電針“委中”穴調節PGC-1α及相關影響因子對腰多裂肌損傷大鼠線粒體功能的影響
2022-02-11 12:30:02李霞呂巧巧徐菁陳莉田圓張莉劉通
環球中醫藥 2022年11期
李霞 呂巧巧 徐菁 陳莉 田圓 張莉 劉通
腰多裂肌是從腰背部一直跨越至骶部的肌肉,其在維持腰椎穩定中提供70%力量[1],也是腰椎術后疼痛綜合征發病的重要原因之一[2-3]。因此,恢復腰多裂肌結構和功能至關重要。腰多裂肌屬于骨骼肌,骨骼肌損傷修復的機制與線粒體功能的變化緊密相關。線粒體功能正常一方面可減少活性氧(reactive oxygen species,ROS)等氧化應激對骨骼肌的傷害[4];另一方面,促進肌衛星細胞成肌分化,幫助骨骼肌再生[5]。過氧化物酶體增殖物活化受體γ共刺激因子-1α(peroxisome proliferator-activated receptor γ coactivator-1α,PGC-1α)主要調節線粒體生物合成和抗氧化活性,在線粒體有氧磷酸化過程中發揮重要作用[6]。大量研究發現,PGC-1α受Ca2+信號通路調控[7]。鈣/鈣調蛋白依賴性蛋白激酶(Ca2+/calmodulin dependent protein kinaseⅡ,CaMKⅡ)可被Ca2+和鈣調蛋白激活,導致自身磷酸化,磷酸化CaMKⅡ可活化PGC-1α并促進其表達,提高線粒體生物發生水平[8]。不僅如此,激活的CaMKⅡ還可直接活化cAMP反應元件結合蛋白(cAMP-response element binding protein,CREB),CREB磷酸化后可啟動PGC-1α基因表達,上調PGC-1α蛋白水平[9]。因此,鈣/鈣調蛋白依賴性蛋白激酶、CREB與PGC-1α對線粒體功能的影響具有重要意義,其可能是促進損傷骨骼肌線粒體功能恢復的機制。
課題組既往研究通過注射布比卡因制備大鼠多裂肌損傷模型,以電針“委中”為干預手段,電鏡觀察發現肌漿網腫脹,線粒體大小不一、結構異常,而針刺能夠改善這一現象并促進多裂肌的損傷修復[10]。本研究擬通過多時間點觀察,電針“委中”是否通過調節PGC-1α及相關影響因子的表達,減少線粒體功能損害,促進骨骼肌的損傷修復,從而闡釋電針修復骨骼肌損傷的部分機制。
1 材料與方法
1.1 實驗動物
健康雄性SD大鼠72只,體質量200~250 g,由北京斯貝福實驗動物中心提供[許可證號:SYXK(京)2019~0010]。在北京中醫藥大學動物房(清潔級)喂養,自然照明,自由攝食、飲水。整個實驗過程中對動物的各種處理均遵照中華人民共和國科技部2006年頒布的有關動物的使用及倫理學規定。
1.2 實驗分組
將動物隨機分為正常組、模型組、電針組,每組24只。每組再根據干預天數隨機分為1天、2天、3天和7天,4個亞組。其中左側組織置于多聚甲醛浸泡,用于蘇木精—伊紅染色(hematoxylin-eosin staining,HE),右側組織迅速置于液氮中,進蛋白免疫印跡(Western blot,WB)、聚合酶鏈式反應(polymerase chain reaction,PCR)生化法檢測。
