冬小麥擴(kuò)展蛋白TaEXPA8黑曲霉工程菌的構(gòu)建及纖維素水解作用分析
2022-02-22 11:35:34王雪馮旭賀付蒙徐永清袁強(qiáng)李麗劉丹孔德興李鳳蘭
食品與發(fā)酵工業(yè) 2022年2期
關(guān)鍵詞:效率
王雪,馮旭,賀付蒙,徐永清,袁強(qiáng),李麗,劉丹,孔德興,李鳳蘭
(東北農(nóng)業(yè)大學(xué) 生命科學(xué)學(xué)院,黑龍江 哈爾濱,150030)
秸稈是農(nóng)業(yè)生產(chǎn)中的副產(chǎn)物,我國(guó)的秸稈總量巨大,但綜合利用效率極低[1]。焚燒是處理秸稈的主要方式,但該方法不僅造成資源浪費(fèi),同時(shí)導(dǎo)致大氣環(huán)境的嚴(yán)重污染[2-3]。秸稈中富含纖維素,通過(guò)降解纖維素使其轉(zhuǎn)化為新型能源能夠極大的緩解現(xiàn)階段的能源短缺和環(huán)境污染問(wèn)題,但降解效率過(guò)低仍是現(xiàn)階段限制該項(xiàng)產(chǎn)業(yè)發(fā)展的瓶頸之一[4]。纖維素具有微纖維組成的復(fù)雜、多層結(jié)構(gòu),這種結(jié)構(gòu)使纖維素具有較強(qiáng)的硬度且極難溶于水,纖維素酶也因不能進(jìn)入微纖維中纖維鏈的深處從而導(dǎo)致酶解效率較低[5]。因此,探索高效、簡(jiǎn)便的纖維素降解方法具有重要的實(shí)際價(jià)值。
現(xiàn)階段輔助纖維素降解的方式通常有物理法、化學(xué)法、物理化學(xué)法及生物法[6]。在以上幾種方法中,前3種通常需要高溫或高壓,成本較高,而生物法通常在常溫下即能完成,成本低且無(wú)污染[7]。纖維素的水解是一個(gè)復(fù)雜的過(guò)程,自然界中能夠降解纖維素的生物通常具有包含纖維素酶在內(nèi)的多酶體系,包括內(nèi)切葡聚糖酶、纖維二糖酶、β-葡萄糖苷酶等,這些酶能夠發(fā)揮協(xié)同作用,將纖維素降解為可溶性多糖[8]。然而,生物法降解纖維素所需的時(shí)間仍然較長(zhǎng),且效率不高。因此,迫切需要高效降解纖維素的微生物或者提高纖維素酶效率的添加劑[9]。
