中華蜜蜂GABARAP基因克隆、生物信息學分析及原核表達載體的構建
2022-02-15 10:41:50于慧敏吳鵬杰李南南徐書法武江利
中國畜牧獸醫 2022年1期
于慧敏,吳鵬杰,李南南,譚 靜,徐書法,武江利
(中國農業科學院蜜蜂研究所,農業農村部授粉昆蟲生物學重點實驗室,北京 100093)
細胞自噬(autophagy)是維持細胞內環境穩態及抗病調節的重要調控途徑[1-3]。自噬的形成和發生由一系列的自噬相關蛋白(autophagy related genes,ATG)進行調控。在自噬過程中Atg8蛋白是唯一能留在膜上的ATG蛋白,可作為自噬發生的標記蛋白。Atg8蛋白家族可與磷脂酸肌醇結合,參與自噬體膜的形成影響自噬途徑[4-6]。已知的γ-氨基丁酸A受體相關蛋白(gamma-aminobutyric acid receptor-associated protein,GABARAP)是自噬相關蛋白Atg8家族的成員,在自噬過程中發揮著不可替代的作用[7]。人源GABARAP蛋白由117個氨基酸組成,分子質量約為14 ku,在進化上高度保守,其N-端亞結構區域能與微管蛋白結合,而C-端亞結構區域可與GABAA受體結合[8-9]。相關研究證明了在GABAA受體的胞內轉運過程中,GABARAP蛋白能起到組裝微管的作用,且GABRAP蛋白可通過調節GABAA受體的轉運來調控GABAA受體的突觸定位[10-11]。Wang等[12]通過免疫學方法證明了GABARAP蛋白在神經元的胞體和突觸中表達。研究發現,GABARAP蛋白是自噬發生的良好標記,對于自噬體的形成、延伸和成熟非常重要[13-15]。因此,GABARAP蛋白不僅在神經傳遞中發揮重要作用,還參與細胞自噬過程。GABARAP蛋白可激活絲氨酸/蘇氨酸激酶家族中的UNC-51激酶復合體(UNC-51 like kinase,ULK)來調節其特定自噬體的擴張[16-17]。相關研究表明,與LC3家族相比,GABARAP家族在氨基酸匱乏和線粒體自噬方面發揮著更重要的作用[18-19]。GABARAP蛋白能與一種RNA解旋酶DDX47發生相互作用,將二者共轉染到腫瘤細胞系中可誘導細胞凋亡[20-21]。白榮耀[22]研究發現,在感染溶藻弧菌3 h后,鮑魚血細胞SaGABARAP基因轉錄水平上調且達到最大值。由此可知,GABARAP蛋白在生物體細胞凋亡、免疫應答中也發揮著關鍵作用。