1.3 主要儀器和試劑
HANS-100 A電針儀,南京濟生醫療科技有限公司;倒置顯微鏡及成像系統,日本尼康;全自動化學發光圖像分析系統,美國Bio-Rad;酶標儀,M2型,芬蘭Labsystems Multiskan M2;布比卡因鹽酸鹽,美國Sigma;HE染色試劑盒(G1120,北京索萊寶生物技術有限公司);TOMM20抗體(11802-1-AP,proteintech);鈣離子測試盒(C004-2-1,南京建成生物工程研究所);SOD測試盒(A001-3,南京建成生物工程研究所);超微量總ATP酶測試盒(A070-1-2,南京建成生物工程研究所);RNA提取試劑盒(R4115,Magen);逆轉錄試劑盒(K1622,Thermo);熒光染料(A25472,Thermo)。
1.4 大鼠腰多裂肌損傷模型的制備
大鼠稱重后用20%烏拉坦(0.5 mL/100 g)腹腔注射麻醉。腰背部備皮,操作過程保持無菌。在雙側脊椎L4、L5水平的多裂肌注射布比卡因溶液(100 μL,0.5%),使用一次性4號針頭注射器抽取布比卡因溶液,針頭緊貼棘突旁進入肌肉,直到接觸關節突和乳突所在的骨面回抽套管1 mm,無血,表明針頭已到達多裂肌后開始注射,注射時間不得小于3秒,以利于藥物的吸收。共有四個注射點(L4、L5水平旁邊各兩點),共計400 μL(100 μL×4)。單次注射造模完成。通過HE染色進行模型評價,以鏡下見炎性細胞浸潤,肌肉組織間隙增大,肌纖維斷裂為造模成功。
1.5 電針治療方法
將大鼠固定在特制的固定器上,暴露后肢。采用華佗牌0.30 mm×13 mm一次性針灸針,針刺后連接韓式電針儀HANS-100 A,2/15 Hz的疏密波,電流強度2 mA,持續30分鐘,1次/天。“委中”穴定位參照《實驗針灸學》常用動物針灸穴位及圖譜,定位于膝關節正后方凹陷中。模型組同時也在固定器中固定,但不做針刺干預。
1.6 檢測指標及方法
1.6.1 HE染色 左側多裂肌4%多聚甲醛固定24小時,常規石蠟包埋,切片厚度5 μm,置于37℃烘箱一夜,將切片置于二甲苯脫蠟2次,各5分鐘;梯度酒精(100%、95%、80%、70%)各2分鐘,蒸餾水2分鐘,蘇木素10分鐘,自來水沖洗1分鐘,分化液10秒,自來水浸泡15分鐘,伊紅50秒,自來水沖洗1分鐘,梯度脫水(95%、95%、100%、100%)各2秒,二甲苯透明2次各1分鐘,中性樹脂封片。在顯微鏡下進行觀察拍照。
1.6.2 Western blot進行TOMM20檢測 從-80℃冰箱取出凍存組織,稱取適量的肌肉,按照1 mg組織加入6 μL RIPA裂解液,在研磨機上進行充分研磨,裂解完成后離心,收集上清,通過BCA試劑盒進行蛋白定量,剩余上清加入5× loading buffer,95℃沸水煮5分鐘后,置于-20℃冰箱,用于后續上樣。制備5%濃縮膠和10%分離膠,每孔上樣5 μL,濃縮膠60V 15分鐘,分離膠80V 90分鐘。跑膠完成后以400 mA進行電轉,轉膜結束后用10%的牛奶封閉2小時,敷一抗,放置-4℃冰箱過夜。室溫回溫10分鐘后,TBST洗膜3遍,二抗室溫孵育2小時,TBST洗膜3遍,在全自動化學發光圖像分析系統滴加超敏ECL曝光。最終通過Image-lab進行條帶分析。
