寧麥系列小麥品種的性狀特點及相關基因位點分析
2022-02-24 05:16:54姜朋張鵬姚金保吳磊何漪李暢馬鴻翔張旭
中國農業科學 2022年2期
姜朋,張鵬,姚金保,吳磊,何漪,李暢,馬鴻翔,張旭?
1江蘇省農業科學院/現代作物生產省部共建協同創新中心/CIMMYT-JAAS小麥病害聯合研究中心,南京210014;2揚州大學農學院/江蘇省糧食作物現代產業技術協同創新中心/江蘇省作物基因組學與分子育種重點實驗室/植物功能基因組學教育部重點實驗室,江蘇揚州 225009
0 引言
【研究意義】長江中下游麥區是中國第二大麥區[1],江蘇省農業科學院育成的寧麥系列品種在本區域小麥生產中占有重要地位,寧麥3號、寧麥6號、寧麥8號、寧麥9號、寧麥13、寧麥14等均是當地不同年代的主推品種,目前,尤其是寧麥13已連續5年成為江蘇省推廣面積最大的小麥品種(數據由江蘇省農技推廣總站提供);寧麥8號、寧麥9號、寧麥13等是常用的小麥育種親本,其中,寧麥9號的衍生品種已超過20個[2-3];寧麥8號、寧麥9號和寧麥13還是遺傳、生理等理論研究的重點材料[4-6]。對寧麥系列品種的性狀特點、系譜組成、遺傳多樣性及其重要性狀的控制位點等開展系統分析,可為育種利用提供理論指導,同時也有助于針對性地進行品種改良。【前人研究進展】張曉等[7]與王君嬋等[8]對揚麥系列品種的品質性狀、重要性狀功能基因等進行了系統分析,為揚麥系列品種在生產及育種中的應用提供了理論依據。殷貴鴻等[9]總結了周口市農業科學院小麥育種團隊2010年以來的科研成果及育種經驗,并提出下一步的育種目標與措施,對周麥系列品種的發展具有重要意義。關于寧麥系列品種的研究已有諸多報道[10-11],特別是對優良親本寧麥9號的研究較為深入,研究者初步明確了其產量、品質、抗病性等性狀的遺傳特點[3,12-14],并評價了其對后代的遺傳貢獻[2,15]。分子標記技術的發展為作物的遺傳研究提供了重要工具,特別是近年來廣泛應用的SNP標記,具有遺傳穩定、數量多、分布廣等特點,基于其開發的 9K、90K、660K等基因芯片廣泛應用于關聯分析、連鎖作圖及遺傳多樣性評價等研究[16-20]。中國農業科學院作物科學研究所與Affymetrix公司合作開發的Affymetrix 50K基因芯片(北京博奧晶典生物技術有限公司)集成了一些重要性狀的功能標記位點,可直接用于種質資源及育種材料的檢測評價。【本研究切入點】寧麥系列品種經過30年的發展,逐步形成了赤霉病抗性突出、弱筋品質優良的品種特色,在生產與研究中具有重要地位。雖然已對個別品種開展了較為深入的研究,但對大部分寧麥系列品種重要性狀的遺傳組成尚不明確。