擴(kuò)展蛋白以非酶活性的作用方式發(fā)揮功能,即在不改變細(xì)胞壁共價(jià)結(jié)構(gòu)的條件下,只通過(guò)破壞基質(zhì)多糖與纖維素微纖絲之間氫鍵的方式直接作用于細(xì)胞壁[10]。近些年的研究表明,擴(kuò)展蛋白能夠以協(xié)同蛋白的方式提高纖維素酶的水解效率,尤其是來(lái)自微生物中的擴(kuò)展蛋白[11-12]。來(lái)源于B.subtlis的擴(kuò)展蛋白BsEXLX1與纖維素酶混合協(xié)同作用于纖維素底物,其活力相比于單一纖維素酶提高了2~5.7倍[13]。來(lái)源于真菌A.fumigatus的擴(kuò)展蛋白AfSwol與內(nèi)切纖維素酶協(xié)同作用于底物羧甲基纖維素72 h后,水解活力提高了1.2倍[14]。因此,尋找高效、合適的擴(kuò)展蛋白對(duì)于提高纖維素降解效率和降低生產(chǎn)成本是非常必需的。
擴(kuò)展蛋白在天然生物體中的表達(dá)量較低,需要體外高效的表達(dá)系統(tǒng)以滿(mǎn)足生產(chǎn)需求。大腸桿菌通常用于外源蛋白的表達(dá),然而植物及真菌的擴(kuò)展蛋白在大腸桿菌中表達(dá)時(shí),重組蛋白會(huì)發(fā)生變性形成包涵體從而失活[15-16]。因此,本研究選擇了黑曲霉(Aspergillusniger)表達(dá)系統(tǒng)以生產(chǎn)冬小麥(Triticumaestivum)擴(kuò)展蛋白TaEXPA8,同時(shí)對(duì)該蛋白在纖維素水解中的作用進(jìn)行了初步探索,為植物擴(kuò)展蛋白在纖維素水解中的應(yīng)用提供理論基礎(chǔ)及新的方法和思路。
1 材料與方法
1.1 植物材料及菌株
冬小麥“東農(nóng)冬麥1號(hào)”(Dongnongdongmai-1,DN-1)由東北農(nóng)業(yè)大學(xué)植物資源與分子生物學(xué)實(shí)驗(yàn)室提供,大腸桿菌(Escherichiacoli)Trans-T1感受態(tài)、根癌農(nóng)桿菌(Agrobacteriumtumefaciens)AGL1感受態(tài)購(gòu)于北京全式金生物科技有限公司,黑曲霉CICC2462由東北農(nóng)業(yè)大學(xué)遺傳學(xué)實(shí)驗(yàn)室提供。
1.2 試劑盒、酶及表達(dá)載體
RNA、DNA提取試劑盒(Easy Pure?)、反轉(zhuǎn)錄試劑盒(Easy Script?)、高保真PCR酶(Trans Fast?)、pEASY-T3克隆載體,北京全式金生物科技有限公司;SDS-PAGE試劑盒,Thermo Fisher公司;限制性?xún)?nèi)切酶ApaΙ和Hind Ⅲ、T4-DNA連接酶,NEB公司;pSZHGS表達(dá)載體由東北農(nóng)業(yè)大學(xué)遺傳學(xué)實(shí)驗(yàn)室提供。