關于GABARAP基因的研究已從單細胞酵母逐步擴展到了高等動物和植物。但就蜜蜂而言,僅有GABARAP和LC3作為自噬相關蛋白Atg8家族成員在西方蜜蜂上進行報道,然而對GABARAP基因在自噬過程中的作用機制研究仍較少[23]。鑒于此,本研究克隆了中華蜜蜂的自噬相關基因GABARAP,利用生物信息學方法對該基因進行了序列分析及結構預測,構建了原核表達載體,誘導并純化了帶有His標簽的GABARAP蛋白,以期為中華蜜蜂GABARAP基因功能的深入研究提供理論依據。
1 材料與方法
1.1 樣品來源
3日齡中華蜜蜂幼蟲均由中國農業科學院蜜蜂研究所提供,樣品收集后于液氮迅速冷凍,于-80 ℃保存備用。
1.2 主要試劑
總RNA提取試劑購自Invitrogen公司;RNA反轉錄試劑盒購自Promega公司;TransTaq酶、T4 DNA快速連接酶均購自北京全式金生物技術有限公司;SuperReal PreMix Plus(SYBR Green)實時熒光定量試劑盒購自天根生化科技(北京)有限公司;DNA限制性內切酶BamHⅠ和NotⅠ均購自NEB公司;質粒DNA提取試劑盒、瓊脂糖凝膠DNA回收試劑盒均購自Bio-Tek公司;抗His標簽鼠單克隆抗體、HRP標記的山羊抗小鼠IgG、Ni-Agarose His標簽蛋白純化試劑盒、One Step Western Kit HRP(MOUSE)、DAB蛋白顯色試劑盒均購自北京康為世紀生物科技有限公司。
1.3 引物設計與合成
參照GenBank中的西方蜜蜂GABARAP基因序列(登錄號:XM_001120069.5),使用Primer Premier 5.0軟件在非編碼區設計引物,引物序列為:GABARAP-F1:5′-ACTGACGCACGGTGAT-TG-3′;GABARAP-R1:5′-AGCTTCTAAAAGGA-CTAC-3′。同時設計帶有酶切位點的特異性引物用于擴增開放閱讀框,引物序列為:GABARAP-F2:5′-CGCGGATCCATGAAGTTTCATTACAAAG-3′;GABARAP-R2:5′-ATTTGCGGCCGCTTAGTGT-CCATACACATTC-3′;下劃線處分別為BamHⅠ和NotⅠ酶切位點。 引物均由北京博邁德公司合成。
1.4 GABARAP基因克隆
1.4.1 中華蜜蜂總RNA提取與單鏈cDNA合成 取2只中華蜜蜂幼蟲于1.5 mL的EP管中,加入液氮迅速研成粉末,采用Trizol法提取總RNA,通過NanoDrop2000c及瓊脂糖凝膠電泳對RNA進行質控檢測。按照反轉錄試劑盒說明書將RNA反轉錄為cDNA。