1.6.3 PCR運用PCR檢測PGC-1α mRNA、CaMKⅡ mRNA和CREB mRNA,稱取20 mg新鮮組織,按照制造商的說明用RNA提取試劑盒提取RNA,并通過核酸紫外光譜法測定RNA濃度。根據試劑盒制造商的說明,在20 μL系統中將RNA反向轉錄成cDNA。引物序列及長度見表1。

表1 引物序列及產物大小
1.6.4 生化法 稱取組織,按重量(g):體積(mL)=1∶9的比例,加入9倍體積的生理鹽水,冰水浴條件下機械勻漿,3500轉/分,離心10分鐘,取上清液進行待測。按照Ca2+、SOD、ATP酶試劑盒說明書進行檢測。為了消除樣品制備時由于蛋白量的差異而造成的誤差,用BCA試劑盒進行蛋白濃度測定試劑盒測定樣品種的蛋白濃度,然后把濃度換算成nmol/mg蛋白的形式。
1.7 統計學方法
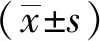
2 結果
2.1 HE染色觀察形態學變化
通過HE染色觀察1天、2天、3天、7天,正常組、模型組和電針組的形態學變化。發現正常組肌纖維排列緊密整齊,細胞核均勻排布且靠近肌膜,未見炎性細胞浸潤。干預1 天后,模型組肌纖維間隙增大,間隙中分布炎性細胞浸潤,肌纖維開始壞死斷裂;電針組較模型組肌間隙縮短,炎性細胞浸潤較少。干預2天后,模型組肌纖維大片壞死、斷裂,肌細胞間隙增寬,大量炎性細胞浸潤;電針組肌間隙內炎性細胞浸潤較模型組少。干預3天,模型組與電針委中組仍可見較多新生的中央核纖維生成,但模型組仍可見較多炎性細胞浸潤,而電針委中組的炎性細胞較少。干預7天,模型組仍可見炎性細胞與大量新生肌纖維,肌纖維細胞核位于中央,肌間隙變小,而電針委中組較多的新生肌纖維的細胞核已經逐漸遷移至細胞邊緣,肌間隙縮小,炎性細胞浸潤少。見圖1。
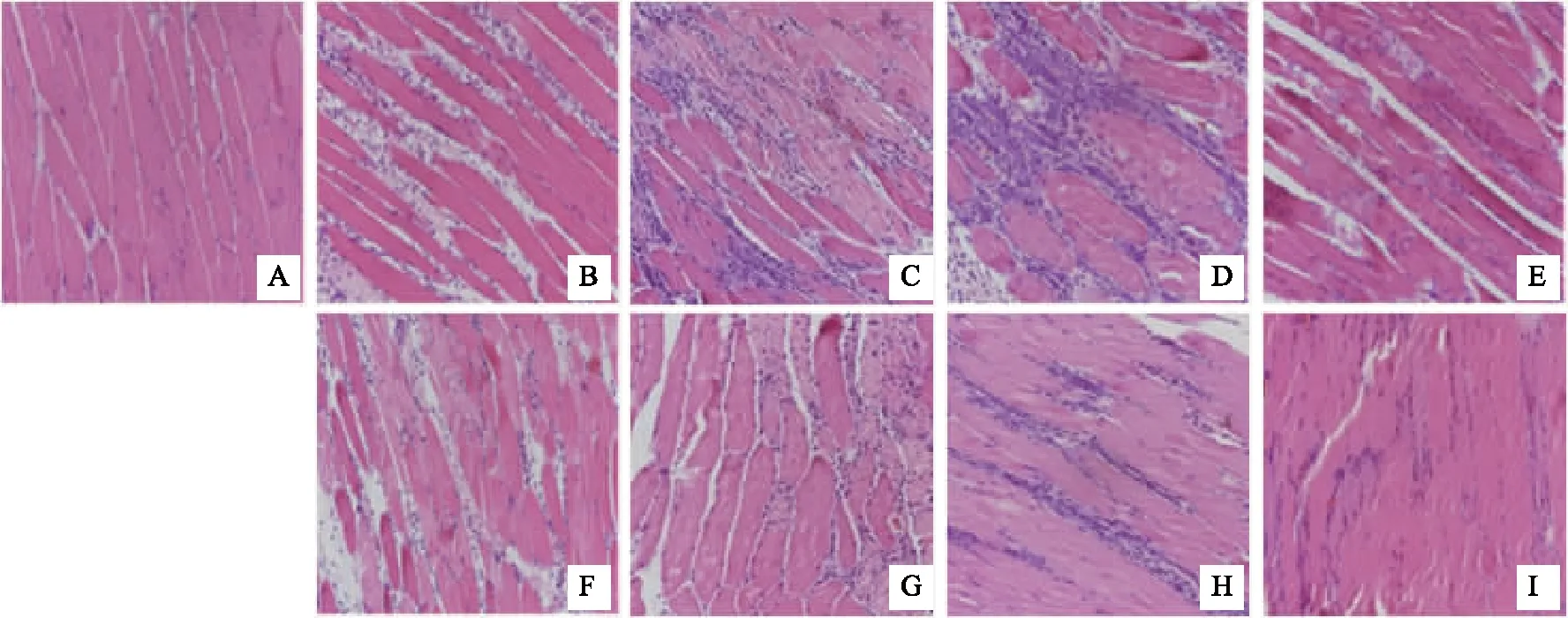
注:A 正常組;B-E 模型組亞組:1天、2天、3天、7天;F-I 電針組亞組:1天、2天、3天、7天。
2.2 多裂肌Ca2+、CaMKⅡ mRNA、CREB mRNA表達含量
為評價電針后多裂肌損傷后Ca2+的含量的變化,課題組進行了多時間點多裂肌Ca2+含量檢測。與同一時間點正常組相比,模型組2天、3天亞組Ca2+含量升高(P均<0.05);與同一時間點模型組相比,電針組2天、3天亞組Ca2+含量降低(P<0.01,P<0.05);同一時間點,模型組和電針組1天、7天亞組均高于正常組(P<0.01,P<0.05)。與同一時間點模型組相比,電針組1天、7天亞組無顯著差異。見表2。
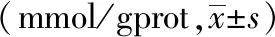
表2 不同時間點各組大鼠多裂肌Ca2+含量的變化比較
