植物香豆素生物合成途徑及關鍵酶基因研究進展
2022-02-10 11:19:14段珍吳凡閆啟張吉宇
草業學報 2022年1期
段珍,吳凡,閆啟,張吉宇
(蘭州大學草地農業生態系統國家重點實驗室,蘭州大學草地農業科技學院,甘肅 蘭州 730020)
香豆素類化合物是一類具有芳香氣味的天然產物,是通過苯丙氨酸和部分植物中的酪氨酸從莽草酸途徑中衍生出來的苯丙烷內酯。已從植物、細菌和真菌次生代謝產物中鑒定出超過1300種香豆素。香豆素最初是在豆科植物零陵香豆(Dipteryx odorata)中發現的,目前在蕓香科、豆科、傘形科等74科、330多個屬的植物中發現了香豆素類化合物[1—2]。在自然界中發現的香豆素有多種類型,既具有抗炎、抗病毒[3]、抗菌[4]、保護心臟[5]、誘導癌細胞凋亡[6]等藥理作用,在植物中還可作為主要的化感物質用作天然除草劑[7],另外,還影響多種植物的生長和發育,特別是種子萌發[8]、調節根系微生物群落結構[9]、抗氧化[10]和對逆境脅迫的響應[11]等。
盡管香豆素對人類和植物具有重要作用,但其生物合成的主要細節仍在不斷探索中。本研究綜述了香豆素在植物體內的發生與功能,以及生物合成途徑的主要步驟,重點介紹了所涉及的酶,討論了近年來發現香豆素合成相關酶(基因)的進展,可為香豆素的后續研究提供參考和理論支撐。
1 香豆素概述
1.1 分類
香豆素類化合物通過苯丙烷途徑產生,以游離態或糖苷的形式存在于植物中,具有極性結構,可分為簡單香豆素和復雜香豆素。一般結構簡單的化合物如香豆素、東莨菪內酯、傘形酮內酯等廣泛存在于菊科、豆科、傘形科和禾本科等多種植物中;而一些結構復雜的化合物如補骨脂素、花椒樹皮素等可在傘形科、蕓香科和豆科植物中見到。復雜香豆素以苯并α—吡喃酮為母核,通過在簡單香豆素核心苯環上添加雜環化合物而產生,并進一步分為呋喃香豆素、吡喃香豆素、苯并香豆素、二氫呋喃香豆素和雙香豆素[12]。簡單香豆素是指僅僅在苯環上有取代,且7位羥基與6位或者8位沒有形成呋喃環或者吡喃環的香豆素,取代基包括羥基、甲氧基、亞甲二氧基和異戊烯基等。由于香豆素吸收紫外光,因此具有特有的藍色熒光[13],香豆素母體本身無熒光,羥基化香豆素在紫外光下大多顯藍色熒光。
1.2 分布與含量
香豆素主要存在于植物根[14—19]、莖[20]、葉[21—25]、花[26—30]、果皮[31—33]、種子[34—35]、果實[36]以及根系分泌物中[37],其含量與植物體的利用密切相關,不同植物、不同品種、不同種質、不同部位香豆素含量存在差異。