【擬解決的關鍵問題】本研究以寧麥系列23個已審定品種及51份高代品系為材料,利用中國農業科學院作物科學研究所與Affymetrix公司合作開發的50K基因芯片獲取基因型,同時結合功能基因鑒定,一方面對寧麥系列品種(系)的遺傳多樣性進行評價;另一方面系統分析寧麥系列品種(系)的重要性狀功能基因的組成,為寧麥系列品種的遺傳改良及育種利用提供理論依據。
1 材料與方法
1.1 試驗材料
23個已審定的寧麥系列品種和51份高代品系(圖1和電子附表1),均由江蘇省農業科學院麥類作物研究室保存。
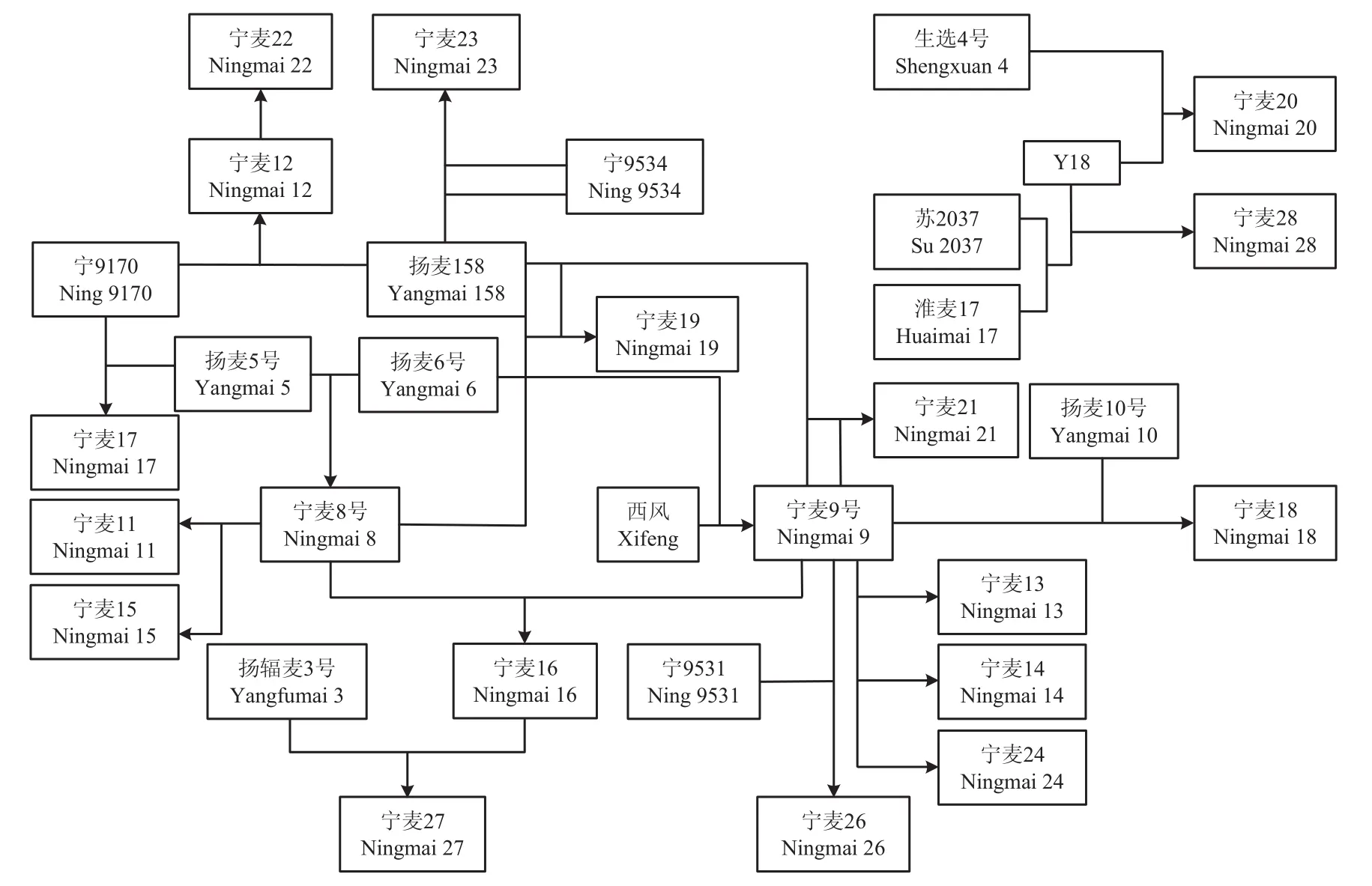
圖1 寧麥系列品種的系譜Fig.1 Pedigrees of Ningmai series wheat cultivars
對寧麥系列品種的主要性狀表現進行系統整理,數據主要來源于各品種的審定報告,產量性狀數據來源于生產試驗數據,同時通過省審定和國家審定的品種則采用省審定數據,品質數據采用2年區試結果平均值(表1)。中國品種審定制度于20世紀90年代確立[21],因此,早期品種寧麥3號與寧麥6號缺乏相關數據。
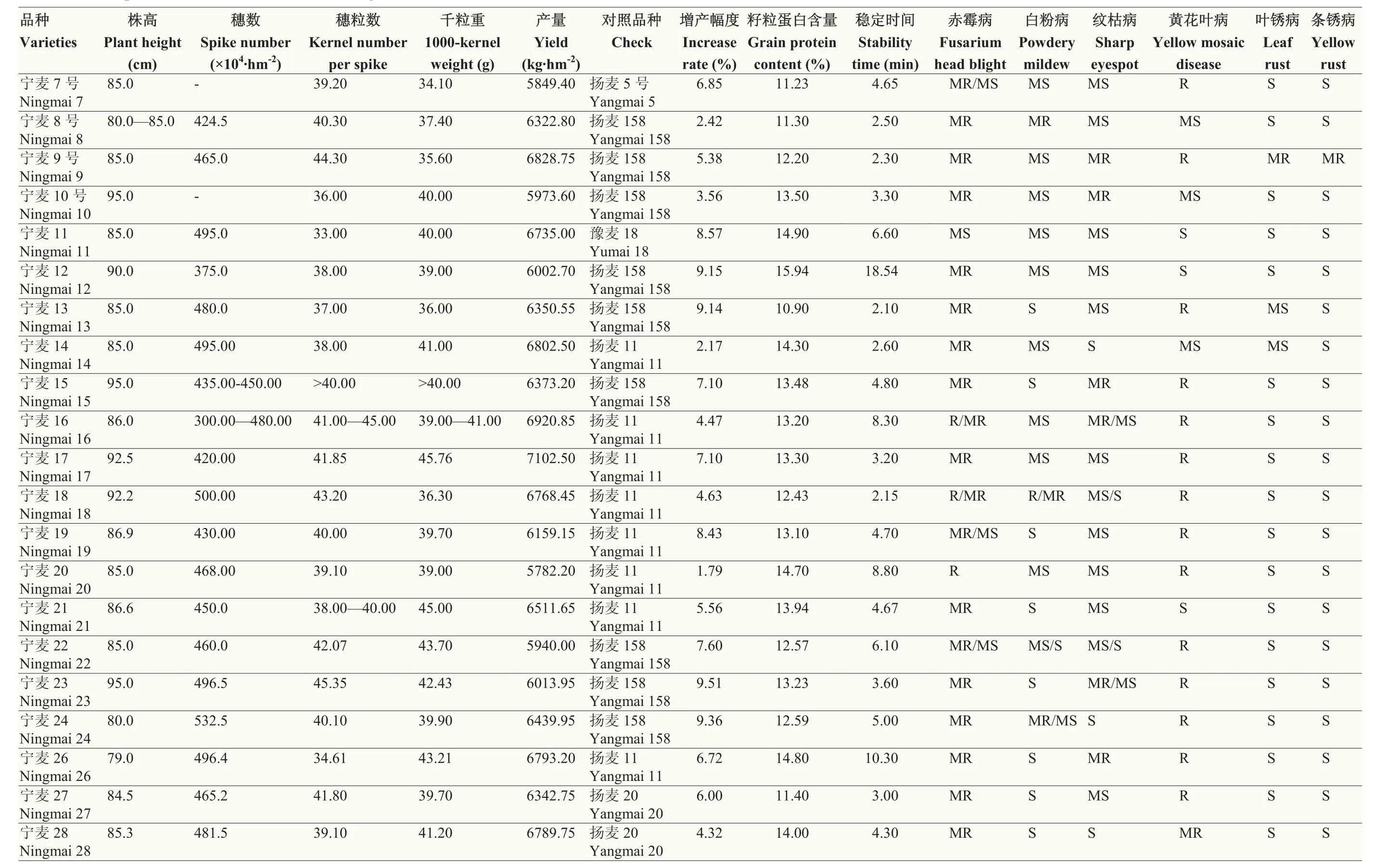
表1 寧麥系列品種的主要性狀表現Table 1 The performance of main traits in Ningmai series wheat cultivars
1.2 基因型分析
將74份試驗材料種子室溫萌發7 d左右,剪取幼嫩葉片,采用 CTAB法提取基因組 DNA[22]。利用中國農業科學院作物科學研究所與Affymetrix公司合作開發的小麥 50K SNP芯片對所有材料進行全基因組掃描,芯片包含部分重要性狀的功能基因[23],基因芯片測試服務由北京博奧晶典生物技術有限公司提供。此外,對芯片不包含的重要性狀功能基因進行了補充分子標記鑒定(表2與電子附表2)。
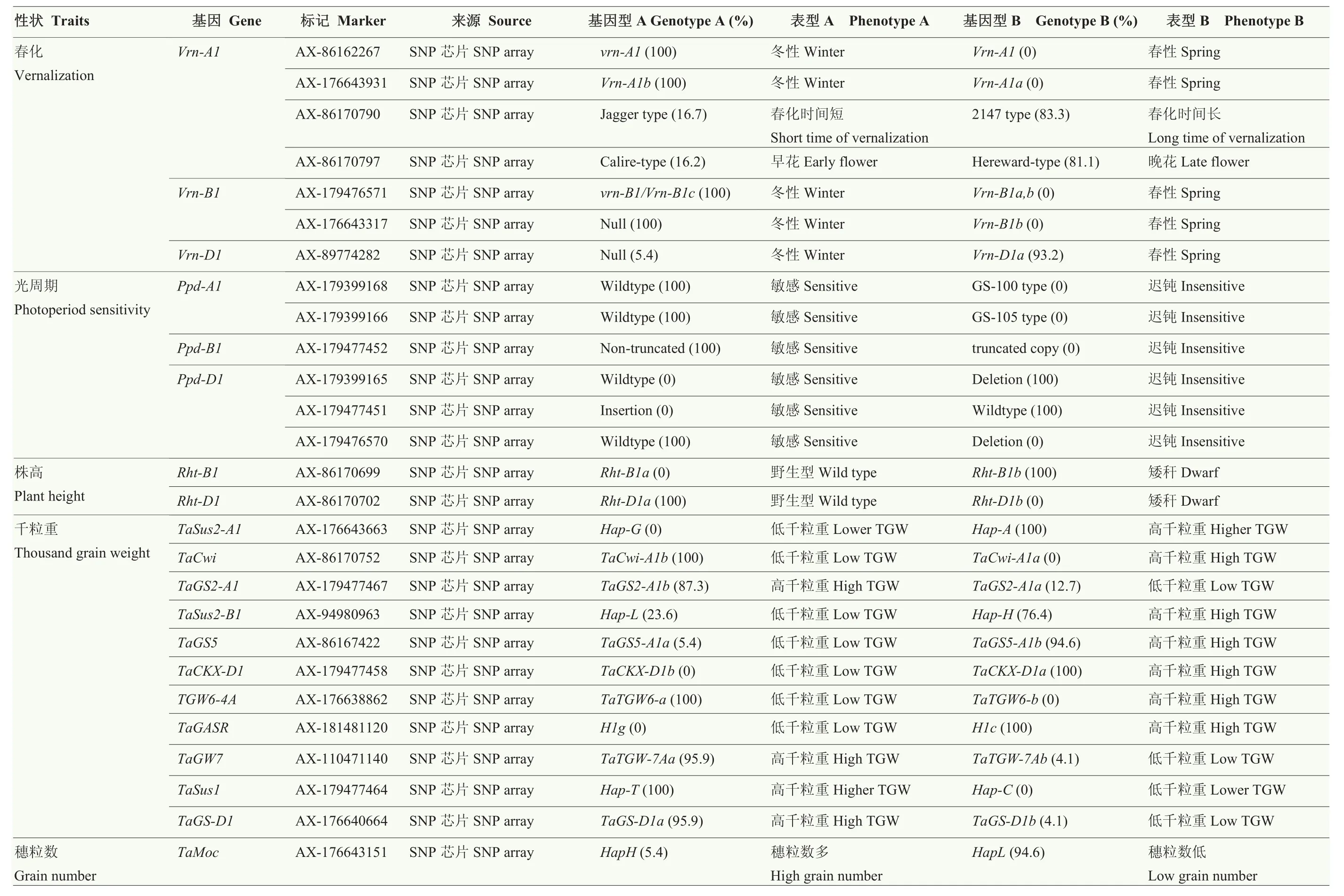
表2 寧麥系列品種(系)重要性狀相關位點的分布Table 2 Distribution of the loci relatedto important traits in Ningmai series varieties (lines)
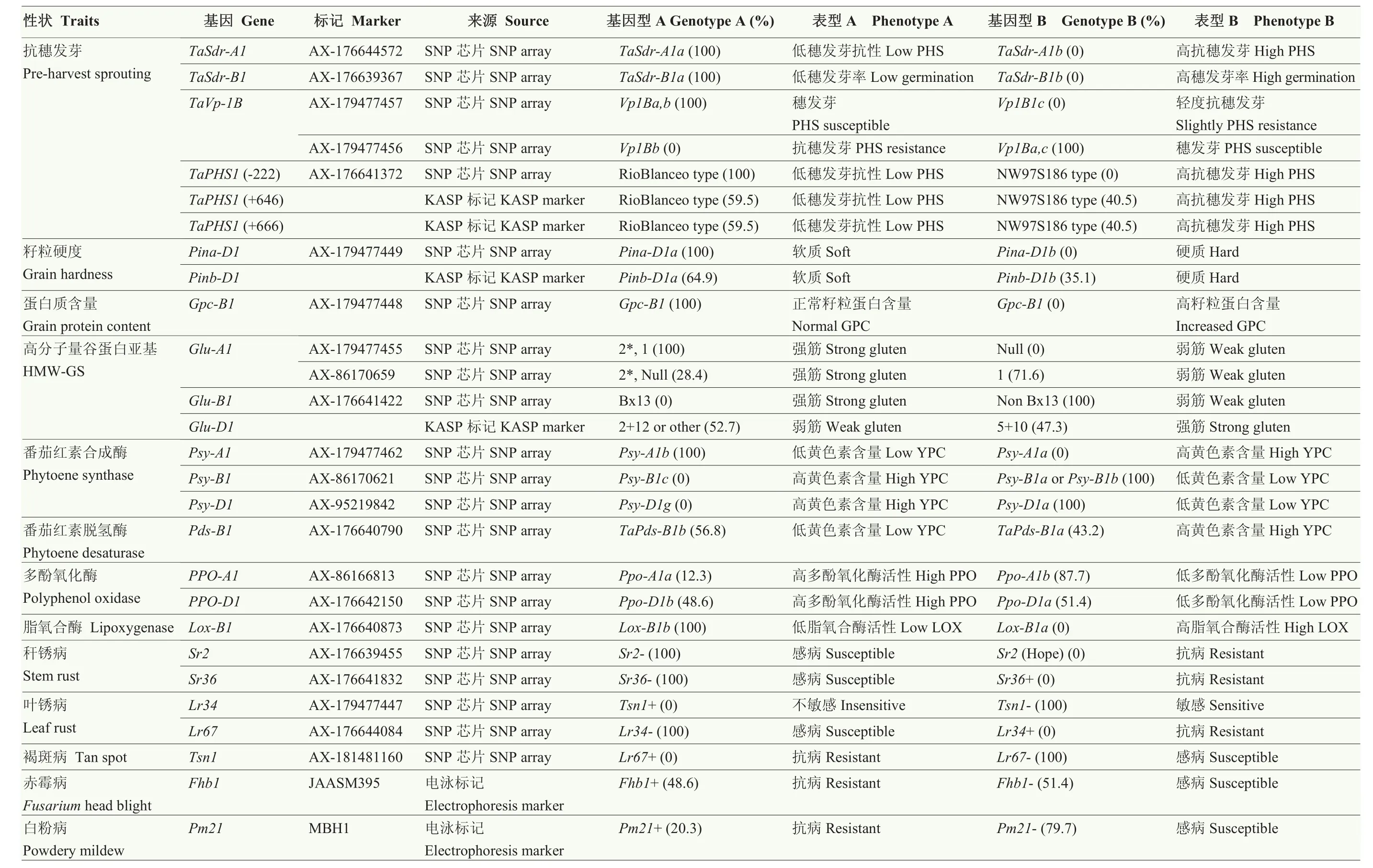
續表2 Continued table 2
