海螺溝冰川退縮區原生演替生態系統中鉛累積的歷史記錄
2022-02-19 12:28:40楊丹荔羅輯賈龍玉陳云飛
生態環境學報 2022年12期
關鍵詞:污染
楊丹荔,羅輯,賈龍玉 ,陳云飛
1.內江師范學院沱江流域高質量發展研究中心,四川 內江 641100;2.中國科學院、水利部成都山地災害與環境研究所,四川 成都 610041;3.成都理工大學地球科學學院,四川 成都 610059
環境問題由來已久,人們所普遍關注的重金屬污染中,鉛由于其生物毒性大且可通過大氣的遠距離運輸而影響偏遠和相對潔凈的地區(Spokes et al.,1995),對全球尺度的生態安全產生危害而備受學者們關注。在歐洲,可檢測到的大氣鉛污染早在6000年前就已開始,主要是由于土壤粉塵和農業活動。自工業革命以來,化石燃料燃燒、金屬冶煉以及汽車尾氣的排放等人類活動導致大氣中的鉛不斷增加。國外不少學者利用冰芯(Rosman et al.,1993)、湖泊及海洋沉積物(Farmer et al.,1996;Renberg et al.,2002)、泥炭沼澤(Fancois et al.,2007)及樹輪(Padilla et al.,2002)等研究大氣鉛的污染歷史,其研究結果均指出在 1950年以后是鉛沉降增強的時期,尤其是 1950—1980年是全歐洲最大的鉛排放時期。同樣在中國,尤其是在偏遠且純凈的青藏高原區,已有不少學者監測到外源污染物對該地區生態環境產生的負面影響已經凸顯,包括在該地區冰芯(李真等,2006)、湖泊沉積物(Bing et al.,2016)以及樹輪(何詠梅等,2021)中均監測到鉛污染在近幾十年以來其濃度呈現出明顯增加的趨勢。但常規用以監測和記錄重金屬污染的手段有其各自的優點,但也有不可避免的缺陷(Alley,2010;Bacon et al.,2017)。在此基礎上,不少學者越來越關注生態系統對重金屬的富集和監測作用,包括利用苔蘚監測歐洲大氣鉛污染(Stankovic et al.,2018);利用青藏高原東部林線樣帶中植物和土壤重金屬的分布監測到鉛濃度由東南向西北遞減(Luo et al.,2014,2015;Tang et al.,2014,2015)。因此,生態系統對鉛污染的響應,為鉛污染的監測提供了方便可靠的條件。而建立在冰川退縮區的植被演替序列,不僅可以較為準確的測定重金屬在生態系統中的吸收、遷移、周轉和分配特征,進而探索其在原生演替序列逐級傳遞和富集的生物地球化學過程;還可測定原生演替序列上大氣沉降輸入的重金屬通量,從而獲得演替序列時間尺度上的重金屬污染記錄,反演一定時間范圍內生態系統中重金屬的輸入情況。
位于青藏高原東緣貢嘎山東坡的海螺溝冰川退縮區,自小冰期結束以來發育了典型的土壤年代序列和完整的植被演替序列,其獨特的自然條件和演化過程,為探索復雜環境中重金屬的污染歷史記錄提供了天然的實驗場所。不少學者已在該演替序列上開展了鎘和汞的生物地球化學研究,揭示了鎘和汞在該演替序列的富集過程,并反演出近120多年來大氣鎘和汞的污染歷史(Wang et al.,2020a,2020b;佘佳,2014)。而關于鉛在該演替序列的研究包括苔蘚對大氣鉛污染的監測(Wang et al.,2019)、表層土壤鉛的富集(楊丹荔等,2020)以及利用鉛同位素對其來源示蹤(Bing et al.,2014)等。但有關整個生態系統鉛的富集特征和積累過程的研究還不夠完善。因此,本文以海螺溝冰川退縮區為載體,研究各演替階段生態系統中鉛的富集特征并利用 1890—2017年生態系統鉛的積累速率來反演該演替序列鉛的污染過程。這對貢嘎山東坡生態系統重金屬含量與貯量的研究具有重要意義,同時豐富了相關研究的方法和數據,為重建重金屬的污染歷史提供科學依據。