例如在豆科草木樨屬(Melilotus)中,細齒草木樨(Melilotus dentatus)香豆素含量較低,為0.1%(干物質含量),黃香草木樨(Melilotus suaveolens)香豆素含量較高,為0.753%(干物質含量)[38];黃花草木樨(Melilotus officinalis)種質間香豆素含量為0.3%~1.5%(干物質含量),白花草木樨(Melilotus albus)種質間香豆素含量為0.2%~1.3%(干物質含量)[39];朱宏等[40]的研究發現草木樨葉、花中香豆素平均含量為6.64 mg·g—1,莖中香豆素平均含量為6.18 mg·g—1,全草香豆素含量為6.30 mg·g—1;湯春妮[41]發現草木樨富含香豆素的最佳時間是盛花期。而在傘形科植物白花前胡(Peucedanum praeruptorum)的干燥根中總香豆素提取物中白花前胡甲素含量為602.42~721.44 mg·g—1,白花前胡乙素的含量為38.34~101.46 mg·g—1[42]。蕓香科柑橘屬(Citrus)柚子(Citrusmaxima)皮中檢測到7-甲氧基香豆素含量較高,為11.4~123.7 mg·g—1[43]。在傘形科植物珊瑚菜(Glehnia littoralis)中,不同采收期根和葉的香豆素總量不同,根中在不同采收期的香豆素總含量為0.0466~0.0772 mg·g—1,葉中不同采收期的香豆素總含量為0.0131~0.2065 mg·g—1[44]。
1.3 功能
香豆素是最簡單的植物次生代謝物之一,在植物中分布廣泛,在細胞受損傷后釋放,其功能大多和防御有關,例如抗微生物、紫外線屏蔽、抑制種子萌發等[45]。
1.3.1香豆素可參與植物的生長和生理過程 香豆素可以通過抑制水稻(Oryza sativa)脫落酸分解代謝和胚活性氧的積累來延遲種子萌發[8];能誘導α-淀粉酶的產生,抑制超氧化物歧化酶、過氧化氫酶和抗壞血酸過氧化物酶的活性,降低細胞活力,破壞小麥(Triticum aestivum)糊粉層氧化還原的穩態[46];可以通過調節質膜H+-ATP酶活性來提高玉米(Zea mays)根系對硝酸鹽的吸收,且發現它可能在基因轉錄中起誘導因子的作用[47];香豆素還是黃花草木樨的主要化感物質,對許多雜草種子萌發和幼苗生長有較強的抑制作用,可作為天然除草劑[7]。
1.3.2東莨菪內酯等羥基化香豆素參與植物對脅迫的響應 Chong等[48]的研究發現東莨菪內酯具有清除活性氧自由基的作用,可能參與減輕植物細胞氧化應激反應。在擬南芥(Arabid opsis thaliana)中,東莨菪內酯和生長素刺激了細胞結構的發育[49],低溫顯著誘導了東莨菪內酯的積累[50]。另外植物在缺鐵條件下根系會分泌東莨菪內酯、秦皮素和兒茶酚香豆素、七葉亭、瑞香素等小分子氧化還原活性分子至根際以促進鐵的活化和吸收,兒茶酚香豆素可與土壤中的鐵形成鐵螯合物或還原Fe3+,東莨菪內酯因不含兒茶酚基不能被Fe3+所吸附[51]。另外,Chutia等[52]發現香豆素的組成變化還依賴于鐵和磷。