1.4.2GABARAP基因巢式PCR擴增 以cDNA為模板,用引物GABARAP-F1/R1進行第1輪PCR擴增。 PCR反應體系50 μL:10×TransTaqHiFi BufferⅡ 5 μL,dNTPs 4 μL,上、下游引物各1 μL,cDNA 4 μL,TransTaqHiFi DNA Polymerase 1 μL,ddH2O 34 μL。PCR反應程序:94 ℃預變性3 min;94 ℃變性30 s,52 ℃退火30 s,72 ℃延伸1 min,共30個循環;72 ℃延伸10 min。以第1輪PCR產物為模板,用GABARAP-F2/R2引物進行第2輪PCR擴增,PCR反應體系與第1輪擴增相同。PCR反應程序:94 ℃預變性3 min;94 ℃變性30 s,60 ℃退火30 s,72 ℃延伸1 min,共30個循環;72 ℃延伸10 min。PCR產物進行1.5%瓊脂糖凝膠電泳檢測,膠回收純化獲取目的片段。
1.4.3 pEasy-T5-GABARAP重組質粒的克隆與鑒定 將回收的目的片段與pEasy-T5克隆載體在25 ℃金屬浴下連接15 min。連接體系5 μL:PCR產物4 μL,pEasy-T5克隆載體1 μL。 將連接產物轉化大腸桿菌Trans-T1感受態細胞,涂布于含氨芐青霉素(Amp)(50 μg/μL)、X-Gal(20 mg/mL)和IPTG(500 mmol/L)的LB固體培養基中培養,37 ℃培養約12 h后挑取菌落,放入具有Amp抗性的LB液體培養基中震蕩培養過夜。 利用菌液PCR方法鑒定分析pEasy-T5-GABARAP陽性克隆,并測序鑒定。
1.4.4 生物信息學分析 用ORFfinder(https:∥www.ncbi.nlm.nih.gov/orffinder/)在線軟件對中華蜜蜂GABARAP基因的陽性克隆序列進行核苷酸翻譯;利用NCBI的BLAST程序對中華蜜蜂GABARAP基因序列進行比對;篩選出與GABARAP氨基酸序列相似性高的物種;應用Mega 5.1軟件對氨基酸序列進行對比并利用鄰接法建立系統進化樹;采用PrediceProtein(https:∥www.predictprotein.org/)在線分析軟件預測中華蜜蜂GABARAP蛋白二級結構及組成;通過SWISS-MODEL(https:∥swissmodel.expasy.org/)在線預測GABARAP蛋白三維結構[24]。
1.5 GABARAP的原核表達
1.5.1 pET-32a(+)-GABARAP重組質粒的構建與鑒定 用限制性內切酶BamHⅠ和NotⅠ將pEasyT5-GABARAP與pET-32a(+)原核表達載體于37 ℃酶切3 h,對雙酶切產物鑒定并回收純化,然后用T4 DNA連接酶連接。反應體系10 μL:5×T4 DNA Ligase Buffer 2 μL,pET-32a(+)酶切產物1 μL,pEasyT5-GABARAP酶切產物5 μL,T4 DNA連接酶1 μL,ddH2O 1 μL。25 ℃連接15 min。將重組質粒連接產物轉化大腸桿菌DH5α感受態細胞,篩選陽性克隆并測序。最后將測序正確的陽性重組質粒轉化大腸桿菌BL21(DE3)感受態細胞,涂布于具有Amp抗性的LB固體培養基中培養過夜。