1 材料與方法
1.1 樣地的設置
結合海螺溝冰川最早的科學記載資料(1930年)以及先鋒樹種定居(即冰川消退到樹木幼苗發芽之間的時間)和冰川退縮區最大樹木年齡的時間間隔確定各樣地的冰川退縮時間,并通過樹木年輪進行校正。根據2017年野外實地調查資料,分別在冰川退縮時間的 2017年(S0),2000年(S1),1980年(S2),1970年(S3),1958年(S4),1930年(S5)和1890年(S6)布設7個連續樣地(圖1)。S0為冰川退縮后形成的裸地。在冰川退縮后的第4年開始有先鋒物種入侵,在S1樣地,一些灌叢及喬木幼苗如沙棘(Hippophae rhamnoides)、柳樹(Salixspp.)和冬瓜楊(Populus purdomii)等進入以黃芪(Astragalus membranaceus(Fisch.) Bung)為主的草本群落,形成演替初期的草灌階段;S2—S4樣地,冬瓜楊憑借較高的光合速率和較快的高生長優勢,占據林冠上層,形成演替中期以冬瓜楊為優勢種的落葉闊葉林群落;峨眉冷杉(Abies fabri)在冬瓜楊形成的郁閉環境下快速生長逐步成為群落優勢種,到S5樣地形成以冬瓜楊和峨眉冷杉為主的針闊混交林群落;隨著峨眉冷杉進入主林層,喜陽的冬瓜楊生長受限并逐步退出群落,最終在S6樣地形成以峨眉冷杉和麥吊云杉(Picea brachytyla)為主的針葉林頂極群落(楊丹丹等,2015)。
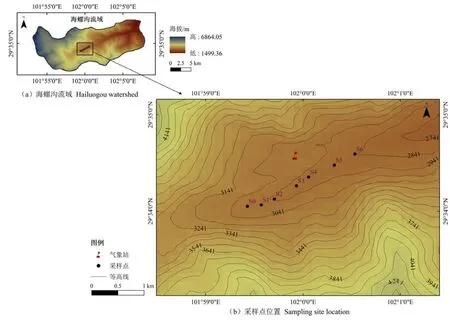
圖1 研究區及采樣點設置Figure 1 The studied area and sampling sites
1.2 野外取樣及樣品分析
作者于2017—2018年進行野外取樣。由于S0樣地為原生裸地,土壤尚未發育,僅取冰川末端堆積物細砂進行元素含量的測定。在 S1—S6各樣地隨機布設3個10 m×10 m的樣方,記錄樣方內喬木層各優勢種的名稱、胸徑、樹高等信息,并采集樣地內各優勢種干、皮、枝、葉和根各部位的樣品。根據實測的胸徑和樹高估算喬木層各優勢種的生物量。有關該地區喬木層的生物量已在相關的研究中發表(楊丹丹等,2015)。在每個10 m×10 m的樣方內,設置1 m×1 m的代表性小樣方,記錄樣方內所有灌木、草本和苔蘚的名字和數量,采用收獲法分別獲得樣方內所有灌木、草本及苔蘚,并將所有灌木、草本及苔蘚的樣品分別混合后記錄其各自的鮮重。隨后在設置0.5 m×0.5 m的小樣方并挖一個土壤剖面,按照野外土壤發育性狀,將土壤分為:Oe層,土壤有機質未完全分解的半分解層;Oa層,土壤有機質完全分解形成腐殖質的全分解層(土壤Oe層和Oa層為土壤O層);C層,母質層。由于該演替序列土壤發育時間較短,未見淀積層和淋溶層的形成。