此外,東莨菪內酯被分泌到根際會選擇性地影響擬南芥根系的菌群群落組成,抑制植物病原體的生長,調節根系菌群,促進植物健康生長[9]。Beyer等[53]發現擬南芥東莨菪內酯可以抑制亞洲銹病真菌的活性以保護大豆(Glycine max)免受感染,可以作為一種天然殺菌劑。抗菌素東莨菪內酯和秦皮素在煙草(Nicotiana attenuata)根中積累,以抵御壞死性真菌的感染[54]。從苦楝(Melia azedarach)果實中分離的東莨菪內酯與香草醛等其他化合物聯用,具有協同抗真菌作用[36]。說明植物可以利用香豆素等代謝產物的混合物從根際消滅致病菌。
1.3.3香豆素與牧草品質 香豆酸主要以沒有香氣的糖苷形式存在于液泡中,由于外界條件的改變可與葡萄糖苷酶接觸,在水解的同時閉環生成香豆素,可放出芳香氣味。豆科草木樨植株高大,產量高,是優質的蛋白牧草,但其香豆素含量較高,且具有特殊的香氣嚴重影響其適口性。有一種香豆素衍生物4-羥基香豆素,其通常形成于微生物的代謝過程,在發霉的含有香豆素的植物材料中,霉菌將反式鄰香豆酸轉化為4-羥基香豆素,兩分子4-羥基香豆素與甲醛自發結合生成雙香豆素[55],雙香豆素是眾所周知的抗凝血劑,會引起牲畜內出血導致死亡。因此含有香豆素的植物在貯藏或調制時如遇霉爛,體內的香豆素就轉變為雙香豆素或出血素,從而對牲畜產生危害。Luo等[56]對飼料作物白花草木樨半同胞家系進行多年多地點試驗評價,選育出了低香豆素草木樨育種材料,從而提高牧草品質。
2 香豆素的生物合成途徑
在生物合成中,簡單香豆素、呋喃香豆素、吡喃香豆素結構的轉化過程是簡單香豆素在6位或8位烷基化,取代異戊烯基與7位羥基形成呋喃環或者吡喃環的香豆素,進一步形成呋喃香豆素類和吡喃香豆素類。因此本研究主要以簡單香豆素生物合成途徑為重點進行綜述。
植物中的L-苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)能夠將L-苯丙氨酸轉化為反式肉桂酸,反式肉桂酸在肉桂酸2-羥化酶(cinnamic acid 2-hydroxylase,C2H)和肉桂酸4-羥化酶(cinnamic acid 4-hydroxylase,C4H)的作用下發生鄰羥基化和對羥基化反應,分別轉化為鄰香豆酸和對香豆酸,再經過進一步的反應最終形成香豆素類化合物等次生代謝產物。反式肉桂酸的對羥基化反應先于鄰羥基化反應[20],如果葉綠體中缺乏C4H時,C2H可以將肉桂酸轉化為鄰香豆酸[57]。香豆素生物合成途徑簡單框架如圖1[13,58—62]。

圖1 簡單香豆素生物合成途徑Fig.1 Pathway of simple coumarin biosynthesis
2.1 反式肉桂酸形成香豆素
香豆素的合成源于反式肉桂酸[63],反式肉桂酸首先在C2H作用下發生鄰羥基化生成鄰香豆酸[64],再與UDP-葡萄糖在2-香豆酸O-β-葡萄糖基轉移酶(UDP-glucosyltransferase,UGT)的作用下生成反式鄰香豆酸β-葡萄糖苷[58],在液泡中積累并發生反/順式異構化,在紫外線輻射下引起非酶光化學反應,轉換為順式香豆酸葡萄糖苷[65]。香豆素的非游離狀態就是以香豆酸葡萄糖苷的形式存在,它在葡萄糖苷酶(β-glucosidases,BGLU)的作用下釋放糖苷水解為香豆酸,最后自發內酯化形成游離香豆素[66]。