1.5.2 GABARAP蛋白的誘導表達 挑取含有pET-32a(+)-GABARAP重組質粒的BL21(DE3)陽性菌,在含有氨芐青霉素(50 μg/mL)的LB液體培養基中振蕩培養直至D600 nm值為0.8,保存菌液,取部分菌液作為對照組,余下菌液按1∶1 000加入IPTG進行誘導,于37 ℃搖床中振蕩培養4 h后,取出少量菌液,離心后去上清,沉淀用PBS重懸后加入等體積的2×SDS-PAGE上樣緩沖液,沸水中煮5 min,進行SDS-PAGE,之后用考馬斯亮藍染色初步鑒定GABARAP蛋白是否表達。
1.5.3 GABARAP蛋白的Western blotting鑒定 將經SDS-PAGE的蛋白條帶轉印至NC膜上,采用抗His標簽鼠單克隆抗體進行一抗孵育,山羊抗小鼠IgG(H+L)二抗孵育后用One Step Western Kit HRP(MOUSE)進行顯色,進一步驗證表達產物。
1.6 GABARAP蛋白的純化
1.6.1 GABARAP蛋白可溶性分析 對誘導的菌液用超聲波破碎,經12 000 r/min離心30 min后,分別取上清和沉淀進行Western blotting,鑒定其表達形式。
1.6.2 GABARAP蛋白包涵體純化 按照His標簽蛋白純化試劑盒操作說明書對包涵體蛋白進行純化,將超聲處理后的沉淀依次在包涵體洗滌液Ⅰ和洗滌液Ⅱ中進行重懸溶解,在含8 mol/L尿素的變性液中進行變性,然后在不同濃度尿素的復性液中低溫透析,最后置換到PBS中,獲得純化蛋白[25]。
2 結 果
2.1 GABARAP基因克隆
中華蜜蜂幼蟲總RNA經巢式PCR擴增結果顯示,第1輪PCR擴增獲得大小約606 bp的目的片段,第2輪PCR擴增得到大小約354 bp的目的片段(圖1),均與預期相符。將目的片段成功轉化后,利用GABARAP-F2/R2引物經菌液PCR方法擴增得到大小為354 bp的片段(圖2),與第2輪PCR擴增的目的片段一致,且測序結果證明了該片段為GABARAP基因的有效序列,方向正確。NCBI同源檢索表明中華蜜蜂GABARAP基因與西方蜜蜂GABARAP基因的序列一致性為100%。
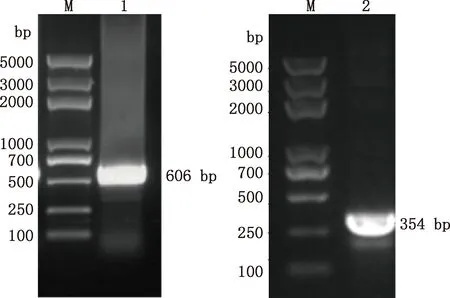