記錄每層土壤的厚度,并挑出土壤中的根系和石礫,采用環刀法測定土壤的容重,然后分層采集土壤樣品。將收集的所有樣品帶回實驗室,植物樣品置于 65 ℃烘干至恒定質量并測定其干質量,將灌木、草本、苔蘚的干質量作為其生物量,同時用作樣品元素分析。土壤樣品經自然風干后,用瑪瑙研缽粉碎至≤200目,植物等樣品經粉碎或剪碎至長度≤2 mm。將處理好的植物樣品和土壤樣品分別經鹽酸-硝酸-氫氟酸-高氯酸敞開消解和硝酸雙氧水氫氟酸微波消解后,其全溶液通過 ICP-MS分別測定鉛的含量。
1.3 數據分析
(1)表層土壤鉛的富集系數,其計算公式為(Aloupi et al.,2001):
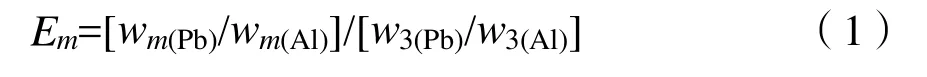
式中:
E——富集系數;
m——土層;
w3(Pb)/w3(Al)——C層鉛含量與鋁含量之比;
wm(Pb)/wm(Al)——土壤某層中鉛含量與鋁含量之比。
(2)生物轉運系數,為植物某部位重金屬含量與該植物細根中元素含量的比值,反映了植物不同部位對重金屬的運移能力。其計算公式為:轉運系數=地上部分各部位鉛含量/細根鉛含量(陳文德等,2009)。
(3)生物富集系數,為植物某部位某元素含量與土壤該元素含量之比,反映植物從土壤中攝取該元素的能力。當富集系數>1時,表征該元素在植物體內富集。其計算公式為:生物富集系數=植物各部位鉛的含量/土壤鉛的含量。(其中土壤鉛的含量以各層的厚度為權重通過加權平均得到)(董林林等,2008)。
(4)生態系統各層鉛的貯量及積累速率:植被各層鉛的貯量通過生物量與其含量的乘積得到,土壤各層鉛的貯量通過含量與厚度和容重的乘積得到。各層的積累速率為:

式中:
R——積累速率;
P——各演替階段活體植被和土壤的鉛貯量;
t——冰川退縮的時間;
i=1—6,即S1—S6樣地。
本文的研究區位置圖采用Arcgis軟件繪制,數據分析及研究結果圖采用 Origin 2020軟件進行繪制,研究區的大氣軌跡圖通過 HYSPLIT后向軌跡模型進行繪制。
2 研究結果與分析
2.1 土壤鉛的富集特征
以 S0樣地所采集的冰川細砂中鉛的含量作為該地區的背景值,土壤Oe層的鉛含量在S4—S6樣地高于背景值,土壤Oa層的鉛含量在各樣地均高于背景值,土壤C層的鉛含量除S6樣地外均低于背景值,且土壤各層鉛的含量在 S2—S6樣地主要表現為Oa層>Oe層>C層(圖2)。此外,土壤Oe層鉛含量為 14.69—52.03 mg·kg?1,Oa層為 33.00—68.47 mg·kg?1,且土壤Oe層和Oa層中鉛含量的變化趨勢較為一致,均隨演替呈先增長后下降的趨勢;土壤 C 層鉛含量在 15.95—30.16 mg·kg?1,隨演替的波動較小。
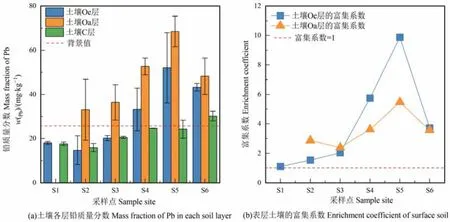
圖2 不同樣地土壤各層鉛質量分數及表層土壤的富集系數Figure 2 Mass fraction of Pb in each soil layer and enrichment coefficient of surface soil
土壤Oe層和Oa層鉛的富集系數隨演替呈先增長后下降的趨勢,且均在S5樣地分別達最大值9.88和5.47,遠高于富集系數的臨界值1,說明該演替序列土壤中的鉛表現出明顯的表層富集,尤其是土壤Oe層在S5樣地的富集系數接近10,顯示出該演替序列土壤O層中的鉛并非來源于成土母質,極大可能受凋落物的分解或大氣鉛沉降的影響導致含量較高。