2.2 反式肉桂酸形成傘形花內酯等其他羥基化香豆素
反式肉桂酸在C4H的作用下進行對羥基化形成對香豆酸,首先與4-香豆酸CoA連接酶(4-coumarate CoA ligase,4CL)作用生成對香豆酰CoA,再與對香豆酰CoA 2’-羥化酶(p-coumaroyl CoA 2’-hydroxylase,C2’H)發生鄰羥基化作用生成2’,4’-二羥基香豆酰CoA[60],進而內酯化形成傘形花內酯(7-羥基香豆素)。
對香豆酸也可以先后與香豆酸-3-羥基化酶(4-coumarate-3-hydroxylase,C3H)、咖啡酸O-甲基轉移酶(caffeic acid O-methyltransferase,COMT)作用生成咖啡酸、阿魏酸,咖啡酸和阿魏酸分別與4CL作用生成咖啡酰CoA和阿魏酰CoA。對香豆酰CoA亦可先后在羥基肉桂酰基轉移酶(hydroxycinnamoyl transferase,HCT)、對香豆酰酯3’-羥化酶(p-coumaroylshikimate/quinate 3-hydroxylase,C3’H)和羥基肉桂基轉移酶作用下生成對香豆酰莽草酸/奎寧酸、咖啡酰莽草酸/奎寧酸、咖啡酰CoA[67],咖啡酰CoA再與咖啡酰輔酶A O-甲基轉移酶(caffeoyl CoA O-methyltransferase,CCoAOMT)作用生成阿魏酰CoA[68],接著在阿魏酰-CoA 6’-羥化酶1(feruloyl-CoA 6’-hydroxylase1,F6’H1)的作用下發生鄰羥基化生成6-羥基阿魏酰CoA[69],并在香豆素合酶(coumarin synthase,COSY)的作用下發生內酯化形成東莨菪內酯[61]。F6’H 1是東莨菪內酯生物合成的控制酶。東莨菪內酯經東莨菪內酯8-羥化酶(scopoletin 8-hydroxylase,S8H)生成秦皮素[70],植物細胞色素P450酶(cytochrome P450 82C4,CYP82C4)進一步把秦皮素氧化為兒茶酚香豆素[51]。
咖啡酰CoA可在體外與各種含有酚酶活性的植物提取物作用轉化成七葉亭(6,7-二羥基香豆素)[63],而七葉亭的有效前體是傘形花內酯而不是咖啡酸,且傘形花內酯的6-羥基化可能是在P450單加氧酶的作用下完成的[71]。Bayoumi等[72]的研究也提出了東莨菪內酯及其糖苷的生成是通過對香豆酸到咖啡酸再到阿魏酸的步驟實現的,而七葉亭及其糖苷的合成是通過對香豆酸到2’,4’-二羥基肉桂酸再到傘形花內酯的步驟實現的。
此外,Liu等[55]發現了4-羥基香豆素新的生物合成途徑,它是由聯苯合酶(biphenyl synthase,BIS)催化底物水楊酸酰基CoA而合成,是一種不需要鄰羥基化的生物合成途徑。