根據RASHEED等[23]報道的KASP標記序列合成引物,每個標記設計2條SNP特異性引物(F1/F2)和1條通用引物(R),F1尾部添加能夠與FAM熒光結合的特異性序列(5′-GAAGGTGACCAAGTTC ATGCT-3′),F2尾部添加能夠與HEX熒光結合的特異性序列(5′-GAAGGTCGGAGTCAACGGATT-3′)。根據吳磊等[24]和 BIE等[25]研究合成抗赤霉病基因Fhb1的診斷標記 JAASM395與抗白粉病基因 Pm21的診斷標記MBH1設計引物。引物均由生工生物工程(上海)股份有限公司合成。
KASP反應總體系為5 μL,包含2×KASP Master Mix 2.5 μL、KASP Assay Mix(引物混合工作液)0.07 μL、濃度為 20 ng·μL-1的模板 DNA 2.43 μL。KASP 反應程序為 94℃ 15 min;94℃ 20 s,61—55℃1 min,每個循環降低0.6℃,共10個循環;94℃ 20 s,55℃ 1 min,共26個循環。通過KASP熒光分析儀(LGC公司型號為PHERAstar plus)掃描分析PCR結果。
電泳檢測標記的 PCR擴增體系為 10×buffer 1μL、25 mmol·L-1的MgCl20.5 μL、2.5 mmol·L-1的dNTP 0.5 μL、10 μmol·L-1的前后引物各 0.1 μL、5 U·μL-1的Taq 聚合酶 0.2 μL、50 ng·μL-1的 DNA 模板 3 μL,ddH2O 補至 10 μL。PCR 擴增程序為 94℃ 3 min;94°C 15 s,58°C 30 s,72°C 30 s,35 個循環;72°C 5 min。擴增產物用1.5%瓊脂糖凝膠電泳檢測。
1.3 數據分析
利用Microsoft Excel 2016對基因芯片數據進行初步處理,過濾掉缺失率>10%,最小基因頻率<5%的標記。參照 BOTSTEIN等[26]的方法計算多態性信息量(polymorphism information content,PIC),利用NTSYSpc version 2.10t軟件計算遺傳相似性系數,在Mega 6.0軟件中作Neighbor Joining聚類分析[27]。
2 結果
2.1 寧麥系列品種的主要性狀表現
寧麥系列品種的豐產性較為突出,在統計的 21個品種中,較對照增產超過5%的有14個,更有4個品種增產超過9%(表1);從產量三要素來看,不同品種的穗數、穗粒數及千粒重差異較大,千粒重隨著審定時間的推移呈上升趨勢,而穗數與穗粒數的變化趨勢不明顯。不同品種的株高也存在較大差異,寧麥24、寧麥26的株高約80.0 cm,而寧麥15、寧麥23等則達到95.0 cm,其余品種均在此范圍內。