為評價電針對多裂肌損傷后CaMKⅡ mRNA表達的變化,課題組進行了各時間點多裂肌CaMKⅡ mRNA含量的檢測。與同一時間點正常組比較,模型組和電針組2天、3天、7天亞組CaMKⅡ mRNA表達高于正常組(P<0.05);與同一時間點模型組比較,電針組2天、3天、7天亞組含量明顯低于模型組(P<0.01)。1天時,三組無明顯差異(P>0.05)。見表3。
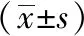
表3 不同時間點各組大鼠多裂肌CaMKⅡ mRNA表達含量的變化比較
為評價電針對多裂肌損傷后CREB mRNA表達的變化,同樣進行了各時間點多裂肌CREB mRNA含量的檢測。與同一時間點正常組比較,模型組和電針組1天、2天、3天、7天亞組均高于正常組(P<0.01);與同一時間點模型組比較,電針組2天、3天、7天亞組均高于模型組(P<0.01)。1天時則無統計學差異(P>0.05)。見表4。
表4 不同時間點各組大鼠多裂肌CREB mRNA表達含量的變化比較
2.3 PGC-1αmRNA表達含量
為評價電針對多裂肌損傷后PGC-1α mRNA含量表達的變化,進行了各時間點PGC-1α mRNA含量的檢測。與同一時間點正常組比較,模型組和電針組2天、3天、7天亞組均高于正常組(P<0.01);與同一時間點的模型組相比,電針組2天、3天亞組含量明顯低于模型組(P<0.05),7天時,電針組和模型組比較無差異(P>0.05),1天時,三組含量表達無統計學差異(P>0.05)。見表5。
表5 不同時間點各組大鼠多裂肌PGC-1α mRNA表達含量變化比較
2.4 線粒體功能指標TOMM20、SOD與ATP酶表達情況
為檢測多裂肌損傷后線粒體功能我們進行了TOMM20含量的檢測。與同一時間點正常組比較,1天、2天、3天、7天時電針組與模型組均低于正常組(P<0.01);與同一時間點模型組比較,2天時電針組高于模型組(P<0.05);1天、3天、7天時電針組與模型組無統計學差異(P>0.05)。見圖2、表6。
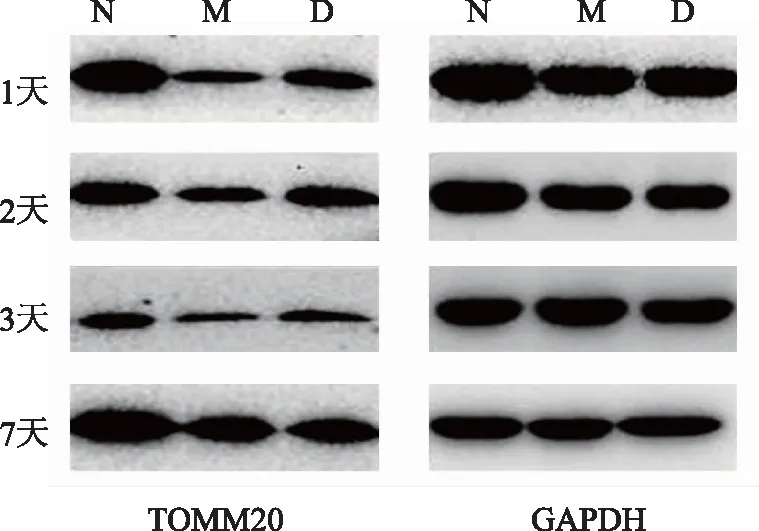
注:N 正常組;M模型組;D 電針組。
表6 不同時間點各組大鼠多裂肌TOMM20蛋白含量的變化比較
為檢測多裂肌損傷后線粒體功能進行了SOD含量的檢測。發現與正常組、模型組和電針組在1天、7天時SOD含量表達無統計學差異(P>0.05);2天時模型組含量明顯低于正常組,但是電針組明顯高于正常組(P<0.05);與同一時間點模型組相比,電針組含量明顯高于模型組(P<0.01)。3天時模型組明顯低于正常組(P<0.05),電針組與正常組無統計學差異(P>0.05)。與同一時間點模型組相比,電針組明顯高于模型組(P<0.01)。見表7。
表7 不同時間點各組大鼠多裂肌內SOD含量表達的變化比較
為檢測多裂肌損傷后線粒體功能進行了ATP酶含量的檢測。與同一時間點正常組比較,1天、2天、3天時模型組低于正常組(P<0.01),但7天時無顯著差異(P>0.05);2天、3天時電針組低于正常組(P<0.05,P<0.01),1天、7天時無顯著差異(P>0.05)。與同一時間點模型組相比,1天、3天時電針組含量明顯高于模型組(P<0.05),2天、7天時無統計學差異(P>0.05)。見表8。