3 香豆素生物合成關鍵酶基因
Luo等[73]對香豆素含量存在顯著差異的5份白花草木樨進行轉錄組測序分析,通過京都基因與基因組百科全書(Kyoto Encyclopedia of Genes and Genomes,KEGG)分析鑒定出213個unigenes屬于苯丙氨酸合成途徑,其中在種質間差異表達的unigenes有17個,初步歸納了白花草木樨香豆素合成通路,總結出已知香豆素合成途徑相關酶,包括PAL、C4H、4CL、C3H、HCT、CCoAOMT、COMT和BGLU;Wu等[74]對不同香豆素含量的5份白花草木樨可能調控香豆素合成基因的microRNA(miRNA)進行了分析,共鑒定出417個已知和76個未知的miRNAs,利用GO和KEGG對預測的4196個靶基因進行功能注釋,預測出2個HCT基因和3個miRNA參與香豆素生物合成途徑。研究者在香豆素的合成途徑中,挖掘到了PAL、C4H、HCT、C3H等多個參與香豆素生物合成的基因。本研究對香豆素生物合成通路涉及的關鍵酶進行總結,具體信息見表1。
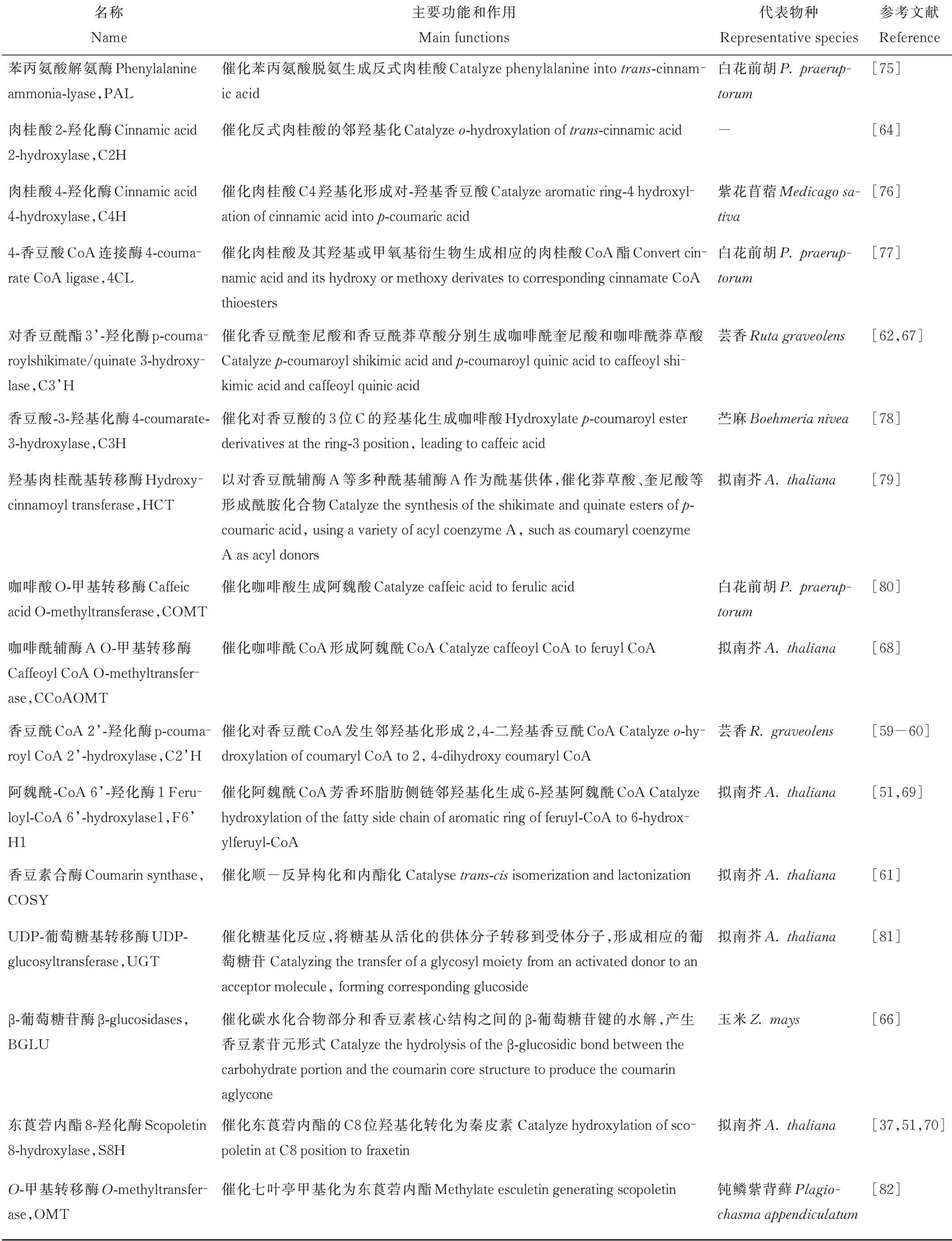
表1 香豆素生物合成途徑關鍵酶及其作用Table 1 Key enzymes and their functions in coumarin biosynthetic pathway
香豆素合成通路中的大多基因都是以基因家族的形式存在于植物中,不同亞族參與不同的代謝過程。PAL作為限速酶來調節進入苯丙烷代謝的總通量,擬南芥AtPAL 1、AtPAL 2、AtPAL 4與木質素的生物合成密切相關,同時發現AtPAL 1和PAL2控制類黃酮的合成[83]。4CL基因分為ClassⅠ和ClassⅡ兩組,其中ClassⅠ與木質素合成相關,ClassⅡ參與類黃酮生物合成[84]。Liu等[77]從白花前胡中分離出了4CL 1、4CL7和4CL10,明確了Pp4CL1是參與香豆素生物合成的酶。Vogt[68]認為,CCoAOMT不僅與木質素的合成相關,也可以催化阿魏酰CoA合成東莨菪內酯[69]。擬南芥根中東莨菪內酯和東莨菪苷的產生需要C3H的活性,在c3h突變體中,東莨菪內酯和東莨菪苷的含量下降3%[11]。另外參與香豆素合成的酶基因C2H、C2’H、F6’H和S8H均屬于2-氧戊二酸依賴雙加氧酶(2-oxoglutarate-dependent dioxygenases,2OGDs)超家族。Kawai等[85]將2OGDs分為DOXA、DOXB和DOXC三類,其中參與簡單香豆素生物合成的鄰羥基化酶屬于DOXC30,蕓香RgC2’H僅催化p-香豆酰CoA形成傘形花內酯[60],甘薯(Ipomoea batatas)2OGDs基因根據其底物特異性分為Ib1s和Ib2s兩組,Ib1s對阿魏酰CoA具有鄰羥基化活性,而Ib2s具有對p-香豆酰CoA和阿魏酰CoA的雙重活性,且主要利用p-香豆酰CoA合成傘形花內酯[86];擬南芥AtF6’H 1介導阿魏酸CoA的鄰羥基化生成的東莨菪內酯和東莨菪苷是擬南芥中含量最多的香豆素[50],而AtF6’H2只是具有對肉桂酸CoA硫代酯的活性,對東莨菪苷含量變化無影響[59]。