1.3 擴(kuò)展蛋白TaEXPA8基因的克隆
采用試劑盒對(duì)東農(nóng)冬麥1號(hào)葉片中的總RNA進(jìn)行提取,1.2%(質(zhì)量分?jǐn)?shù))瓊脂糖凝膠電泳檢測(cè)RNA質(zhì)量并測(cè)定濃度,利用反轉(zhuǎn)錄試劑盒獲得cDNA,方法詳見(jiàn)說(shuō)明書(shū)。通過(guò)PCR法獲得TaEXPA8基因的全長(zhǎng),本試驗(yàn)所用引物如表1所示。PCR產(chǎn)物經(jīng)純化后連接T3克隆載體并轉(zhuǎn)化至大腸桿菌中用于測(cè)序,同時(shí)對(duì)結(jié)果進(jìn)行NCBI在線(xiàn)Blast分析。
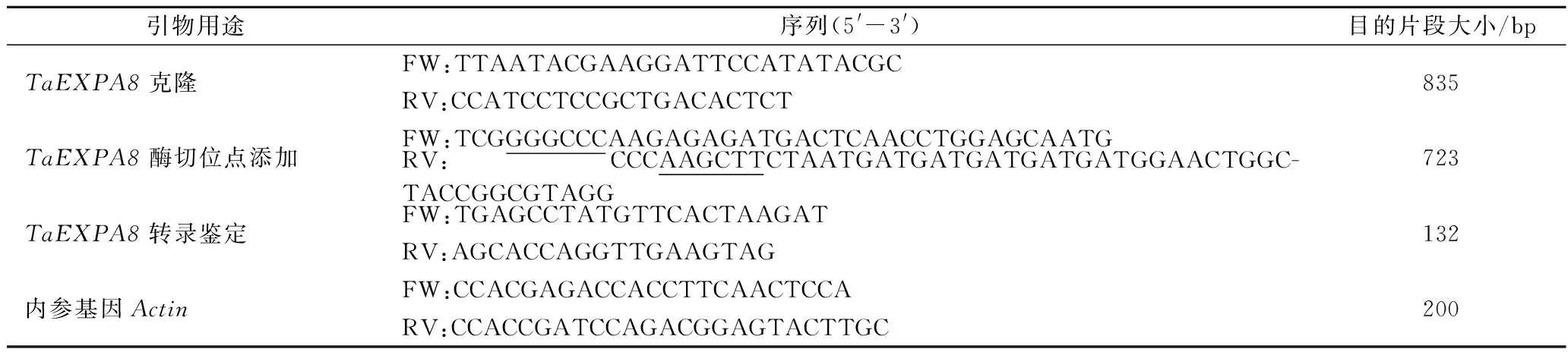
表1 試驗(yàn)所用引物列表Table 1 List of primers used in the experiment
1.4 pSZHGS-TaEXPA8表達(dá)載體的構(gòu)建
通過(guò)PCR法在TaEXPA8基因的上下游分別添加ApaΙ和Hind Ⅲ酶切位點(diǎn)并切除信號(hào)肽,產(chǎn)物經(jīng)純化后備用。利用限制性?xún)?nèi)切酶ApaΙ和Hind Ⅲ分別對(duì)上述PCR產(chǎn)物和pSZHGS表達(dá)載體進(jìn)行雙酶切,酶切產(chǎn)物經(jīng)純化后通過(guò)T4-DNA連接酶進(jìn)行連接,連接產(chǎn)物轉(zhuǎn)化至大腸桿菌中用于測(cè)序。對(duì)測(cè)序正確的陽(yáng)性菌株進(jìn)行擴(kuò)大培養(yǎng),提取質(zhì)粒進(jìn)行雙酶切鑒定。
1.5 表達(dá)載體轉(zhuǎn)化黑曲霉
根癌農(nóng)桿菌介導(dǎo)的黑曲霉遺傳轉(zhuǎn)化方法參照李杰等[17]進(jìn)行,具體方法如下。將pSZHGS-TaEXPA8表達(dá)載體轉(zhuǎn)化至根癌農(nóng)桿菌AGL1中,方法詳見(jiàn)說(shuō)明書(shū),陽(yáng)性農(nóng)桿菌經(jīng)PCR鑒定后進(jìn)行活化培養(yǎng)。分別取二次活化的農(nóng)桿菌菌液1.5 mL和適量的黑曲霉菌絲置于無(wú)菌EP管中,混合后涂布于鋪有玻璃紙的固體馬鈴薯葡萄糖瓊脂培養(yǎng)基(potato dextrose agar,PDA)(含200 μmol/L的乙酰丁香酮),28 ℃共培養(yǎng)2 d 后進(jìn)行轉(zhuǎn)膜培養(yǎng),潮霉素及頭孢噻肟鈉為抗性篩選,提取陽(yáng)性菌落的DNA并進(jìn)行PCR鑒定。