從籽粒蛋白含量與穩定時間來看,達到弱筋小麥標準的品種有5個,分別為寧麥8號、寧麥9號、寧麥13、寧麥18和寧麥 27,而達到強筋小麥標準的品種僅有寧麥 12與寧麥26。在抗病性方面,寧麥系列品種的赤霉病與黃花葉病抗性突出,赤霉病多數表現為中抗,寧麥20更是達到抗,寧麥9號、寧麥13、寧麥16等表現為抗黃花葉病;白粉病與紋枯病抗性一般,多數為中感,少數品種達到中抗水平;銹病抗性較差,葉銹病、條銹病及稈銹病均表現感病。
2.2 寧麥系列品種的遺傳分析
2.2.1 標記的多態性分析及其分布 經過多態性及缺失標記篩選,共獲得28 253個高質量SNP位點用于進一步遺傳分析,A、B、D染色體組分別包含8 973、10 532和8 748個標記,其中,3B染色體上標記最多,達到2 430個,4D染色體標記最少,為729個(圖2)。各位點多態性信息含量(PIC)值為0.10—0.67,差異較大,平均為0.36。

圖2 各染色體的標記分布及其多態性信息含量Fig.2 SNP distribution and the polymorphism information content on the chromosomes
2.2.2 寧麥系列品種的遺傳相似性及聚類分析 已審定的23個品種間的遺傳相似系數為0.407—0.964,平均為 0.600;51個高代品系間的遺傳相似系數為0.456—0.985,平均為0.684,呈現更高的遺傳相似性(圖3和電子附表3)。從高代品系與已育成的品種的遺傳相似系數看,各品系與寧麥13、寧麥14、寧麥24、寧麥26、寧麥27、寧麥28等品種的遺傳相似性較高,平均在0.650以上,而與寧麥11、寧麥15、寧麥16、寧麥17、寧麥23的遺傳相似性較低,平均在0.500以下。
品種系譜顯示,寧麥8號、寧麥9號與揚麥158等3個品種扮演了核心親本的角色,而寧麥8號的親本揚麥5號是揚麥158的姊妹系,結合聚類分析結果,可將育成品種分為2個類群(圖4-A),寧麥12、寧麥15、寧麥17、寧麥22、寧麥23等可與寧麥8號歸為一類,它們有的是從寧麥8號系統選擇而來,有的與寧麥8號擁有相同的親本;寧麥13、寧麥14、寧麥16、寧麥18、寧麥24、寧麥26等可與寧麥9號歸為一類,它們有的是從寧麥9號系統選擇而來,有的是寧麥9號與其他親本雜交育成,與寧麥8號的類群相比,其品種間遺傳距離較近。結合遺傳相似性及聚類分析發現新選育的高代品系與寧麥9號相似性更高,親緣關系更近(圖3和圖4-B)。
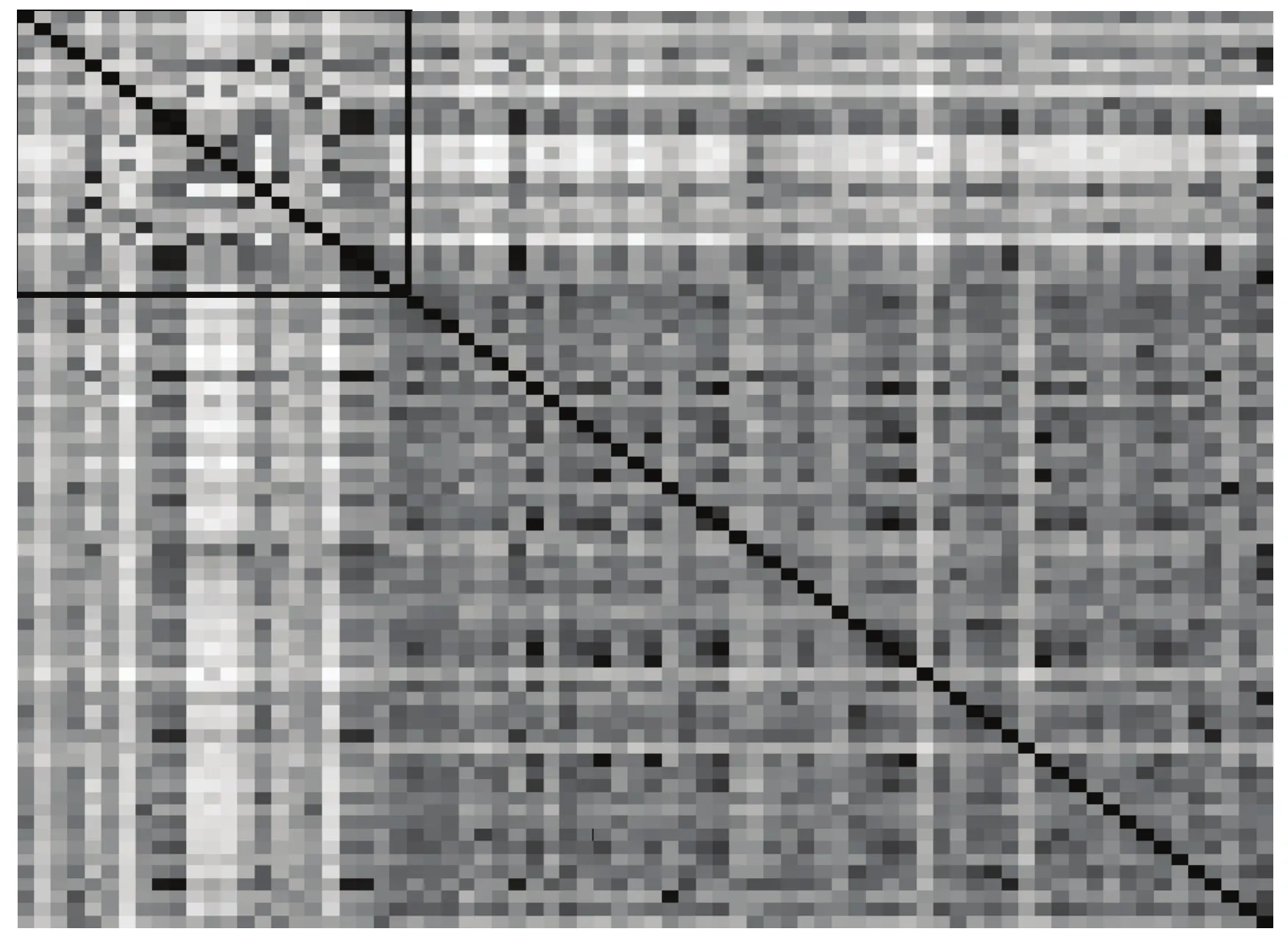