2.2 活體植被鉛的富集特征
2.2.1 優勢喬木
從各樣地優勢喬木不同部位的鉛含量來看(圖3),沙棘表現為細根>皮>葉>中根>粗根>枝>干,柳樹為細根>葉>中根>枝>皮>粗根>干,冬瓜楊為細根>枝>中根>葉>皮>粗根>干,峨眉冷杉為細根>枝>中根>葉>粗根>皮>干。可見,各優勢喬木均表現為細根中的鉛含量最高,干中的鉛含量最低。此外,沙棘地下部分根的平均鉛含量(0.32—4.6 mg·kg?1)比地上部分各部位平均鉛含量(0.45—3.31 mg·kg?1)高 1.47倍,柳樹地下部分(0.49—21.1 mg·kg?1)比地上部分(0.34—3.12 mg·kg?1)高 3.45倍,冬瓜楊地下部分(0.57—16.2 mg·kg?1)比地上部分(0.35—7.08 mg·kg?1)高 1.71倍,峨眉冷杉地下部分(0.60—7.4 mg·kg?1)比地上部分高(0.25—7.43 mg·kg?1)高 1.55倍。可見,各優勢喬木不同部位的鉛含量均表現為地下部分>地上部分。
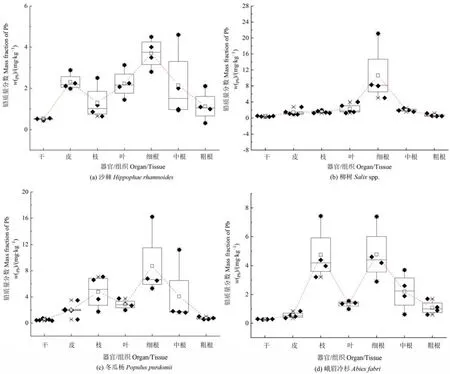
圖3 各優勢喬木不同部位的鉛質量分數Figure 3 Mass fraction of Pb in different parts of dominant trees
為進一步說明鉛在喬木各部位的遷移與富集,本文引入生物轉運系數和生物富集系數反映植物各部位對鉛的運移能力和富集程度。從喬木地上各部位的生物轉運系數來看(圖 4),沙棘皮的運移能力最高,柳樹葉的運移能力最高,而冬瓜楊和峨眉冷杉枝的運移能力最高。此外,沙棘地上各部位的轉運系數最高,其次為峨眉冷杉、冬瓜楊和柳樹。但除峨眉冷杉枝的轉運系數 (1.09)>1以外,其余優勢喬木各部位的轉運系數均<1。從喬木各部位的生物富集系數來看,除峨眉冷杉皮中鉛的富集系數較高外,其余均在細根中的富集系數最高。此外,各優勢喬木地下部分的富集系數均高于地上部分,且柳樹的富集系數最高,其次為冬瓜楊、沙棘和峨眉冷杉。由此可見,各優勢喬木地上部分的運移能力較低,而地下部分的富集程度均高于地上部分,根系所吸收的鉛可能大部分被存留在根系組織中。
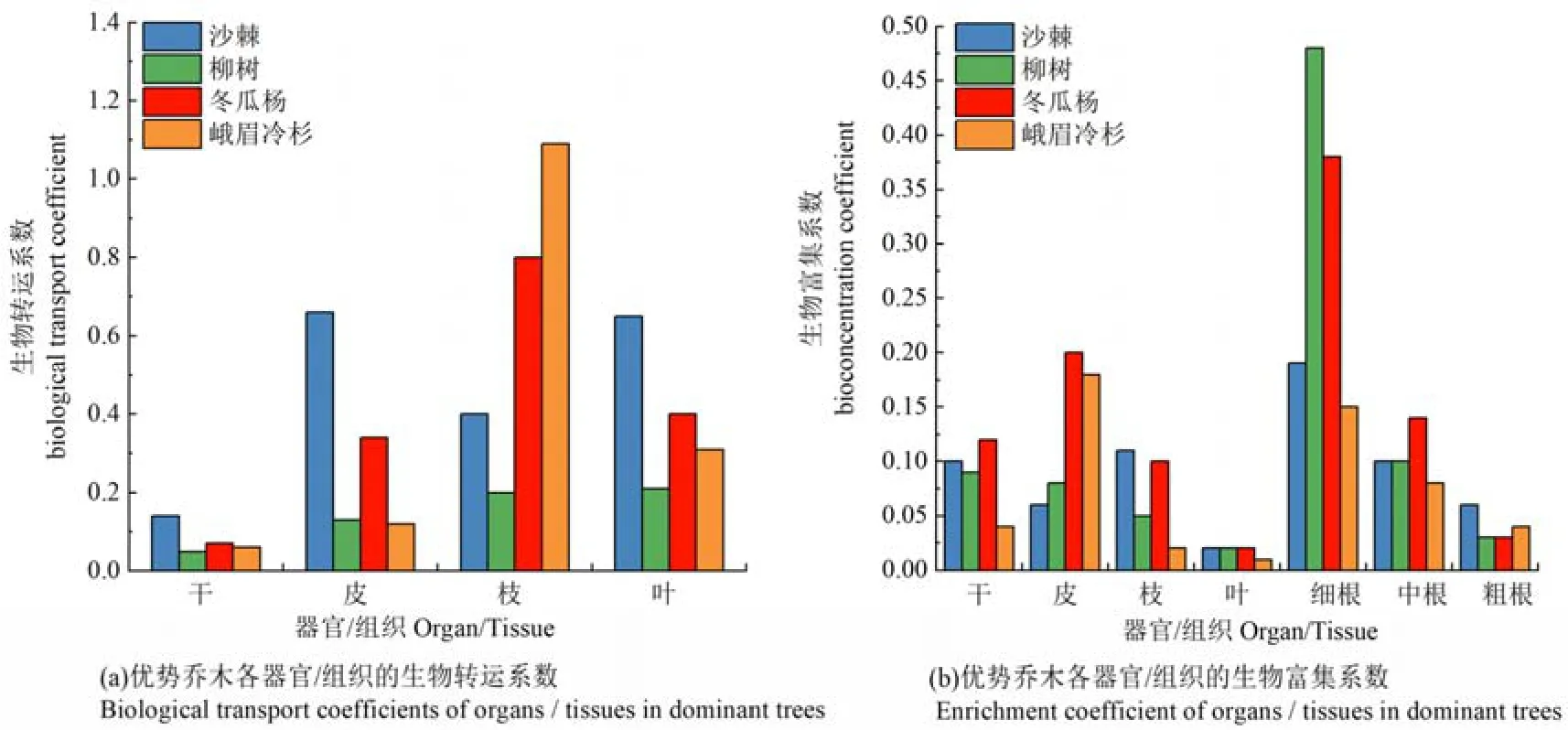
圖4 各優勢喬木不同部位鉛的生物轉運系數和生物富集系數Figure 4 Biological transport coefficient and bioaccumulation coefficient of Pb in different parts of dominant trees
2.2.2 林下植被層
灌木層枝鉛的含量高于葉,但二者在整個演替序列的變化不大(圖5);草本層鉛的含量在S1樣地最高達8.7 mg·kg?1,且隨冰川退縮時間的增加呈先下降后增長的趨勢;地被層(苔蘚)鉛的平均含量為17.23 mg·kg?1,尤其是S1樣地地被層的鉛含量高達31.62 mg·kg?1,并超過該樣地表層土壤中鉛的含量,且明顯高于林下的其他植被層。可見,林下植被中地被層對鉛的富集能力較強,可能會造成生態系統中鉛等有毒重金屬在其生物量中的積累。