M,DL5000 DNA Marker;1,第1輪PCR產物;2,第2輪PCR產物M,DL5000 DNA Marker;1,The first round PCR product;2,The second round PCR product圖1 中華蜜蜂GABARAP基因巢式PCR擴增結果Fig.1 Nested PCR amplification results of GABARAP gene of Apis cerana cerana
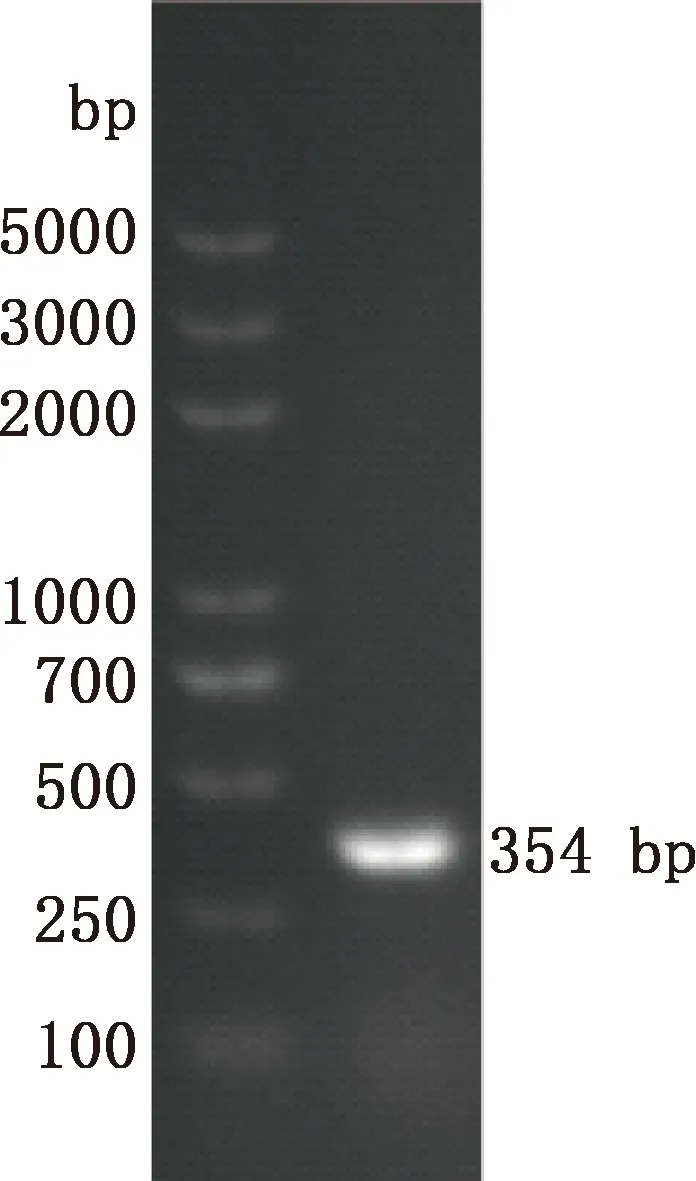
圖2 重組質粒pEasy-T5-GABARAP的PCR檢測Fig.2 PCR detection of recombinant plasmid pEasy-T5-GABARAP
2.2 GABARAP相似性比對與系統進化分析
GABARAP蛋白氨基酸序列分析顯示,GABARAP基因的354 bp有效序列共編碼117個氨基酸,分子質量為13.99 ku,等電點為9.48。
BLAST在線分析結果顯示,中華蜜蜂GABARAP氨基酸序列與西方蜜蜂、大蜜蜂、小蜜蜂、歐洲熊蜂、東方熊蜂的氨基酸序列相似性均高達100%;與蘆蜂的GABARAP氨基酸序列相似性也較高,達99%(表1)。由構建的進化樹可知,中華蜜蜂與同為膜翅目的其他昆蟲如西方蜜蜂、小蜜蜂、大蜜蜂、歐洲熊蜂和東方熊蜂親緣關系最近,處于同一簇中(圖3)。
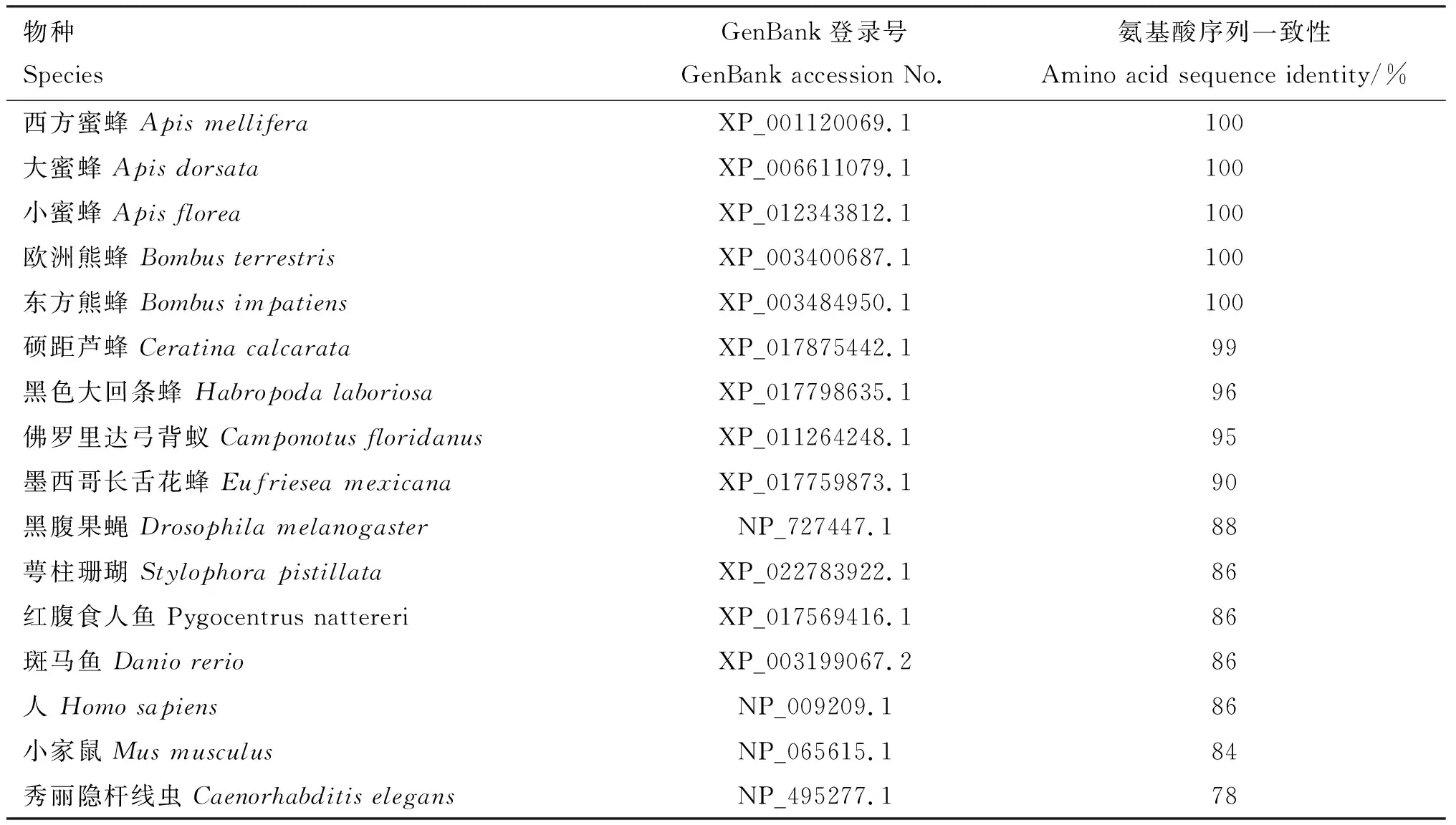
表1 中華蜜蜂GABARAP蛋白與其他物種同源蛋白氨基酸序列一致性比對
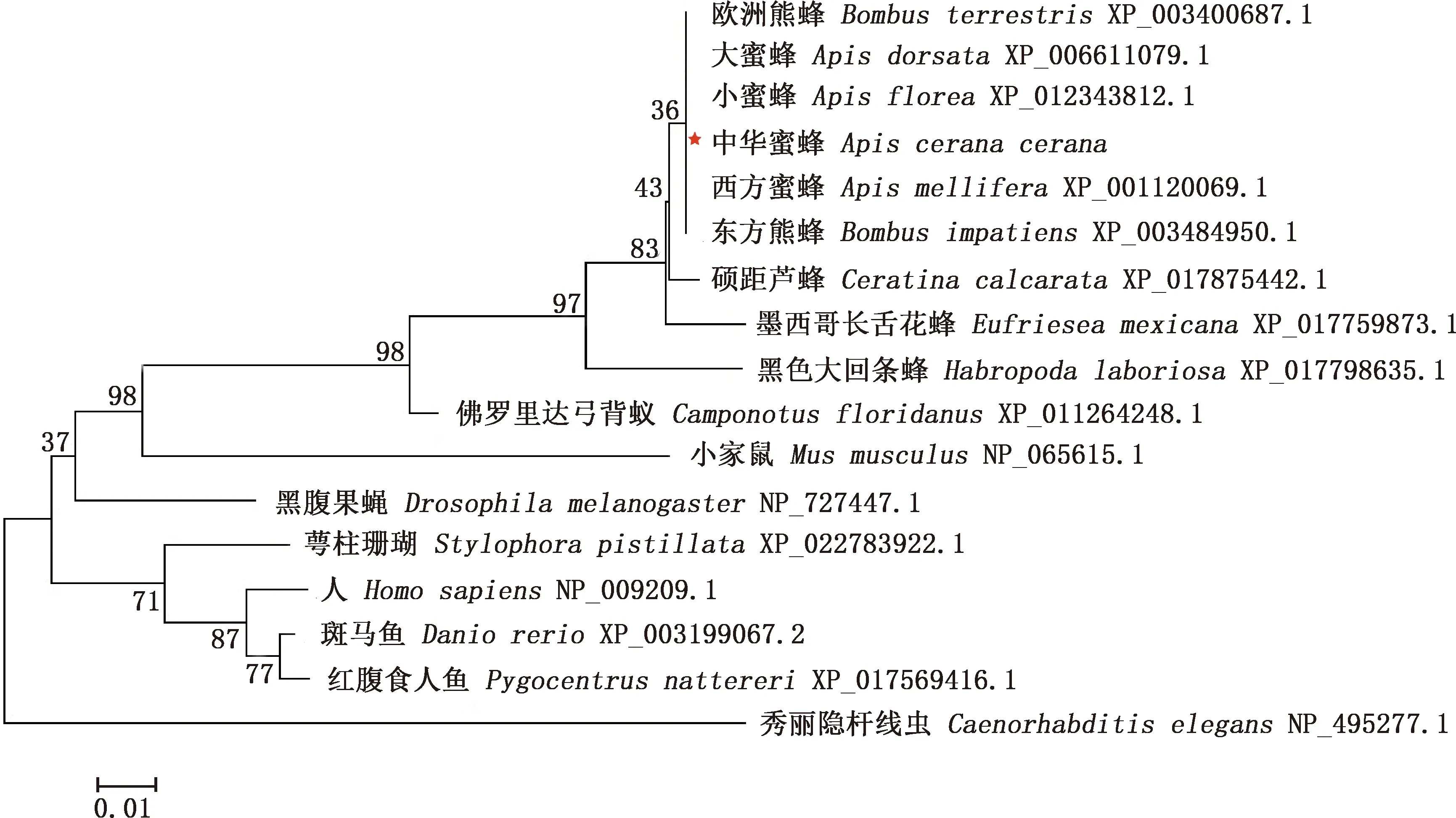
圖3 基于中華蜜蜂與其他物種GABARAP氨基酸序列的系統進化樹Fig.3 Phylogenetic tree based on amino acid sequences of GABARAP from Apis cerana cerana and other species
2.3 GABARAP蛋白結構功能預測
經PrediceProtein在線預測分析獲得GABARAP基因編碼蛋白二級結構,GABARAP蛋白含有3個α-螺旋和4個β-折疊結構,以alpha-beta-beta-alpha-beta-alpha-beta的排列方式存在,具有6個多肽結合位點,沒有預測到蛋白含有二硫鍵或跨膜結構域(圖4)。
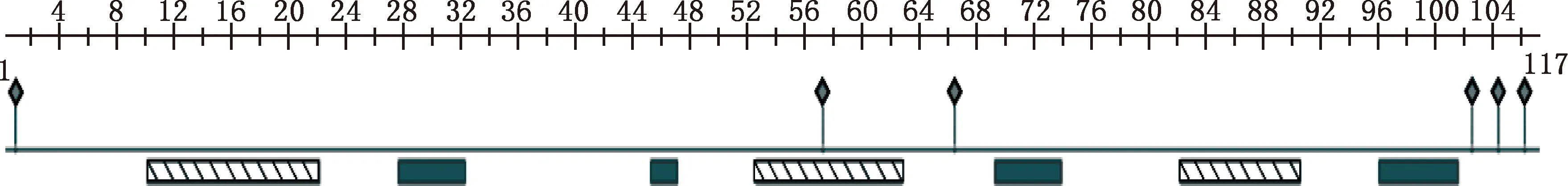