圖3 寧麥系列品種(系)的遺傳相似系數Fig.3 Genetic similarity coefficients among the Ningmai varieties (lines)
2.3 寧麥系列品種(系)重要性狀相關位點的分布
2.3.1 農藝性狀相關標記檢測 春化基因Vrn-A1在所有材料中均表現為Vrn-A1b冬性變異類型,通過對其另外 2個功能變異位點的檢測結果顯示,83.3%的材料需較長的春化時間,81.1%的材料表現為晚花類型;Vrn-B1均表現為冬性變異類型;93.2%的材料為Vrn-D1a春性變異類型。控制光周期反應的主要基因Ppd-D1在所有材料中均為光周期遲鈍類型,而其同源位點Ppd-A1與Ppd-B1則均為光周期敏感類型。株高位點的檢測結果顯示所有材料均為 Rht-B1b/Rht-D1a變異類型。
2.3.2 產量相關標記檢測 千粒重調控位點 TaGS2-A1、TaGS5、TaGS-D1、TaGASR、TaSus1、TaSus2、TaCKX6-D1及TaGW7等在多數材料中(76.4%—100.0%)均表現為增加千粒重的等位變異,TaCwi-A1與TaTGW6在所有材料中表現為降低千粒重的等位變異,僅有 4份材料攜帶穗粒數控制位點 TaMOC1-7A的優勢等位變異。
2.3.3 抗穗發芽相關標記檢測 寧麥系列小麥品種多為紅皮小麥,抗穗發芽,分子標記檢測結果顯示抗穗發芽基因 TaSdr-A1在所有材料中均表現為TaSdr-A1a不抗穗發芽類型,而其同源基因TaSdr-B1則均為TaSdr-B1a抗穗發芽類型;TaVp-1B均呈現不抗穗發芽的Vp1Ba變異類型;TaPHS1具有多個變異位點,-222變異位點處均為不抗穗發芽的等位變異,+646和+666處功能變異位點在檢測材料中呈現 GT和AA 2種單倍型,其中40.5%的材料為抗穗發芽的單倍型(AA)。
2.3.4 品質性狀相關標記檢測 寧麥系列品種的硬度控制位點Pina-D1均為Pina-D1a軟麥類型,Pinb-D1位點在不同材料存在差異,35.1%的材料為 Pina-D1b變異類型。蛋白質含量控制位點 Gpc-B1均表現為正常水平蛋白質含量的等位變異類型。高分子量谷蛋白亞基Glu-A1位點處有71.6%的材料為1類型;Glu-B1位點處均不攜帶Bx13亞基;在Glu-D1位點處,47.3%的材料攜帶5+10亞基,52.7%的材料為2+12或其他亞基類型。與黃色素含量(yellow pigment content,YPC)相關的基因中,番茄紅素合成酶基因的3個同源位點 Psy-A1、Psy-B1與 Psy-D1分別為 Psy-A1b、non-Psy-B1c與Psy-D1a類型,均為降低黃色素含量的等位變異;43.2%的材料在番茄紅素脫氫酶 TaPds-B1位點處呈現TaPds-B1a增加黃色素含量的等位變異。