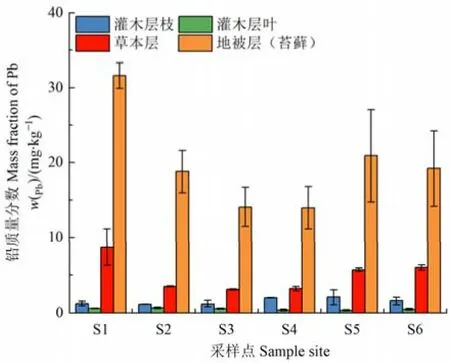
圖5 林下各植被層鉛質量分數Figure 5 Mass fraction of Pb in each undergrowth vegetation layer
2.3 生態系統鉛的積累動態
生態系統鉛的總貯量隨演替呈波動增長的趨勢(圖6),從S1樣地的64.64 kg·hm?2增長到S6樣地的 227.16 kg·hm?2。其中,活體植被的鉛貯量隨演替從S1樣地的0.01 kg·hm?2增長到S5樣地的0.58 kg·hm?2,到S6樣地略有下降,而土壤鉛貯量從S1樣地的64.83 kg·hm?2增長到S6樣地的226.68 kg·hm?2。可見,各樣地土壤鉛貯量占生態系統總貯量的99%以上,是生態系統鉛的最大儲存庫。
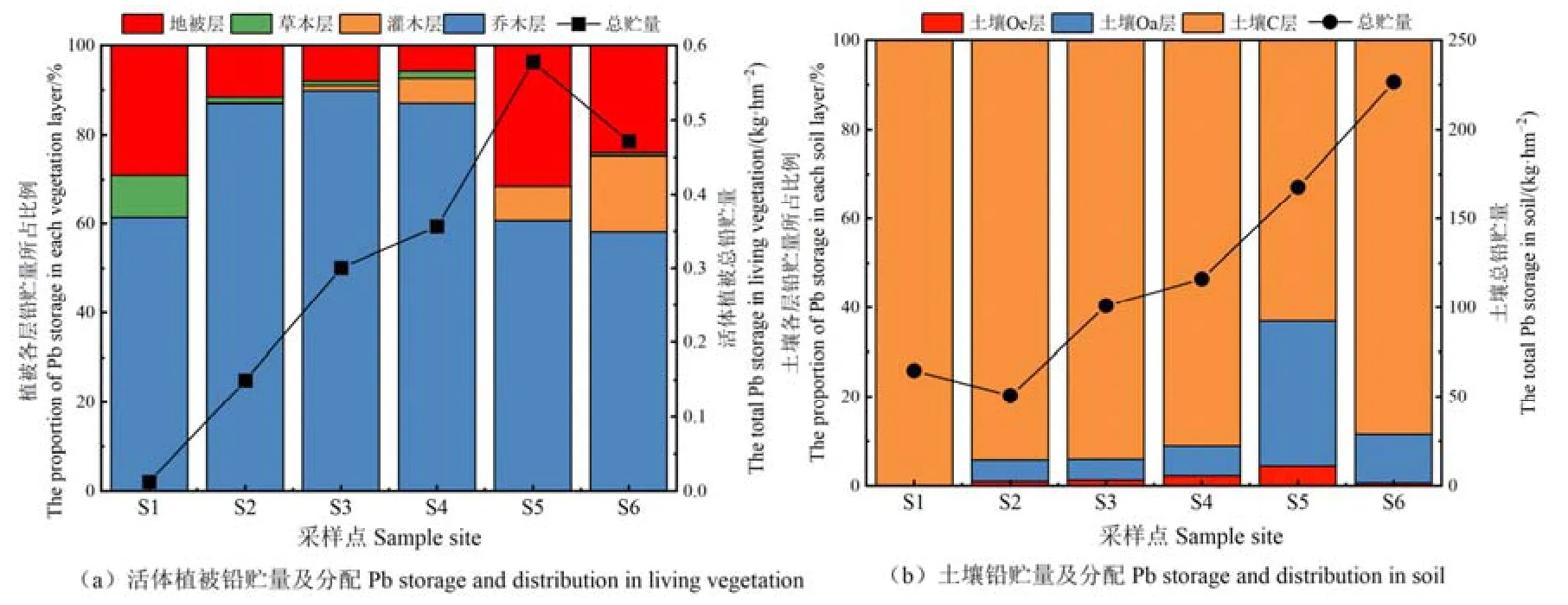
圖6 生態系統活體植被與土壤鉛的貯量Figure 6 Pb storage in living vegetation and soil of ecosystem
在活體植被中,喬木層鉛貯量隨演替逐漸增加,到S5樣地達最大值0.35 kg·hm?2,其所占比例在各樣地均達60%以上,尤其是S3樣地所占比例達90%。其中,S1樣地喬木層中沙棘的鉛貯量最高,但隨植被的演替,到S4樣地冬瓜楊的鉛貯量達0.27 kg·hm?2,約占整個活體植被鉛貯量的76%;到演替末期形成以峨眉冷杉為主的針葉林后,峨眉冷杉的鉛貯量為0.26 kg·hm?2,約占活體植被鉛貯量的50%。可見,各優勢喬木鉛貯量的動態變化與其演替趨勢是一致的。灌木層的鉛貯量隨演替從S1樣地的0.02 g·hm?2增長到 S6 樣地的 81.02 g·hm?2,但在整個演替序列中其鉛貯量僅占活體植被鉛貯量的5.35%。草本層鉛貯量呈先增長后下降的趨勢,并在S4樣地達最大值 6.02 g·hm?2,其貯量占活體植被鉛貯量的比重也非常小,僅為2.38%。地被層(苔蘚)鉛貯量的變化趨勢與喬木層一致,均在S5樣地達最大值0.18 kg·hm?2,其占活體植被鉛貯量的比重最大可達36%。因此,喬木層和地被層是活體植被中鉛的主要存貯單元,而灌木層和草本層的鉛貯量較低且所占比重小,對鉛在活體植被中的分配格局影響較小。
土壤鉛總貯量隨演替呈波動的增長趨勢,在S2樣地略有下降。土壤Oe層和Oa層的鉛貯量隨演替呈先增長后下降的趨勢,并在 S5樣地分別達最大值 7.50 kg·hm?2和 54.35 kg·hm?2,土壤 C 層的鉛貯量在整個演替序列呈波動增長的趨勢。
此外,生態系統鉛的積累速率不僅能夠量化整個演替序列對鉛的積累過程,還能反演一定時間范圍內鉛的污染歷史。本文根據貢嘎山東坡樹木年輪(距離海螺溝冰川退縮區峨眉冷杉頂極群落直線距離約200 m)反演該地區百年來重金屬污染歷史(何詠梅等,2021),并結合海螺溝冰川退縮區原生演替序列120多年來生態系統鉛的積累過程,進一步說明生態系統對鉛污染的監測作用。從圖7可以看出,1860—1920年間,樹輪中鉛的含量較低且變化較小,1920年以后樹輪中的鉛含量呈波動性增長,尤其是1905—1955年以及1970—1995年這兩個時間范圍內,樹輪中鉛的含量增長較為明顯。