圖中條紋框代表α-螺旋;填充框代表β-折疊;菱形框代表多肽結合位點The striped frame indicates alpha-helices;The fill frame indicates beta-sheets;The diamond frame indicates polypeptide binding sites圖4 中華蜜蜂GABARAP蛋白二級結構預測Fig.4 Secondary structure prediction of GABARAP protein of Apis cerana cerana
利用SWISS-MODEL在線預測GABARAP蛋白三級結構。根據相似性選擇氨基酸序列一致性為85.47%的1gnu.1.A作為同源模擬的模板,進行GABARAP單體的同源建模。由圖5A可知,3個α-螺旋在圖示蛋白結構的外表面,4個β-折疊在圖示蛋白結構的內表面。預測到的中華蜜蜂GABARAP蛋白的三級結構與圖5B中真核生物的GABARAP的晶體結構(PDB:1GNU)極為類似,由此推測中華蜜蜂GABARAP蛋白可能執行與真核生物GABARAP蛋白相似的功能。
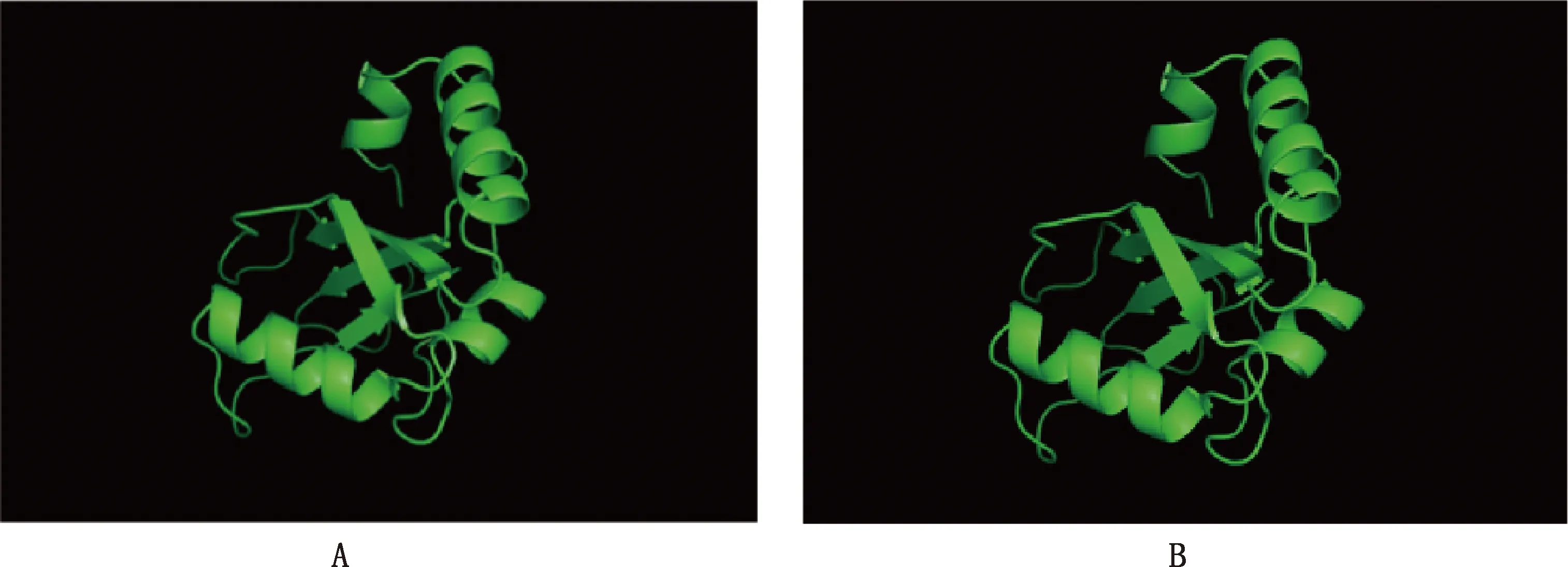
圖5 中華蜜蜂GABARAP蛋白單體三級結構同源模擬(A)和1GNU三維結構(B)比較Fig.5 Comparison of the tertiary structure homology modeling in Apis cerana cerana GABARAP monomer protein (A) and 1GNU three-dimensional structure (B)
2.4 GABARAP蛋白原核表達
SDS-PAGE結果顯示,誘導的pET-32a(+)-GABARAP重組質粒在35 ku處有一條特異蛋白條帶,與預期大小一致,而未誘導樣品中則無相應的條帶(圖6A)。利用His 標簽抗體進行Western blotting檢測進一步證明了35 ku的單一蛋白條帶為重組蛋白(圖6B)。
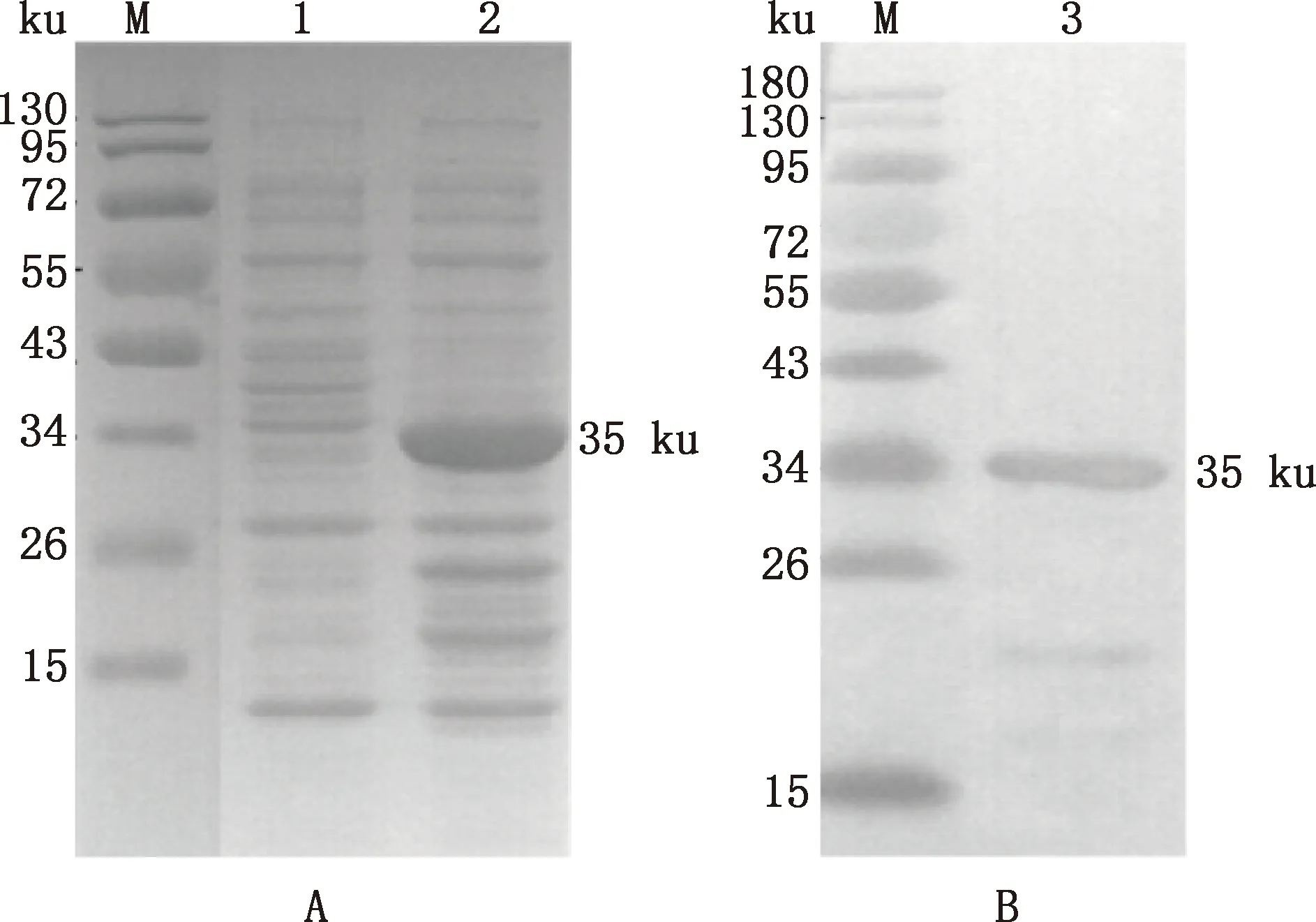
M,蛋白質分子質量標準;1,未誘導重組質粒;2、3,誘導后重組質粒M,Protein Marker;1,Uninduced recombinant plasmid;2 and 3,Induced recombinant plasmid圖6 GABARAP蛋白的誘導表達(A)和Western blotting檢測(B)Fig.6 Induced expression (A) and detection of Western blotting (B) of GABARAP protein