表8 不同時間點各組大鼠多裂肌ATP酶含量的變化比較
3 討論
“腰背委中求”針刺委中治療腰背痛已廣泛在臨床應用[11]。Dar等[12]人研究發現針刺可顯著改善腰多裂肌的功能,有課題組前期運用布比卡因注射進行多裂肌損傷大鼠模型,以電針“委中”為干預手段,通過1天、3天、7天多時間點進行多裂肌損傷大鼠的觀察,發現多裂肌的損傷修復存在時間關聯[13],并且發現針刺可以增加肌源性轉錄因子(paired box7,Pax-7)、成肌分化抗原(myogenic differentiation antigen,MyoD)和磷酸化蛋白激酶B(phosphorylated protein kinase B,P-AKT)蛋白表達的影響促進肌衛星細胞的增殖,且有促進成肌分化的趨勢[14]。線粒體作為細胞的能量提供者,與肌肉能量代謝關系緊密,線粒體功能正常則促進衛星細胞成肌分化[5],而線粒體異常將抑制成肌分化。因此,促進線粒體功能的恢復,是促進成肌分化,幫助骨骼肌損傷修復的第一步。文獻研究顯示骨骼肌損傷2天時,線粒體功能影響最嚴重,這與先前的研究存在關聯。因此,在本研究中,課題組通過布比卡因注射液誘導的大鼠多裂肌損傷模型,設置1天、2天、3天、7天的四個時間點進行觀察,著力于研究對腰多裂肌損傷大鼠線粒體功能改變的相關機制,探討電針的起效的具體機制。
HE染色結果發現,模型組肌纖維破壞,肌間隙增寬,炎性細胞浸潤提示造模成功。干預后電針加速了新生肌纖維的融合,減少炎性細胞的浸潤,幫助多裂肌的恢復。PGC-1α是調節線粒體生物合成的關鍵性信號分子,可以通過活化線粒體功能、提高骨骼肌氧化代謝能力[15]。當骨骼肌受到缺血、缺氧、低溫、收縮及運動等刺激下, 可導致其表達增加, 進而啟動線粒體DNA復制和轉錄而誘導線粒體生物合成[16]。因此,通過PCR進行PGC-1α mRNA表達變化的檢測,發現造模后電針組與模型組損傷多裂肌內PGC-1α均較正常組增加,并且在模型組3天含量表達最多,而電針組則在2天含量表達最多,并且電針組的最高表達量明顯低于模型組。張靜等[17]發現PGC-1α mRNA的增加,可以顯著提高SOD與ATP酶活性,以此幫助細胞改善線粒體能量代謝和減輕氧化應激。TOMM20已被證實是線粒體膜復合物易位酶的一個重要亞基,負責識別線粒體蛋白并將其從細胞溶質轉移到線粒體中[17],降低提示線粒體丟失,可以反映線粒體的相對數量[18]。為進一步了解PGC-1α含量的增加對線粒體功能的影響,課題組通過檢測TOMM20、ATP酶含量以及SOD的含量變化。結果顯示,在造模后TOMM20、ATP酶、SOD的含量均減少,與同一時間點比較,電針能提高線粒體功能。值得注意的是模型組3天 PGC-1α含量表達最多,但線粒體功能反而受到抑制[19]。似乎PGC-1α含量的在一定范圍內的增加會增加線粒體功能,但是PGC-1α的過表達會抑制線粒體功能。劉長華[20]研究認為,適當的PGC-1α表達增加可以改善心臟收縮功能、促進心肌能量代謝、改善心力衰竭預后,但是在PGC-1α超表達的轉基因小鼠體內卻發現該類動物心肌細胞過度肥大、線粒體過度增殖、三羧酸循環相關的酶系大量表達,從而引起心臟過度的肥厚及擴張。研究發現,成年小鼠PGC-1α過表達會造成線粒體生物合成過度激活,線粒體結構排列紊亂和心肌病的發生[20]。因此,應抑制PGC-1α表達的過度激活,而PGC-1α的適當增加會促進線粒體功能。本研究電針似乎通過減輕了PGC-1α的過度的激活,促進線粒體功能的恢復。