從海螺溝冰川退縮區原生演替生態系統鉛的積累速率來看,1890—1930年生態系統鉛的積累速率變化平穩,1930年以后則呈現出波動性增長,尤其是在1958—1970年和1980—2000年這兩個時間范圍內生態系統對鉛的積累較為明顯,并且能很好的與樹輪中鉛含量的變化趨勢相對應。這說明生態系統對鉛的反應較為敏感,其植被和土壤對鉛污染的響應能夠反演近百余年時間尺度上鉛的污染記錄。
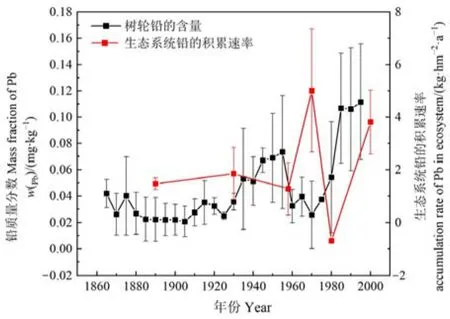
圖7 生態系統鉛的貯量與樹輪鉛質量分數Figure 7 Pb storage in ecosystem and mass fraction of Pb in tree rings
3 討論
3.1 原生演替生態系統對鉛的富集作用
不同演替階段各優勢喬木地上部分鉛含量及其轉運能力和富集程度均較低,而細根中鉛的含量和富集程度明顯高于其他部位。可見植物根部所吸收的鉛絕大部分被存留在根系組織,只有很少一部分鉛借助共質體被轉移到地上部分(夏時雨等,1994)。這主要是由于土壤中的鉛大部分以Pb(OH)2、PbCO3等難溶性鹽及有機絡合態存在,導致土壤中鉛的移動性較差且植物有效性低(唐榮貴,2015)。此外,植物根系所吸收的鉛在向地上部分轉運的過程中還會受到內皮層組織中凱氏帶屏障作用的影響,進一步阻擋鉛的輸送。而土壤中的鉛能被根毛吸附并保留在細胞壁上從而促進細根對鉛的吸收(Kabata-Pendias,2000;Uzu et al.,2010)。因此,鉛經過根系吸收再向植物地上部位運移較為困難。雖然葉片能夠吸附大氣中的鉛而導致其含量較高(陳英旭等,2000),但該植被演替序列各優勢喬木葉片的鉛含量并沒有明顯高于其他地上部分。因此,受土壤鉛的存在形態以及植物根系不同部位和組織差異化分布的影響,導致該演替序列各優勢喬木地下根系鉛的含量明顯高于地上部分。值得注意的是,峨眉冷杉枝鉛的含量與細根鉛的含量相當,且枝中鉛的轉運系數也明顯高于峨眉冷杉及其他優勢喬木不同部位的轉運系數。這可能是由于針葉根系較強的有機酸分泌能力使得土壤 pH值下降從而增加土壤溶液中鉛的溶解度(夏時雨等,1994)。此外,有機酸還可以促進鉛從植物根系向地上部分轉移(Tyler,1990),而導致峨眉冷杉枝的鉛含量較高。林下植被中,地被層(苔蘚)鉛的平均含量高達 17.23 mg·kg?1,明顯高于喬木、灌木各部位以及草本層的鉛含量,且苔蘚生物量對該演替序列生態系統活體植被總生物量的貢獻不足 5%(楊丹丹等,2015),但其鉛的積累貢獻最大可達36%,表現出對鉛的明顯富集。在早期的研究中,已有不少學者利用苔蘚生物監測大氣有毒金屬污染水平(Boutron et al.,1991;Stankovic et al.,2018),并指出苔蘚具有較強的鉛吸收能力(Wang et al.,2019)。而苔蘚植物主要分布于高山高寒地區。因此,基于苔蘚在高山生態系統中的分布范圍較廣且對鉛的吸收能力較強,可能會造成這些地區對鉛的富集。
