泛耐藥鮑曼不動桿菌生物被膜形成能力與同源性分析*
2022-02-13 05:55:10孟千琳凌保東
醫藥導報 2022年2期
孟千琳,凌保東
(1.成都醫學院結構特異性小分子藥物研究四川省高校重點實驗室,成都 610500;2.成都醫學院藥學院,成都 610500)
近年來,泛耐藥鮑曼不動桿菌(extensively drug resistantAcinetobacterbaumannii,XDRAB)檢出率呈逐年上升趨勢,被稱為“21世紀革蘭陰性桿菌中的MRSA”[1],對全球公共衛生構成重大威脅。鮑曼不動桿菌的致病性與多種毒力因子有關,生物被膜作為重要的毒力因子,具有極強的黏附性、抗吞噬性、耐藥性,有助于鮑曼不動桿菌在醫院環境中的生存與轉移[2]。相關分子流行病學研究表明[3],鮑曼不動桿菌流行菌株具有不同的多位點序列分型(multilocus sequence typing,MLST),鮑曼不動桿菌克隆的基因組主要分為3個國際克隆組:國際克隆Ⅰ (IC1)、國際克隆Ⅱ(IC2)和國際克隆Ⅲ(IC3),其中IC2是歐洲和亞洲的主要克隆系。有研究表明[4],亞洲國家的耐碳青霉烯類鮑曼不動桿菌以IC2的克隆復合體(CC92)為主。此外,腸桿細菌基因間重復序列聚合酶鏈反應(ERIC-PCR) 常用于研究從不同標本中收集的鮑曼不動桿菌菌株之間的關系,有助于確定鮑曼不動桿菌與流行性克隆的傳播關系[5]。本研究通過MLST與ERIC-PCR對泛耐藥鮑曼不動桿菌進行同源性分析,同時檢測泛耐藥鮑曼不動桿菌的生物被膜形成能力,進一步掌握院內鮑曼不動桿菌流行現狀,以期為臨床防控泛耐藥鮑曼不動桿菌感染提供依據。
1 材料與方法
1.1材料
1.1.1菌株來源 37株XDRAB來自成都醫學院第一附屬醫院2018年1月—2020年7月患者,痰液20份,灌洗液9份,導管4份,傷口分泌物、膿液、尿、腦脊液各1份,其中臨床分布重癥醫學科15株,呼吸科重癥監護病區11株,外科6株,內科5株。質控菌金黃色葡萄球菌ATCC29213、大腸埃希菌ATCC25922、銅綠假單胞菌ATCC27853、大腸埃希菌DH5α為本實驗室保存。
1.1.2儀器與試劑 恒溫培養箱(上海一恒科技有限公司)、生物安全柜(Thermo Fisher Scientific)、酶標儀(SpectraMa190)、96孔細胞培養板(科茲莫生物科技有限公司)。胰酪大豆胨液體培養基(TSB, 青島海博生物技術有限公司,批號:20200815),結晶紫(成都金山化學試劑有限公司,批號:20170416)、磷酸鹽緩沖液(PBS,上海生工生物工程有限公司,批號:20201228)、聚合酶鏈反應(PCR)引物和測序(北京擎科生物科技有限公司)、2×Taq Master Mix (Dye Plus,南京諾唯贊生物科技股份有限公司)。
1.2方法
1.2.1XDRAB藥敏試驗 參照美國臨床實驗室標準化協會(CLSI2020)標準,金黃色葡萄球菌ATCC29213、大腸埃希菌ATCC25922、銅綠假單胞菌ATCC27853為質控菌株,采用微量肉湯稀釋法測定碳青霉烯類、β-內酰胺類、氨基苷類、喹諾酮類、多粘菌素類等16種抗菌藥物的最低抑菌濃度(MIC)值。