超聲處理過的誘導表達重組菌液,經Western blotting分析可知該重組蛋白的表達在包涵體中(圖7)。經His標簽蛋白純化試劑盒(包涵體蛋白)純化后,在獲得第3 mL穿流液時檢測到重組蛋白(圖8)。
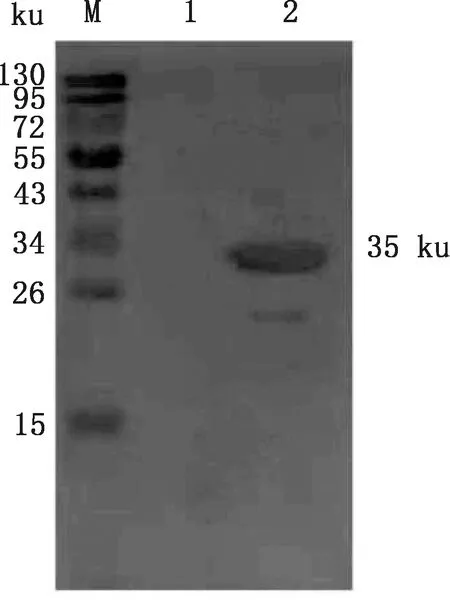
M,蛋白質分子質量標準;1,重組蛋白超聲后上清;2,重組蛋白超聲后沉淀M,Protein Marker;1,Supernatant of recombinant protein after ultrasonic processing;2,Precipitation of recombinant protein after ultrasonic processing圖7 GABARAP重組蛋白可溶性分析Fig.7 Soluble analysis of recombinant protein GABARAP
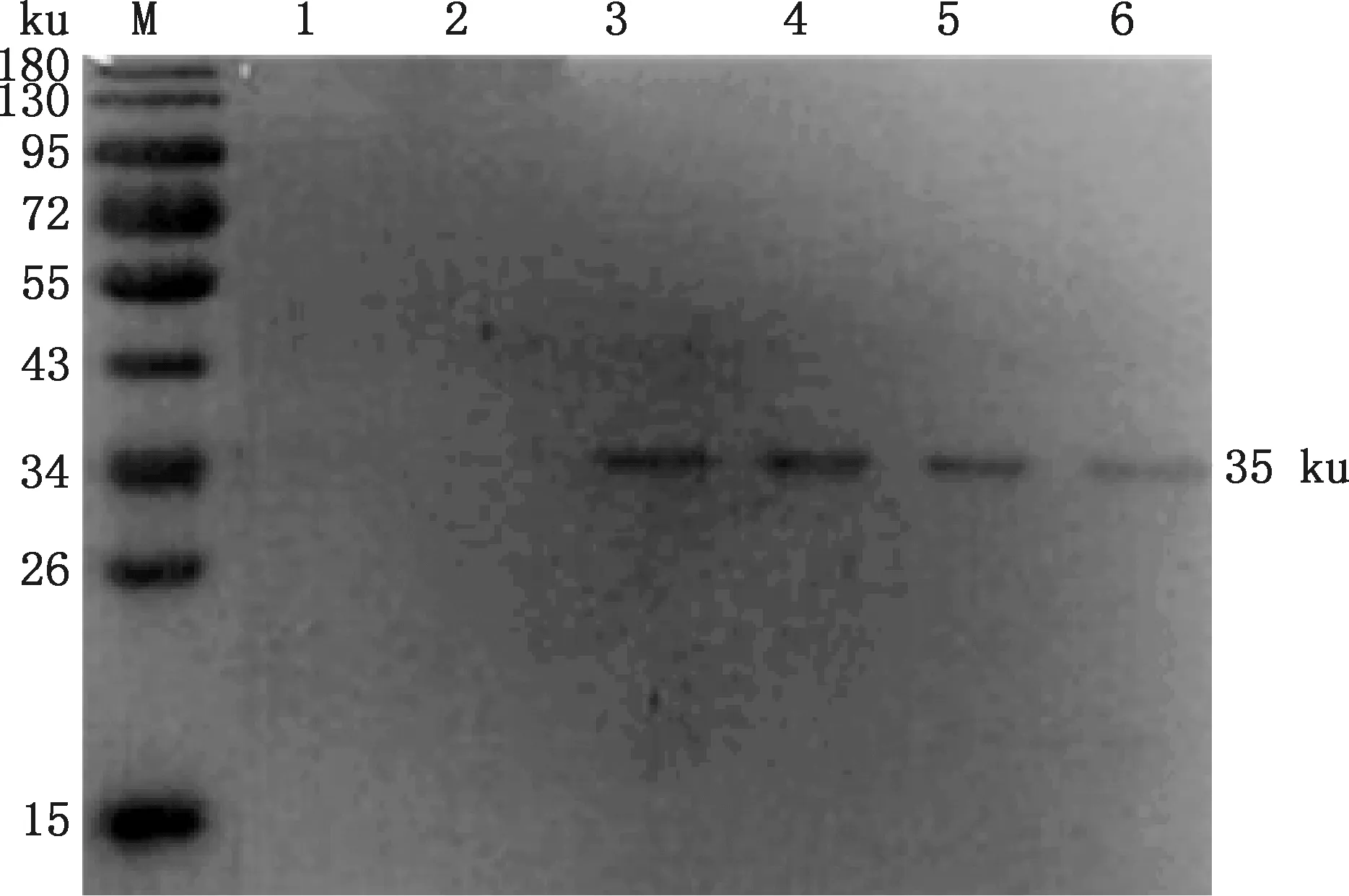
M,蛋白質分子質量標準;1~6,第1~6 mL流穿液M,Protein Marker;1-6,The 1-6 mL filtration圖8 重組蛋白GABARAP純化Fig.8 Purification of the recombinant protein GABARAP
3 討 論
細胞自噬廣泛存在于真核生物中,是一種高度保守的自我降解系統,可參與生物的生長發育、分化、炎癥反應及免疫調控等,在維持胞內環境穩定性方面發揮著重要作用[26]。自噬的發生與調節是由一系列的ATGs共同發揮作用。Atg8蛋白可通過調控自噬體膜的形狀和功能影響細胞自噬。GABARAP蛋白是Atg8家族的成員,是一種高度保守的蛋白,能與多種細胞因子互作,高度調控自噬體的運輸、形成與成熟等過程[4-6,27]。GABARAP能與上游調控因子MAPK15和ERK8發生相互作用,從而誘導自噬的起始[28-30]。Huber等[31]研究表明,GABARAP蛋白可通過不依賴脂化的方式調節泛素修飾蛋白活化酶5(UBA5)在內質網膜上的定位和功能。此外,在人類免疫缺陷病毒(HIV)感染早期輔助蛋白Nef(負調控因子)大量表達,該蛋白能通過干擾內吞和晚期分泌途徑中的蛋白質來調控在免疫和病毒生命周期中起重要作用的各種細胞表面受體的表達[32]。綜上,GABARAP蛋白與自噬體的發生、組織定位密切相關。