挑選鑒定結(jié)果正確的菌落于馬鈴薯葡萄糖水培養(yǎng)基中進(jìn)行擴(kuò)培,按照10%的接種量接種于發(fā)酵培養(yǎng)基,30 ℃,200 r/min培養(yǎng)6 d后取發(fā)酵液于50 mL離心管中,離心后取沉淀進(jìn)行RT-PCR鑒定以檢測(cè)TaEXPA8基因的轉(zhuǎn)錄水平。
1.6 TaEXPA8蛋白的SDS-PAGE檢測(cè)
取少量TaEXPA8黑曲霉工程菌的發(fā)酵上清液進(jìn)行SDS-PAGE試驗(yàn)以檢測(cè)TaEXPA8蛋白的表達(dá)情況,操作方法詳見(jiàn)試劑盒說(shuō)明書(shū)。
1.7 TaEXPA8工程菌的纖維素水解作用分析
采用濾紙崩解試驗(yàn)檢測(cè)TaEXPA8工程菌的纖維素水解作用,在纖維素酶液中分別添加等體積的TaEXPA8工程菌、野生型黑曲霉菌株的發(fā)酵上清液,以培養(yǎng)基為空白對(duì)照。取10 mL上述液體分別添加到鋪有50 mg無(wú)淀粉濾紙的培養(yǎng)皿中,25 ℃,2 d后取出濾紙于顯微鏡下觀察,同時(shí)采用3,5-二硝基水楊酸(3,5-dinitrosalicylic acid,DNS)法測(cè)定體系中的葡萄糖含量[18]。
1.8 數(shù)據(jù)處理
GraphPad Prism 5.0軟件用于差異顯著性分析及作圖,所有數(shù)值均以平均值±標(biāo)準(zhǔn)差表示。
2 結(jié)果與分析
2.1 冬小麥總RNA的提取
東農(nóng)冬麥1號(hào)植株葉片總RNA的電泳結(jié)果如圖1所示,28S、18S和5S條帶清晰,無(wú)降解現(xiàn)象,濃度的測(cè)定結(jié)果為(540±17)ng/μL,OD260/OD280為2.0±0.1。結(jié)果顯示RNA的純度較高,濃度適宜,可以用于下一步試驗(yàn)。
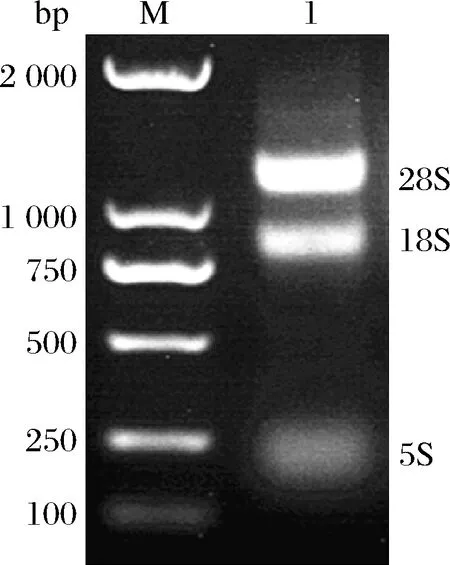
M-maker DL 2 000;1-RNA圖1 東農(nóng)冬麥1號(hào)葉片總RNA電泳結(jié)果Fig.1 Results of total RNA electrophoresis from DN-1 leaves
2.2 TaEXPA8基因的克隆