CaMKⅡ的磷酸化對PGC-1α的激活有至關重要的影響。當骨骼肌損傷,內質網應激釋放大量的Ca2+,細胞內鈣離子濃度突然升高,CaMKⅡ會發生快速磷酸化作用,此時CaMKⅡ被激活, 這種激活方式也會使CaMKⅡ產生最大活性[21]。激活的CaMKⅡ可直接活化CREB, CREB磷酸化后可與PGC-1α mRNA上游啟動子區中的CRE序列結合,從而啟動PGC-1α基因表達,上調PGC-1α蛋白水平[7]。CREB對PGC-1α調控障礙是導致線粒體損傷的重要病理事件。LIU等[22]發現布比卡因誘導的SH-SY5Y細胞氧化損傷中CREB磷酸化升高,用siCaMK2α或KN93抑制CaMKⅡα活性可降低CREB磷酸化。此外,Wrighet等[23]發現咖啡因激活CaMK后,PGC-1α mRNA表達顯著升高,且CaMK抑制劑KN93抑制了這種增加,提示CaMK同樣參與調控PGC-1α mRNA的表達。因此,CaMKⅡ直接或間接的影響PGC-1α的表達。課題組檢測各時間點多裂肌內的Ca2+、CaMKⅡ mRNA和CREB mRNA,發現造模后模型組Ca2+、CaMKⅡ mRNA和CREB mRNA在3天達到表達高峰,而電針組Ca2+、CaMKⅡ mRNA的表達在2天已經達到高峰,CREB mRNA在3天達到表達高峰。值得注意的是電針組Ca2+、CaMKⅡ mRNA和CREB mRNA表達最高量明顯低于模型組的最高表達量。這與PGC-1α變化趨勢一致。因此,課題組大膽推測,由于造模后損傷多裂肌Ca2+、CaMKⅡ mRNA和CREB mRNA的過度表達,導致PGC-1α的過度激活,抑制了線粒體功能,電針通過抑制降低Ca2+、CaMKⅡ mRNA和CREB mRNA的表達,減輕對PGC-1α的過度激活,保護線粒體功能,進而促進骨骼肌的損傷修復。
綜上,課題組通過復制腰多裂肌損傷模型,設置1天、2天、3天、7天四個時間點,檢測PGC-1α以及PGC-1α相關因子,評估電針對損傷多裂肌線粒體功能的影響,發現電針可以減輕PGC-1α過度激活。較模型組而言,電針似乎加快了線粒體功能的恢復進程,促進骨骼肌快速修復。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中華詩詞(2022年6期)2022-12-31 06:41:24
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
中國科技論壇(2017年7期)2017-07-25 08:49:53
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