在多酚氧化酶(polyphenol oxidase,PPO)活性控制位點上,87.7%的材料為低活性的 PPO-A1b類型,51.4%的材料為低活性的 PPO-D1a類型。脂氧合酶(lipoxygenase,LOX)活性控制位點 Lox-B1均為低活性的Lox-B1b類型。
2.3.5 抗病性相關標記檢測 稈銹病、葉銹病及褐斑病均不是長江中下游麥區的常發病害,抗稈銹病基因(Sr2、Sr36)、抗葉銹病基因(Lr34和 Lr67)及抗褐斑病基因(Tsn1)在所有材料中均未發現。抗赤霉病主效基因 Fhb1在檢測材料中的分布頻率達到48.6%,是寧麥系列小麥品種赤霉病抗性的重要來源。寧麥系列品種在白粉病抗性方面表現一般,已審定品種均不攜帶抗白粉病基因 Pm21,而在高代品系中這一情況有所改善,分布頻率達到近30%。
3 討論
3.1 寧麥系列品種的性狀特點與遺傳多樣性
寧麥系列品種立足長江中下游麥區的氣候條件及生產特點,培育了寧麥9號、寧麥13、寧麥18等優質弱筋品種,寧麥14、寧麥16、寧麥24等中筋品種,以及中強筋品種寧麥 26,突出的赤霉病抗性與良好的豐產性是寧麥系列品種的重要特色,為保障糧食安全、農業增效及農民增收作出了重要貢獻。近年來,小麥市場受國際市場和國內需求變化的影響,弱筋粉較多地以進口淀粉加中強筋面粉混合替代,使得淮南麥區的中強筋、強筋小麥新品種越來越受到市場的青睞[28-29],受極端氣候影響,小麥白粉病、銹病等的發病范圍與發病程度呈上升趨勢[30],嚴重威脅小麥生產,給小麥育種帶來了新的挑戰。
中國現代品種的遺傳多樣性隨著育種歷程呈現逐漸下降的趨勢[31],寧麥系列品種亦呈現這一特點,已審定品種的遺傳相似系數平均為0.60,而新選育的高代品系的遺傳相似系數平均達到0.68。育種過程中包含強烈的人工選擇,一些承受強選擇作用的基因在群體中的多樣性顯著降低,同時,這些基因附近區域由于牽連效應遺傳多樣性也明顯下降[32]。從系譜來看,2000年之后育成的品種多含有寧麥8號和寧麥9號的遺傳背景,并且這兩個品種也是本區域近年來小麥生產中廣泛采用的親本。其中,寧麥8號具有高產、半矮稈抗倒、大穗、多粒的優點,寧麥9號則多穗、多粒、品質優良且抗病性突出。兩個品種優良性狀具有一定的互補性,利用二者配組選育出寧麥16、生選6號、揚輻麥4號等品種。隨著寧麥9號的系選品種寧麥13在生產上取得巨大成功,寧麥9號的衍生品種更是逐漸增多,如寧麥 14、寧麥 24、寧麥 26、生選6號、揚麥18、揚麥21、南農0686、鎮麥8號、蘇麥8號、農麥126等。