以RNA反轉(zhuǎn)錄的cDNA為模板對(duì)TaEXPA8基因進(jìn)行克隆,電泳結(jié)果(圖2)顯示目的條帶大小在750~1 000 bp,與引物設(shè)計(jì)時(shí)預(yù)期的條帶大小一致,可以進(jìn)行測(cè)序。將PCR產(chǎn)物純化后連接至T3克隆載體并轉(zhuǎn)化大腸桿菌進(jìn)行測(cè)序。NCBI的在線(xiàn)Blast分析結(jié)果(圖3)表明所獲得PCR產(chǎn)物與TaEXPA8基因的序列吻合度為99%,即已經(jīng)成功從東農(nóng)冬麥1號(hào)中獲得了TaEXPA8基因,可以進(jìn)行下一步試驗(yàn)。
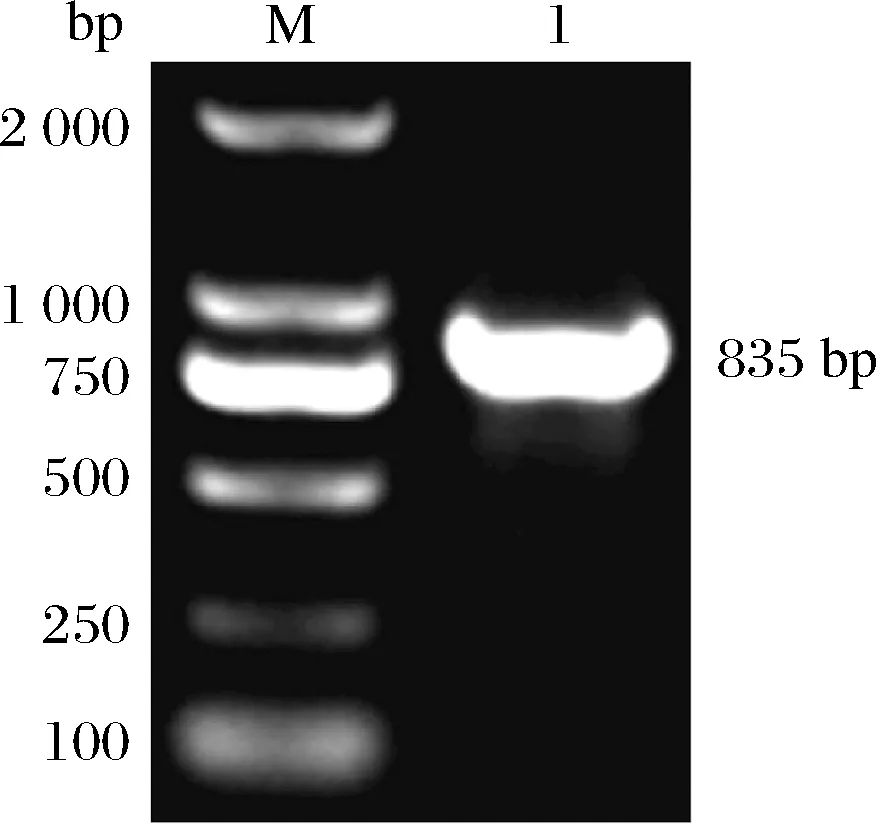
M-maker DL 2 000;1-TaEXPA8基因克隆結(jié)果圖2 TaEXPA8基因的克隆結(jié)果Fig.2 Cloning results of TaEXPA8 gene

圖3 TaEXPA8基因測(cè)序結(jié)果的NCBI在線(xiàn)Blast分析Fig.3 NCBI online blast analysis of TaEXPA8 gene sequencing results
2.3 pSZHGS-TaEXPA8表達(dá)載體的構(gòu)建
通過(guò)PCR在TaEXPA8基因的上下游分別添加ApaΙ和Hind Ⅲ酶切位點(diǎn)并切除信號(hào)肽。分別對(duì)PCR產(chǎn)物和pSZHGS表達(dá)載體進(jìn)行雙酶切,酶切產(chǎn)物連接后轉(zhuǎn)化至大腸桿菌。對(duì)構(gòu)建好的pSZHGS-TaEXPA8表達(dá)載體進(jìn)行雙酶切鑒定,結(jié)果(圖4)表明TaEXPA8基因已經(jīng)成功插入pSZHGS表達(dá)載體的ApaΙ和Hind Ⅲ酶切位點(diǎn)之間。
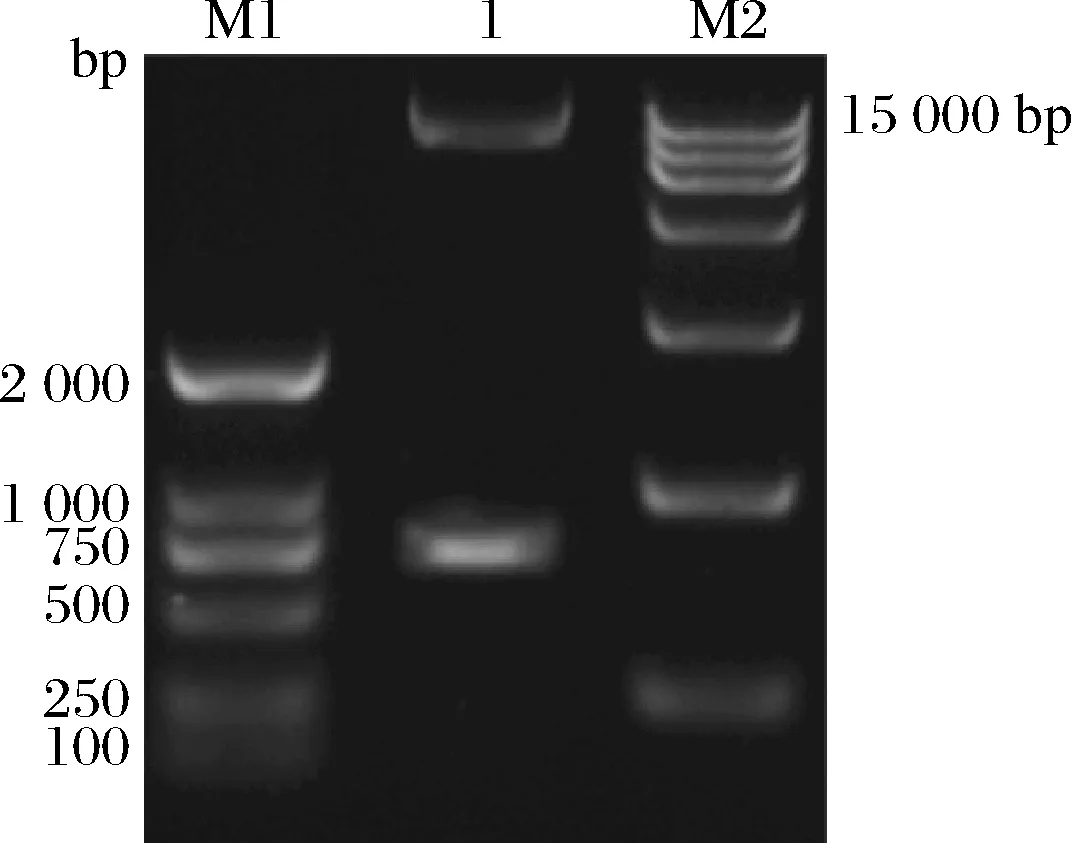
M1-maker DL 2 000;1-pSZHGS-TaEXPA8表達(dá)載體的雙酶切結(jié)果;M2-maker DL 15 000圖4 pSZHGS-TaEXPA8表達(dá)載體的雙酶切結(jié)果Fig.4 Results of double digestion of pSZHGS-TaEXPA8 expression vector
2.4 轉(zhuǎn)化pSZHGS-TaEXPA8表達(dá)載體黑曲霉菌株的獲得
