE W, DEBAAR H J W. Iron-mediated effects on nitrate reductase in marine- phytoplankton[J]. Marine Biology, 1994, 121(2): 389- 396.
[42] GREENE R M, GEIDER R J, FALKOWSKI P G. Effect of iron limitation on photosynthesis in a marine diatom[J]. Limnology and Oceanography, 1991, 36(8): 1772-1782.
[43] MARCHETTI A, HARRISON P J. Coupled changes in the cell morphology and the elemental (C, N, and Si) composition of the pennate diatom Pseudo-nitzschia due to iron deficiency[J]. Limnology and Oceanography, 2007, 52(5): 2270-2284.
[44] BEHRENFELD M J, MILLIGAN A J. Photophy-sio-logical expressions of iron stress in phytoplankton[J]. Annual Review of Marine Science, 2013, 5(1): 217- 246.
[45] CARSLEY J E, FISHER A, MILLIGAN W W, et al. Mechanical behavior of a bulk nanostructured iron alloy[J]. Metallurgical and Materials Transactions a- Physical Metallurgy and Materials Science, 1998, 29(9): 2261-2271.
[46] MOSELEY J L, PAGE M D, ALDER N P, et al. Reciprocal expression of two candidate di-iron enzymes affecting photosystem I and light-harvesting complex accumulation[J]. Plant Cell, 2002, 14(3): 673-688.
[47] VAN LEEUWE M A, STEFELS J. Photosynthetic responses intowards varying light and iron conditions[J]. Biogeochemistry, 2007, 83(1/3): 61-70.
[48] ALDERKAMP A C, DE BAAR H J W, VISSER R J W, et al. Can photoinhibition control phytoplankton abundance in deeply mixed water columns of the Southern Ocean?[J]. Limnology and Oceanography, 2010, 55(3): 1248-1264.
[49] WILSON A, BOULAY C, WILDE A, et al. Light- induced energy dissipation in iron-starved cyano-ba-c-teria: roles of OCP and IsiA proteins[J]. Plant Cell, 2007, 19(2): 656-672.
[50] BEAUFORT L, PROBERT I, DE GARIDEL-THORON T, et al. Sensitivity of coccolithophores to carbonate chemistry and ocean acidification[J]. Nature, 2011, 476(7358): 80-83.
[51] MONTEIRO F M, BACH L T, BROWNLEE C, et al. Why marine phytoplankton calcify[J]. Science Advan-ces, 2016, 2(7): e1501822.
[52] BEHRENFELD M, MILLIGAN A. Photophysiological expressions of iron stress in phytoplankton[J]. Annual review of Marine Science, 2013, 5(1): 217-246.
[53] NISHIYAMA Y, ALLAKHVERDIEV S I, MURATA N. A new paradigm for the action of reactive oxygen species in the photoinhibition of photosystem II[J]. Biochi-mica et Biophysica Acta-Bioenergetics, 2006, 1757(7): 742-749.
[54] RAVEN J A, EVANS M C W, KORB R E. The role of trace metals in photosynthetic electron transport in O2- evolving organisms[J]. Photosynthesis Research, 1999, 60(2/3): 111-149.
[55] RAVEN J A. Predictions of Mn and Fe use efficiencies of phototrophic growth as a function of light availability for growth and of C assimilation pathway[J]. New Phytologist, 1990, 116(1): 1-18.
[56] PIETSCH D, BERNAT G, KAHMANN U, et al. New insights into the function of the iron deficiency-induced protein C fromPCC 7942[J]. Photosynthesis Research, 2011, 108(2/3): 121-132.
[57] PANKOWSKI A, MCMINN A. Iron availability regulates growth, photosynthesis, and production of ferredoxin and flavodoxin in Antarctic sea ice diatoms[J]. Aquatic Biology, 2009, 4(3): 273-288.
[58] JIN P, GAO K, BEARDALL J. Evolutionary responses of a coccolithophoridto ocean acidification[J]. Evolution, 2013, 67(7): 1869-1878.
[59] MARTIN J H, FITZWATER S E. Iron deficiency limits phytoplankton growth in the north-east Pacific subarctic[J]. Nature, 1988, 331(6154): 341-343.
Effects of different iron concentrations on the growth and photosynthesis of
REN Qing-min1, 2, LIN Yan-ni4, WANG Yin-chu1, 3, 5, QIN Song1, 3, 5
(1. Key Laboratory of Coastal Biology and Biological Resource Utilization, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai 264003, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. National Basic Science Data Center, Beijing 100190, China; 4. Yantai University, Yantai 264005, China; 5. Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China)
Two genotypes of, RCC6660 and RCC6666, isolated from a Peruvian upwelling, were cultured in Aquil medium at five different iron concentrations (0, 50, 100, 500, and 1, 000 nmol/L) to study the effect of iron on the growth and photosynthesis of coccolithophores. The difference in growth, pigment content, nutrient consumption, particulate organic matter content, and chlorophyll fluorescence characteristics were compared. The results revealed that iron concentration had no significant effect on the growth of the two strains; however, the specific growth rate of RCC6660 in 0 nmol/L was significantly greater than that of RCC6666. As the iron concentration decreased, cell size, cell volume, the maximum relative electron transport rate (rETRmax), and apparent quantum efficiency (α) also decreased. We speculate that RCC6660 was photoprotected by the lutein cycle and a highly calcified exoskeleton that protected the cells from light damage at low iron levels; moreover, RCC6666 was more likely to be quenched by other forms of non-photochemical fluorescence. The light energy efficiency of RCC6666 was higher at low iron concentrations, as less energy was devoted to calcification. However, RCC6660 had a higher rETRmaxand photosynthetic potential, which may be related to its higher pigment content. This study revealed the responses of twostrains with different genotypes and different calcification abilities to different iron concentrations and the differences in the photosynthetic protective mechanism and light energy utilization under an iron limitation, which provides a reference for further studies of the coupling mechanism between photosynthesis and calcification.
coccolithophores; iron limitation; photosynthesis; chlorophyll fluorescence
Nov. 9, 2020
P735; P76
A
1000-3096(2022)12-0050-13
10.11759/hykx20201109002
2020-11-09;
2020-12-18
國家基礎學科公共科學數據中心“中國海岸帶植物資源數據庫”(NBSDC-DB-22); 內蒙古自治區關鍵技術攻關計劃(2021GG0414)
[National Basic Science Data Center “Database of Coastal Plant Bioresources in China”, No. NBSDC-DB-22; Key Technology Program of Inner Mongolia Autonomous Region, No. 2016YFE0106700]
任慶敏(1996—), 女, 山東濟南人, 碩士, 主要從事海岸帶分子藻類學及基因組學研究, E-mail: hitqmren@163.com; 王寅初(1986—),通信作者, E-mail: ycwang@yic.ac.cn
(本文編輯: 楊 悅)
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
音樂探索(2022年2期)2022-05-30 21:01:37
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