寧麥9號與3個系選品種寧麥13、寧麥14及寧麥24的遺傳相似系數在0.7左右,而這3個品種間的遺傳相似系數均超過 0.9,推斷在寧麥 9號剩余變異群體中的選擇過程中可能出現一次較大的基因重組或突變事件,保留了寧麥9號的諸多優良性狀,如矮稈、多穗、多粒等,同時抗倒性、千粒重等性狀得到改善,使品種的豐產性、穩產性大大提高。
株高是育種選擇的重要指標,寧麥9號類群品種的株高顯著低于寧麥8號類群,可能使寧麥9號類群的后代大量保留,進而使品種同質化程度升高。由此可見,寧麥系列品種急需拓寬遺傳基礎,實現品種升級。通過遠緣雜交[33]或盡可能避免遺傳背景過近品種(系)間雜交[34],有助于豐富品種的遺傳多樣性,培育出突破性的新品種。
3.2 寧麥系列品種(系)重要性狀相關位點的分布
寧麥系列品種(系)幾乎都為春性品種, Vrn-D1位點的變異可能是多數材料表現春性的主要原因;多數材料為光周期遲鈍型,主要由光周期反應基因Ppd-D1決定;當前在育種中主要利用了Rht-B1位點的變異來降低株高,未來可嘗試引入Rht-D1位點的變異。
產量要素中,千粒重的遺傳力最高,其相關遺傳位點報道最多,一些重要的千粒重調控位點的優勢等位變異已經在現有材料中固定下來,如 TaGS2-A1、TaGS5、TaGS-D1等;有的優勢等位變異在寧麥系列品種(系)中未檢測到,可能是受自然選擇與人工選擇影響,前人研究證實TaCwi-A1a在南方麥區材料中分布頻率很低[35],TaTGW6-b在現代小麥品種很少檢測到[36],今后可考慮引入這些優勢等位變異以進一步提高千粒重。穗粒數相關位點 TaMOC1-7A的優勢類型HapH在長江中下游麥區材料中有一定的分布,寧麥9號含有這一優勢等位變異,可能是其多粒特性的變異來源之一,但在目前寧麥系列品種(系)中分布較少,今后可加強利用。
寧麥系列品種的品質類型較為豐富,存在弱筋、中筋和中強筋品質類型,并以中筋、弱筋類型為主。分子標記檢測結果顯示,Pina-D1a/Pinb-D1a基因型比例達到 64.9%,因此,寧麥系列品種(系)多為軟質麥。高分子量谷蛋白亞基的組成類型對面筋強度具有較大影響[37],寧麥系列品種的 Glu-A1、Glu-B1和Glu-D1位點上低面筋強度的亞基類型占有較大比例,與其中筋、弱筋品質類型為主的表型特征是相符的;提升面筋強度的 5+10亞基類型試驗材料中也有一定分布,今后需根據不同的育種目標分別加強對高分子量谷蛋白亞基類型的選擇。YPC、PPO及 LOX活性均對小麥面制品的外觀品質有重要影響[38],特別是YPC更是與營養品質相關;中國傳統主食對面粉白度要求較高,因此多數品種的YPC、PPO及LOX活性均處于較低水平,而隨著人們營養和保健意識的提高,亮黃色的面粉和面制品越來越受到重視[39]。寧麥系列品種(系)主要面向傳統市場,在營養品質研究方面存在不足。