將pSZHGS-TaEXPA8表達(dá)載體轉(zhuǎn)化至根癌農(nóng)桿菌AGL1中,活化后的農(nóng)桿菌菌液與黑曲霉共培養(yǎng)(圖5),并通過(guò)PCR分別在DNA和mRNA水平上對(duì)工程菌中的TaEXPA8基因進(jìn)行檢測(cè),結(jié)果見(jiàn)圖6。PCR的結(jié)果顯示已經(jīng)在黑曲霉工程菌的DNA中檢測(cè)到TaEXPA8基因,RT-PCR的鑒定結(jié)果表明該擴(kuò)展蛋白基因在工程菌中能夠正常的進(jìn)行轉(zhuǎn)錄。
圖5 轉(zhuǎn)化pSZHGS-TaEXPA8表達(dá)載體的農(nóng)桿菌與黑曲霉共培養(yǎng)結(jié)果Fig.5 Co-culture of Agrobacterium transformed pSZHGS-TaEXPA8 expression vector with Aspergillus niger
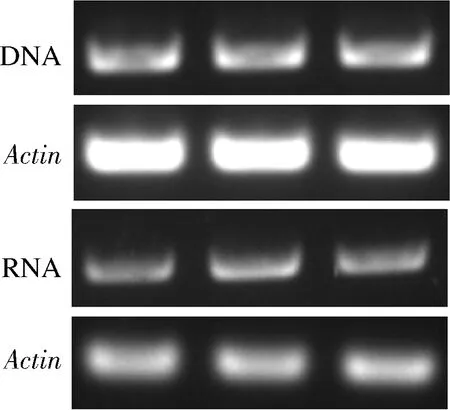
圖6 TaEXPA8黑曲霉工程菌的鑒定結(jié)果Fig.6 Identification results of TaEXPA8 engineered Aspergillus niger
2.5 TaEXPA8蛋白的SDS-PAGE檢測(cè)
通過(guò)SDS-PAGE檢測(cè)黑曲霉工程菌發(fā)酵上清液中TaEXPA8蛋白的表達(dá)情況,結(jié)果如圖7所示。TaEXPA8蛋白的分子質(zhì)量約為25 kDa,相比于野生型菌株,工程菌在相應(yīng)的位置處有條帶出現(xiàn),但不明顯。由此可見(jiàn)TaEXPA8蛋白已經(jīng)表達(dá),但表達(dá)量較低。
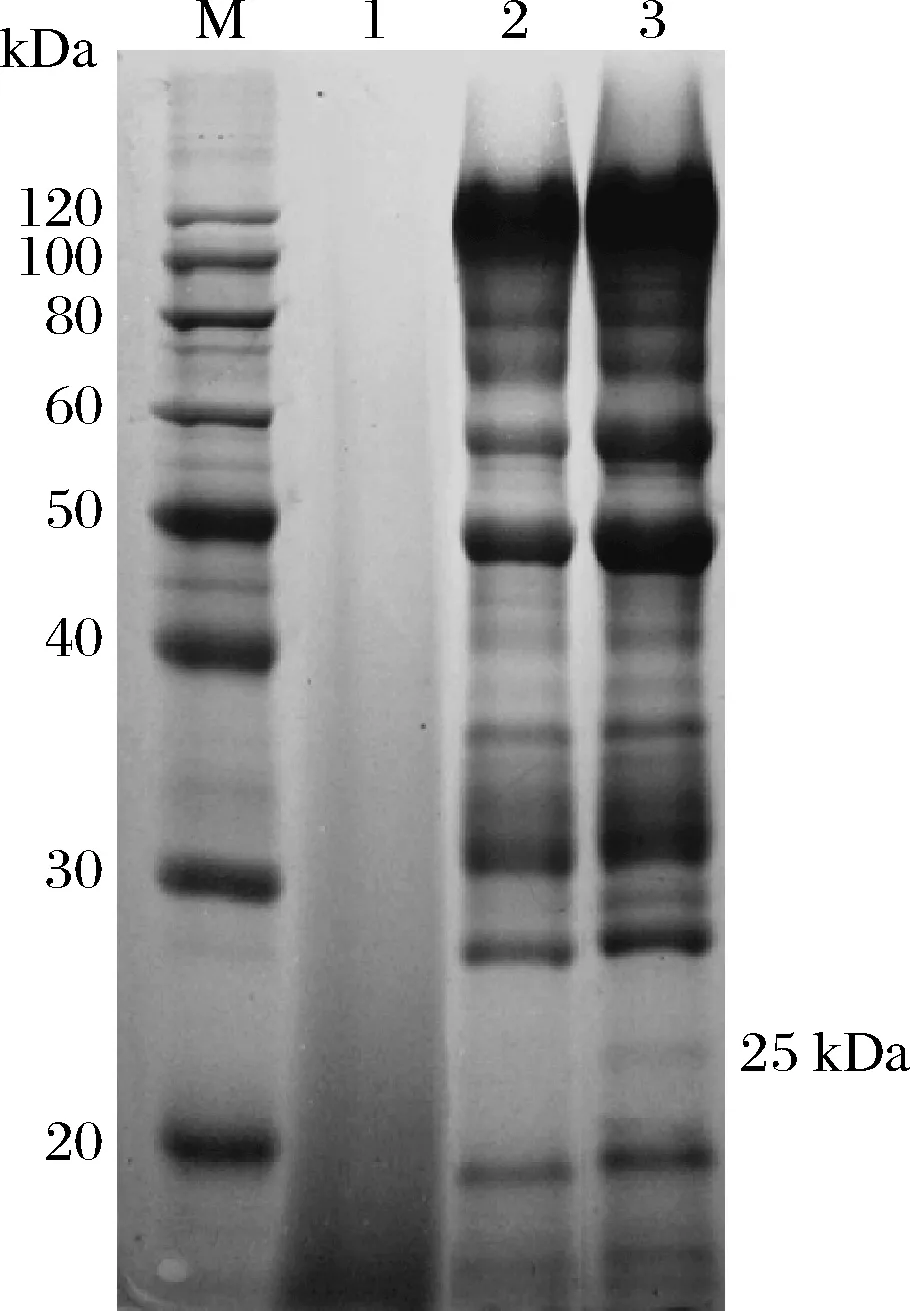
M-蛋白質(zhì)marker;1-培養(yǎng)基上清液;2-野生型菌株發(fā)酵上清液;3-TaEXPA8重組菌株發(fā)酵上清液圖7 TaEXPA8蛋白的SDS-PAGE檢測(cè)結(jié)果Fig.7 SDS-PAGE results of TaEXPA8 protein
2.6 TaEXPA8工程菌的纖維素水解作用分析
濾紙的崩解試驗(yàn)結(jié)果如圖8-A所示,相比于空白對(duì)照和纖維素酶液的單獨(dú)處理,TaEXPA8重組菌株的上清液能夠顯著促進(jìn)纖維素酶發(fā)揮功能,濾紙的崩解程度較高。葡萄糖含量的測(cè)定結(jié)果也進(jìn)一步支持這一觀點(diǎn)。對(duì)照組中葡萄糖的含量為0.064 mg/mL,添加TaEXPA8工程菌上清液的處理組相比于對(duì)照增加了21.2%,差異顯著(圖8-B)。