表層土壤Oe層和Oa層的鉛含量在整個演替序列的變化趨勢一致,表現出隨演替先增長后下降的趨勢。從土壤O層鉛的富集系數來看,其含量的變化并非受成土母質的影響,而可能受到來自凋落物歸還、根系以及大氣沉降等因素的影響(楊丹荔等,2020)。該演替序列中期形成以冬瓜楊為主的落葉闊葉林群落,其凋落量達整個演替階段的最大值(楊丹丹等,2015),枝和葉等吸收的鉛通過凋落物分解歸還于土壤導致表層土壤鉛含量增加;根系作為該演替序列優勢種鉛富集的主要部位,同樣會導致土壤中的鉛含量增加。此外,到 S6樣地,土壤O層中鉛的含量有所下降,這可能與該演替序列表層土壤pH值的變化有關。S1樣地表層土壤的pH值高達8.54(楊丹荔等,2020),隨著植被的演替尤其是 S6樣地形成以峨眉冷杉為主的針葉林后,一方面針葉樹種根系的外生菌根分泌有機酸的能力較強;另一方面針葉樹種凋落物的酸性較低,這導致S6樣地表層土壤的pH值僅為4.3,表現出明顯的酸化。而土壤的pH值越低,土壤對鉛的“固定”能力和吸持能力也越弱(王春苗等,2009)。因此,在酸性土壤條件下,鉛的淋溶遷移能力較強,而滯留在土壤中的鉛相對減少(Glover et al.,2002;潘勝強等,2014),這也導致該演替序列表層土壤鉛含量在演替末期略有下降。
3.2 原生演替生態系統鉛反演的污染歷史
常用的元素地球化學記錄的自然檔案有冰巖芯、樹木年輪、湖泊沉積物以及黃土剖面等。通過對比分析冰芯、湖泊沉積等對重金屬的污染歷史記錄可知,在格陵蘭(Boutron et al.,1991)和阿爾卑斯山 Monta Rosa地區(Schwikowski et al.,2004)的冰芯中發現在 20世紀 70年代鉛的含量達到峰值;在加拿大Clair湖芯(Ndzangou et al.,2005)中也發現 1975年沉積物中的鉛濃度明顯增高。而在研究青藏高原地區重金屬污染歷史的結論中發現,帕米爾高原的慕士塔格冰芯(李真等,2006)中的鉛濃度在1970—1990年間出現兩個峰值,隨后逐漸下降;在喜馬拉雅山系中段的希夏邦馬峰達索普冰芯(霍文冕等,1999)中發現 20世紀 70年代以后的鉛濃度增長非常明顯;在貢嘎山草海子湖泊沉積中鉛的人為通量在 1950年以后顯著增加,并在20世紀90年代中期達到峰值,隨后逐漸減小(Bing et al.,2016);貢嘎山東坡峨眉冷杉樹輪中鉛的含量在1905—1955年以及1970—1995年呈上升趨勢。由此可見,無論從全球還是青藏高原地區鉛的污染歷史記錄來看,工業革命以來冰芯、湖泊沉積以及樹輪所記錄的鉛含量總體呈波動增長的趨勢并在1970年左右達到峰值,隨后略有下降。
已有學者利用海螺溝冰川退縮區原生演替生態系統反演近百年來鎘的污染歷史(Wang et al.,2020a)。同樣,該原生演替序列生態系統中的鉛經歷了從無到有的積累過程,其積累速率在冰川退縮1890—2000年間呈波動變化,尤其是在1958—1970年間積累較快,并在1970年達最大值,隨后略有下降到 1980以后又有所回升。可見,該演替序列生態系統鉛的積累過程能夠很好的與青藏高原乃至全球冰芯、湖泊沉積以及樹輪等對鉛的污染歷史記錄相對應,反映同一時期鉛污染的歷史狀況。貢嘎山地區的大氣運輸在冬季和春季主要受到西風帶的影響,在夏季主要受到印度季風的影響(Fu et al.,2009)。通過 HYSPLIT后向軌跡模型計算出2021年海螺溝冰川退縮區的大氣軌跡圖(圖8),可知該地區的大氣污染物的輸入可能主要來自于中亞和南亞地區。