1.2.2XDRAB生物被膜形成能力檢測 菌液的制備:將細菌接種于LB固體培養板上,37 ℃,孵育16~20 h后,挑取單克隆于0.9%氯化鈉溶液200 μL中,吹打混勻,取100 μL,使用酶標儀測定600 nm波長處吸光度(A值),用0.9%氯化鈉溶液調整至A600值=0.1,即為麥氏0.5濃度,細菌含量達到1×108CFU·mL-1。結晶紫染色法(crystal violet staining,CVS):參照文獻[6-7]的實驗方法,并稍作調整,使用96孔細胞培養板,以大腸埃希菌DH5α為陰性對照組,菌株統一設置6個復孔,每孔加入A600值=0.1菌液20 μL和TSB培養基180 μL,37 ℃,孵育24 h。次日吸出培養基,用PBS 200 μL清洗3次,空氣干燥20 min,0.1%結晶紫染色20 min,吸出結晶紫,重復PBS清洗3次,空氣干燥20 min,加入95%乙醇溶解,酶標儀測定570 nm下A值。生物被膜形成能力判定標準為:①陰性:A模型對照≤A陰性對照;②弱陽性:A陰性對照
1.2.3MLST分型 鮑曼不動桿菌管家基因序列參照MLST數據庫Oxford方案(https://pubmlst.org/organisms/acinetobacter-baumannii),包括7個管家基因,分別為:gltA、gyrB、gdhB、recA、cpn60、gpi和rpoD。PCR反應體系:2 × Taq Master Mix 7.5 μL,上、下游引物各0.5 μL,DNA模板1 μL,ddH2O 5.5 μL,共15 μL。PCR反應條件為:94 ℃預變性5 min,94 ℃變性30 s,55 ℃退火30 s,72 ℃延伸1 min,共30次循環,72 ℃延伸5 min。擴增片段測序后與MLST數據庫進行比對獲得等位基因譜,按照gltA-gyrB-recA-cpn60-gpi-rpoD的排列順序輸入數據庫中獲得對應菌株的序列型(sequence type,ST)進行分析。
1.2.4ERIC-PCR分型 參照文獻[8]合成引物,PCR反應體系:2 × Taq Master Mix 25 μL,上、下游引物各2 μL,DNA模板10 μL,ddH2O 11 μL,共50 μL。PCR反應條件為:94 ℃預變性5 min,94 ℃變性30 s,40 ℃退火1 min,72 ℃延伸1 min,共35次循環,72 ℃延伸10 min。使用Image Lab 5.0軟件定量工具自動分析條帶數量進行同源性分析,參照文獻[9]比較菌株間相似性,以主條帶位置和數量相同定義為同一基因型。
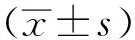
2 結果
2.1XDRAB對抗菌藥物的耐藥情況 37株XDRAB除對多粘菌素類、替加環素敏感外,其余抗菌藥物均耐藥,具體結果見表1。
2.2XDRAB培養24 h后生物被膜形成能力檢測 細菌培養24 h后,使用結晶紫染色法對生物被膜形成進行定量檢測,與陰性對照組DH5α比較,4株為強被膜形成菌株,7株為中被膜形成菌株,19株為弱被膜形成菌株,其余為陰性7株。
2.3MLST對XDRAB分型結果 經測序的37株XDRAB,按照ST分型和等位基因編號后結果顯示,37株XDRAB分為3種ST序列型,其中34株XDRAB為ST208型,2株為ST195型,1株為ST368型。ST208型菌株主要分離于呼吸科重癥監護病區(10/34)和重癥醫學科(14/34),分離標本主要來源于痰液(18/34)和灌洗液(8/34)。與全球序列數據庫進行比對,37株XDRAB均屬于國際克隆組IC2的克隆復合體(CC92),說明CC92為XDRAB感染流行的主要克隆復合體。
2.4ERIC-PCR對XDRAB擴增結果 37株XDRAB經ERIC-PCR擴增后,通過瓊脂糖凝膠電泳,形成2條左右的堿基條帶,條帶長度為250~500 bp,通過分析,菌株被分為I型(22株),II型(15株),共2種類型,結果見圖1。
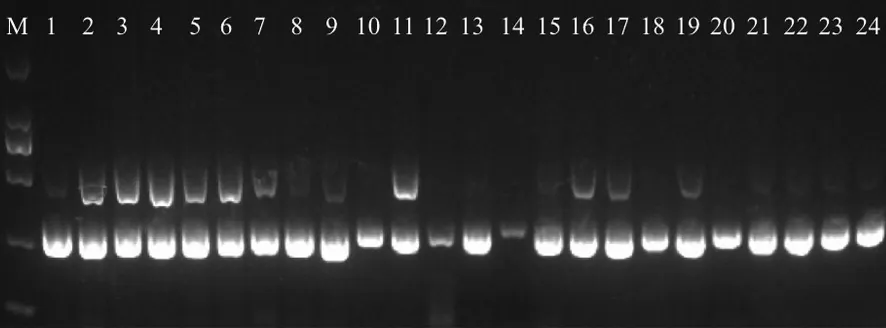