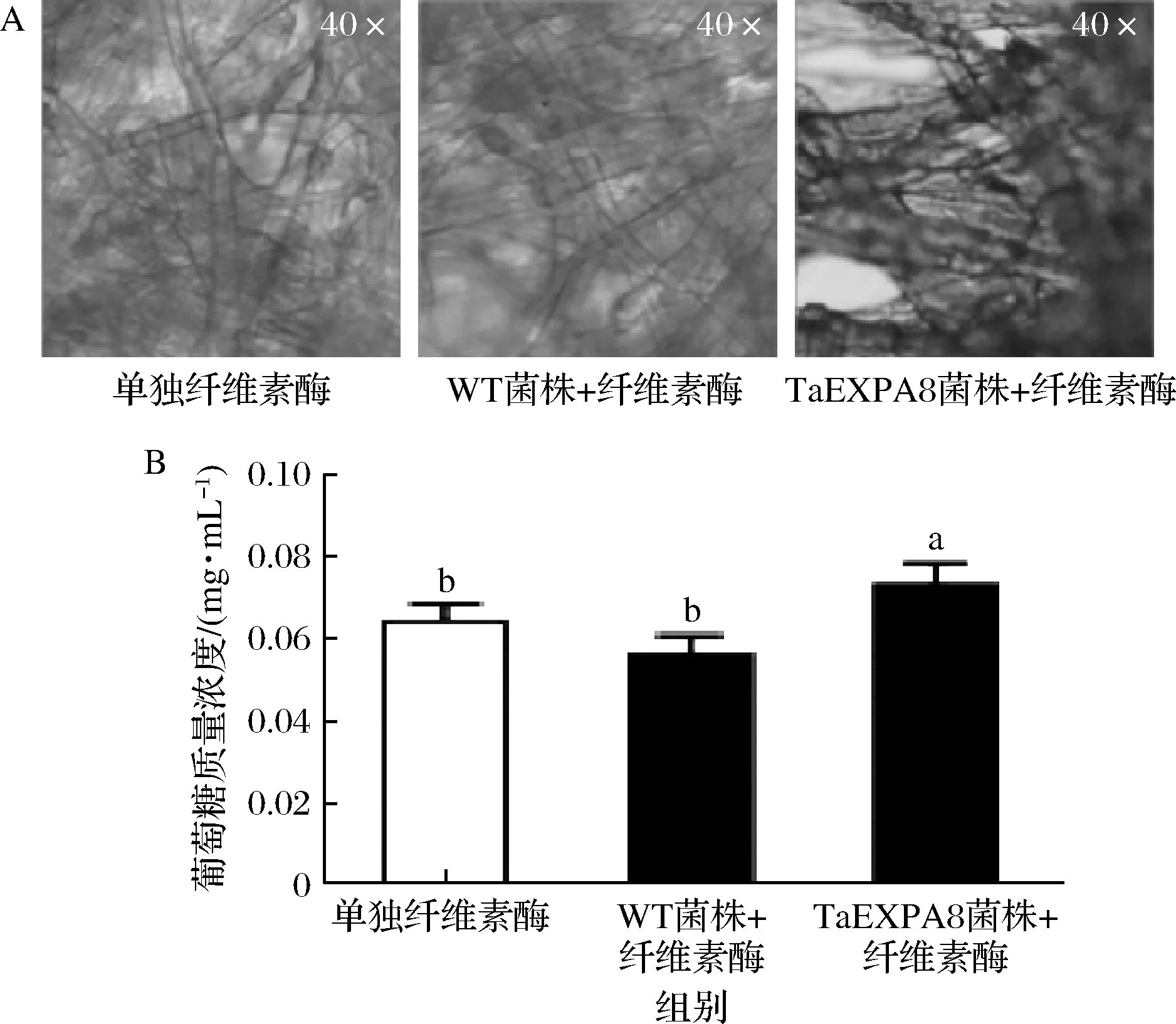
A-TaEXPA8工程菌發(fā)酵上清液的濾紙崩解作用分析;B-濾紙崩解體系中葡萄糖的含量圖8 TaEXPA8黑曲霉工程菌的纖維素水解作用分析結(jié)果Fig.8 Cellulose hydrolysis analysis results of TaEXPA8 engineered Aspergillus niger注:不同小寫(xiě)字母表示差異顯著(P<0.05)
3 討論
纖維素在自然界中的存量非常大,通過(guò)降解纖維素生產(chǎn)乙醇是現(xiàn)階段可行的能源替代方案,但尋求高效的降解方法仍是目前亟待解決的關(guān)鍵難題。生物法降解纖維素相比于傳統(tǒng)的方法具有很多優(yōu)勢(shì),但仍需要高效的酶系統(tǒng)以提高降解效率。多項(xiàng)研究顯示,擴(kuò)展蛋白能夠協(xié)同纖維素酶發(fā)揮功能并提高水解效率。原理為擴(kuò)展蛋白在破壞纖維素微纖絲之間氫鍵的同時(shí)增強(qiáng)了纖維素基質(zhì)的表面積,進(jìn)而提高了纖維素酶的吸附能力,從而使水解效率上升。在纖維素酶解時(shí)添加少量的擴(kuò)展蛋白可以顯著提高纖維素酶的活性,從而減少所需的總酶負(fù)載[19]。在本研究中,TaEXPA8工程菌的上清液顯著促進(jìn)了纖維素酶處理下濾紙的崩解,葡萄糖的測(cè)定結(jié)果也顯示水解效率顯著增加。結(jié)果表明,除了微生物中的擴(kuò)展蛋白外,來(lái)自于冬小麥的擴(kuò)展蛋白TaEXPA8在纖維素降解過(guò)程中也具有相同的功能,這為尋找高效降解纖維素的擴(kuò)展蛋白提供了新的思路。
植物擴(kuò)展蛋白的異源表達(dá)是非常困難的,直到2014年才首次實(shí)現(xiàn)來(lái)自于番茄(Lycopersiconesculentum)的擴(kuò)展蛋白LeEXP2在畢赤酵母(Pichiapastoris)中有活性的表達(dá),但最大表達(dá)量?jī)H為71 mg/L[20]。尋找合適的表達(dá)系統(tǒng)對(duì)于目標(biāo)蛋白的產(chǎn)量和活性至關(guān)重要。在本研究中,RT-PCR的結(jié)果顯示,TaEXPA8基因在工程菌中的轉(zhuǎn)錄水平較高,但SDS-PAGE的結(jié)果卻顯示蛋白的表達(dá)水平較低。