小麥銹病近年來呈波動性持續上升趨勢[30],本研究與前人研究[40-41]均表明寧麥系列品種極少攜帶抗銹病基因,今后應加強相關基因的利用,提高品種銹病抗性水平。寧麥系列品種(系)的赤霉病抗性突出,常被用作抗性親本[42],寧麥9號還被證實為中國小麥品種中主效抗病基因Fhb1的主要來源[43]。Fhb1在寧麥系列品種(系)中有較高頻率的分布,遠高于其他區域或單位的材料[44-46]。低水平的白粉病抗性影響了寧麥系列品種在生產中的推廣應用[47],近些年,寧麥系列品種的育種者十分重視白粉病抗性的選擇,特別是加強抗性親本的應用,使白粉病抗性得到較大改善,已有近 30%的高代品系含有主效抗病基因 Pm21。寧麥系列品種(系)主要攜帶TaSdr-B1與TaVp-1B抗穗發芽基因,TaPHS1的+646和+666功能變異位點在材料中呈現一定分化,總體來說寧麥系列品種(系)穗發芽抗性表現較好,可作為優異親本使用。
3.3 分子育種體系的建立與應用
在分子育種技術應用方面,寧麥系列品種早期主要針對赤霉病抗性開展分子標記輔助選擇,這也是抗病基因Fhb1分布頻率較高的主要原因。近年來,本團隊一直致力于構建系統的分子育種技術體系,一方面利用現有主效抗病位點,包括抗白粉病基因Pm21、抗條銹病基因Yr26等;另一方面是產量、品質等數量性狀相關位點聚合選擇體系的建立,包括千粒重、籽粒蛋白含量等性狀,目前,已篩選出的多個效應顯著的千粒重控制位點[3],籽粒蛋白含量控制位點的效應評價工作也已完成[48]。此外,還期望通過構建高通量 KASP分子標記選擇體系及多重PCR選擇體系來應對不同的分析場景,前者主要應用于大規模育種群體的分析,如前人報道的Fhb1的KASP標記[49],自主開發的Qfhb-5A、Qtkw-1B等[3,14];后者則是小群體的快速檢測,目前,已經成功開發Fhb1與Pm21的多重PCR標記[50]。通過以上分子育種體系的構建與應用,期望提高目標性狀選擇效率,加速育種進程。
4 結論
寧麥系列品種(系)具有突出的豐產性、中弱筋品質及赤霉病抗性,包含較多提升千粒重、抗穗發芽及抗病性的優勢位點,可作為優良親本應用于雜交配組;寧麥系列品種(系)的遺傳多樣性呈現降低趨勢,有必要加強種質創新,拓寬育種群體遺傳背景,同時加強白粉病、銹病的抗性改良及中強筋、強筋品質類型的品種選育。