這些地區對 1958—1970年大氣中鉛沉降的上升均有影響,例如,印度在 20世紀50—80年代末,礦物燃料排放的廢氣位居世界第9(Pan et al.,2015);中亞5國作為前蘇聯主要的軍事工業基地,在該時期美蘇的兩次大規模軍備競賽中可能造成污染物的大量排放(Golomb et al.,1997)。貢嘎山位于中國西南地區,已有研究表明該地區受到來自四川東北部和云南西北部等地區鉛污染長距離傳輸的氣團影響(Golomb et al.,1997),而中國直到 1970年以后大氣污染防治才開始起步。此外,在過去120多年里,該演替序列生態系統鉛的平均積累速率為 2.12 mg·m?2·a?1,但受輸出通量的影響,大氣平均積累速率通常要低于大氣總的沉降通量。例如,中國北方大氣鉛總沉降通量(14.1 mg·m?2·a?1)比平均鉛積累速率高 3 倍(Luo et al.,2015);而在歐洲和北美地區則比鉛的積累速率高1—3倍(Kyll?nen et al.,2009)。貢嘎山地區屬于偏遠地區,當地沒有明顯的工業污染排放和人類活動,生態系統中重金屬多源于外來輸入。若未來大氣鉛的沉積通量仍然升高,這將導致貢嘎山地區存在潛在的鉛污染風險。
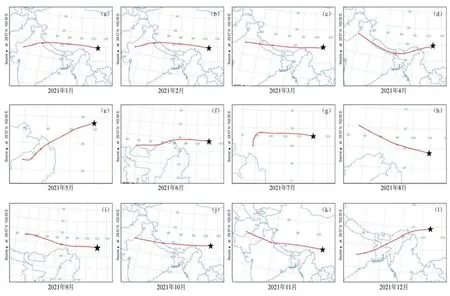
圖8 海螺溝冰川退縮區大氣軌跡圖Figure 8 Atmospheric trajectory of Hailuogou Glacier Retreat area
4 結論
海螺溝冰川退縮區原生演替生態系統對鉛的富集主要集中于土壤表層、優勢喬木的細根以及地被層的苔蘚中,且隨冰川退縮時間的增加,生態系統對鉛的貯存也隨之增加并主要貯存于土壤之中。但值得注意的是,苔蘚的生物量對總生物量的貢獻不足5%,但其對植被鉛的積累貢獻最大可達36%,表現出明顯的富集能力,這對高山高寒等地區的生態系統可能造成潛在的鉛污染。
通過原生演替生態系統對鉛的積累過程反演出近百年來鉛污染在 1958—1970年和 1980—2000年這兩個時間范圍內生態系統積累較快,尤其是在1970年出現明顯的峰值。這與青藏高原冰芯、貢嘎山東坡湖泊沉積和樹輪中鉛含量的變化趨勢相對應,說明原生演替序列提供了鉛在百年時間尺度的生物地球化學記錄,也進一步證明鉛污染通過遠距離的大氣傳輸已影響到相對偏遠且潔凈的地區。
猜你喜歡
小學科學(學生版)(2021年8期)2021-08-30 09:16:18
小學科學(學生版)(2021年7期)2021-07-28 06:44:32
小學科學(學生版)(2020年6期)2020-08-13 07:47:12
當代陜西(2019年7期)2019-04-25 00:22:18
領導決策信息(2018年26期)2018-10-12 02:18:26
新高考(英語進階)(2017年10期)2017-12-23 09:14:50
食品界(2016年4期)2016-02-27 07:36:15
都市麗人(2015年5期)2015-03-20 13:33:49
裝備環境工程(2015年4期)2015-02-28 01:20:10
江蘇年鑒(2014年0期)2014-03-11 17:09:46