在類(lèi)似的研究中,目標(biāo)基因多來(lái)自于微生物中,作為來(lái)自高等植物中的TaEXPA8可能與黑曲霉的表達(dá)系統(tǒng)不相適應(yīng),猜測(cè)這可能是蛋白表達(dá)水平較低的原因之一,更換表達(dá)載體或者對(duì)該基因進(jìn)行密碼子優(yōu)化可能是有效的解決方案。TaEXPA8工程菌的上清液使纖維素酶的水解效率提高了21.2%,對(duì)于純化的TaEXPA8蛋白,這一效率可能會(huì)更高,表明該蛋白具有潛在的應(yīng)用前景。本研究對(duì)植物擴(kuò)展蛋白在纖維素降解過(guò)程中的作用進(jìn)行了新的嘗試,雖然工程菌的產(chǎn)量較低,但對(duì)后續(xù)的相關(guān)研究也具有一定參考價(jià)值。
4 結(jié)論
本研究從冬小麥中克隆了擴(kuò)展蛋白基因TaEXPA8,并構(gòu)建了表達(dá)TaEXPA8蛋白的黑曲霉工程菌。工程菌的發(fā)酵液能夠顯著促進(jìn)纖維素酶的水解效率,但TaEXPA8蛋白的產(chǎn)量較低。同微生物中擴(kuò)展蛋白的功能相似,本研究揭示了來(lái)自于植物中的擴(kuò)展蛋白在協(xié)同纖維素生物降解過(guò)程中也具有積極作用,但異源高效表達(dá)植物擴(kuò)展蛋白的方法還需進(jìn)一步的研究以獲得,如何提高擴(kuò)展蛋白的產(chǎn)量及純度等工作仍是后續(xù)研究的重點(diǎn)。
猜你喜歡
瘋狂英語(yǔ)·初中天地(2021年5期)2021-07-21 02:24:28
甘肅教育(2020年14期)2020-09-11 07:57:42
中學(xué)生數(shù)理化(高中版.高考數(shù)學(xué))(2020年5期)2020-06-02 09:19:08
商周刊(2017年9期)2017-08-22 02:57:49
遼寧經(jīng)濟(jì)(2017年6期)2017-07-12 09:27:16
中國(guó)衛(wèi)生(2016年9期)2016-11-12 13:27:54
時(shí)代英語(yǔ)·高二(2015年1期)2015-03-16 00:08:11
中國(guó)洗滌用品工業(yè)(2015年7期)2015-02-28 19:02:38
電子設(shè)計(jì)工程(2015年12期)2015-02-27 12:06:10
中國(guó)衛(wèi)生(2014年11期)2014-11-12 13:11:32