基于GABARAP功能的重要性,本研究克隆獲得了中華蜜蜂GABARAP基因序列,經氨基酸序列分析發現,該基因編碼117個氨基酸,蛋白分子質量為13.99 ku,研究結果與前期報道的人源GABARAP基因基本一致[33]。Bai等[34]研究表明,在雜色鮑中克隆GABARAP基因的cDNA全長得到963 bp,具有354 bp的開放閱讀框,推導的蛋白分子質量為13.9 ku,等電點為8.73,該結果與本研究中華蜜蜂GABARAP基因的克隆及推導結果幾乎完全相同,可見GABARAP基因在進化中高度保守,且該基因在應對病原體感染及胚胎發育中發揮著重要作用。GABARAP氨基酸序列比對和系統進化樹分析也表明了該基因的保守性。 人源GABARAP蛋白二級結構含有4個α-螺旋,而中華蜜蜂GABARAP蛋白僅預測到3個α-螺旋,在第4-8位的氨基酸沒有預測到α-螺旋,推測是此位置氨基酸的差異不易形成螺旋結構[8]。中華蜜蜂GABARAP三級結構預測結果與已發表的人源GABARAP蛋白的晶體結構(PDB∶1GNU)極為相似,Jatana等[35]研究表明,人源GABARAP蛋白高度參與自噬體的發生,從而調控細胞凋亡和免疫應答等。因此,推測中華蜜蜂GABARAP基因可能執行類似自噬調節相關的功能。GABARAP經原核表達并純化得到了35 ku的蛋白,Western blotting結果驗證了該蛋白的準確性,為后續中華蜜蜂GABARAP多克隆抗體的制備奠定了基礎。人源GABARAP與其3個家族成員(GABARAPL1、GABARAPL2和GABARAPL3)相似性很高,抗體檢測易與其家族成員產生交叉反應,對GABARAP抗體的進一步應用產生一定影響,西方蜜蜂Atg8家族成員僅包含GABARAP和LC3,因此不存在GABARAP抗體檢測過程中的內源性干擾[23,36-37]。GABARAP蛋白廣泛存在于多種組織中,在生物體的生長發育過程中不同程度地表達[12]。有研究證明GABARAP蛋白可以與自噬相關蛋白Atg13發生相互作用來調節自噬的發生[38-39],可見GABARAP蛋白是自噬的良好標記,在自噬過程中起著關鍵的調節作用。相關研究也發現家蠶感染核型多角體病毒(BmNPV)后可觸發自噬的發生,表現為Atg8基因的上調表達且免疫熒光也觀察到Atg8-GFP大量增加[40-41]。
4 結 論
本試驗克隆了中華蜜蜂GABARAP基因,其長度為354 bp,編碼117個氨基酸。氨基酸序列比對及進化樹分析表明GABARAP基因保守,二、三級結構預測該蛋白主要由3個α-螺旋、4個β-折疊和6個多肽結合位點組成;成功表達出GABARAP蛋白,分子質量為35 ku,經Western blotting驗證了GABARAP蛋白的特異性。本研究為進一步探索中華蜜蜂GABARAP蛋白的結構功能提供了基礎材料,為開展中華蜜蜂GABARAP蛋白參與自噬的研究奠定了基礎。