M:DNA2000Marker;1-9,11,13,15-17,19,21-24為I型;其余為Ⅱ型。圖1 部分XDRAB的ERIC-PCR圖譜M:DNA2000Marker; 1-9,11,13,15-17,19,21-24 are type I; the other are type Ⅱ.Fig.1 ERIC-PCR of partial XDRAB strains
3 討論
XDRAB生物被膜的形成使臨床面臨嚴峻挑戰[10],生物被膜形成過程主要分為四個階段:初始黏附、增殖、形成成熟生物被膜、生物被膜的分散再定植。生物被膜一旦形成,可通過滲透限制、營養限制、基因表達上調、免疫逃逸等耐藥機制為鮑曼不動桿菌提供保護模式并增強對抗菌藥物的抵抗力[11-12]。本研究中37株XDRAB經檢測有30株具有生物被膜形成能力,81%以上XDRAB均會形成生物被膜,是導致醫院慢性感染和難治性感染的反復發作的重要原因。
為避免XDRAB在院內感染繼續擴散,選擇有效的準確的菌株同源性分析方法至關重要。MLST作為分子流行病學和菌株種群遺傳分析的通用方法[13],通過分析細菌7個管家基因的等位基因內部片段的DNA序列,清晰呈現每個分離株的序列類型。分離株的遺傳相關性可以通過等位基因圖譜之間的差異矩陣以樹狀圖的形式直觀呈現[14]。ERIC-PCR指紋圖譜是最有效區分鮑曼不動桿菌的分子分型技術之一[15],包含重復序列(如:ERIC序列)的細菌基因組可通過該序列快速評估細菌分離株的克隆變異性。本研究采用MLST和ERIC-PCR兩種國際通用分子分型技術對37株XDRAB進行同源性分析,MLST結果顯示,ST208型為主要分型,其次為ST195和ST368,均屬于CC92,該結果與KIM等[16]研究結果一致,此外,QU等[17]研究表明,ST208型已成為中國西部地區鮑曼不動桿菌感染暴發的主要序列型。ERIC-PCR結果顯示,37株XDRAB分為Ⅰ型(22株)、Ⅱ型(15株),由于ERIC基因為保守序列,其擴增產物無法完全確定,可能會出現部分電泳條帶缺失、模糊等不確定因素[18],因此,本實驗選擇采用兩種分子分型技術對XDRAB進行同源性分析,以保證更全面地呈現XDRAB的流行病學趨勢。
目前,關于XDRAB的流行病學與毒力因子相關性分析的研究很少。生物被膜作為重要的毒力因子[19],其形成的分子機制和基因主要包括Csu基因座(CsuAB/A/B/C/D/ E)參與菌毛合成,Pga基因座參與編碼多聚-N-乙酰基葡糖胺(PNAG),OmpA參與編碼外膜蛋白,Bap參與編碼生物被膜相關蛋白[20]。相關研究表明[21],ST208攜帶主要生物被膜形成基因,其中97.7%存在Csu基因,ST195對Pga和OmpA基因陽性率高。由于本研究菌株ST分型主要以ST208為主,所以無法深入分析生物被膜形成能力與ST分型的相關性,對于XDRAB流行病學與毒力因子相關性分析,有待進一步研究。
綜上所述,XDRAB導致的院內感染的治療作為全球公共衛生的重大難題,亟待制定出可控治療方案,本研究采用目前國際常用分子分型技術MLST與ERIC-PCR對XDRAB進行流行病學分析,針對ST208菌株需要嚴格注意感染路徑,避免交叉感染,盡可能降低臨床感染風險。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
保健醫苑(2022年5期)2022-06-10 07:46:38
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
航空世界(2020年10期)2020-01-19 14:36:20
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
中國當代醫藥(2015年17期)2015-03-01 02:03:58