S8H和其同源基因F6′H 2同時轉化本氏煙草葉片,可顯著增加秦皮素的積累,是由于煙草葉片中自然存在阿魏酸,F6’H2的過量表達導致阿魏酰CoA合成東莨菪內酯,為S8H催化反應提供了底物,導致秦皮素含量顯著升高[70]。
Vanholme等[61]首次在擬南芥中發現COSY,證明它是傘形花內酯和東莨菪內酯生物合成的關鍵酶,負責順反異構化和內酯化。COSY是BAHD-酰基轉移酶家族的成員之一,屬于V亞支,該亞支的蛋白作用于苯并酰CoA或羥基肉桂酰CoA[61]。一直以來認為在C2’H、F6’H 1作用下隨后的順反異構化和內酯化過程是由光催化且自發發生的,因此對這一步酶的要求一直被忽視,該項研究表明COSY在避光器官如根中的香豆素合成中具有重要功能。
BGLUs屬于糖苷水解酶I家族,催化碳水化合物部分和香豆素基本核心結構之間的β-葡萄糖苷鍵的水解,從而產生具有生物活性的香豆素苷元形式[87]。BGLU廣泛參與防御、細胞壁木質化、信號傳導、激素的活化和類黃酮代謝等植物次生代謝重要的生理過程[88],擬南芥中AtBGLU 21-23控制在根中特異產生的東莨菪苷的水解,在抵抗病原體攻擊和非生物脅迫中發揮作用,AtBGLU 42參與誘導機體對細菌疾病的抵抗力,并在缺鐵時對熒光香豆素在根際的分泌有重要作用[37]。
UGT屬于糖基轉移酶I家族,催化糖基活化轉移。根據環境條件,植物可以通過UGT將東莨菪內酯轉化為東莨菪苷,糖基化香豆素儲存在液泡中,在脅迫條件或植物受到病原菌侵染時,糖基化的香豆素與BGLU在細胞質中接觸,將東莨菪苷轉化為東莨菪內酯,使其發揮抗菌活性,清除H2O2對植物的傷害[13]。參與植物次生代謝的UGT具有廣泛的底物特異性,擬南芥UGT 84A 1和UGT 84A 3具有羥基肉桂酸葡萄糖基轉移酶活性[89];AtUGT 84A 1對咖啡酸、p-香豆酸有較高的親和力,AtUGT 84A 3對阿魏酸的活性最顯著[90];煙草UGT 73A 1和UGT73A 2介導香豆素東莨菪苷和七葉苷的合成[81]。
本研究根據已有文獻對香豆素合成關鍵基因在擬南芥、水稻和蒺藜苜蓿(Medicago truncatula)3種模式植物中的基因家族進行統計,結果見表2,以期為非模式植物香豆素合成基因的研究提供參考。

表2 3種模式植物的香豆素合成基因家族統計Table 2 Member statistics of coumar in synthesis gene families in three model plants
4 模式植物中UGT基因家族系統進化分析
在香豆素合成途徑中,UDP-葡萄糖基轉移酶(UGT)基因家族在香豆素代謝過程中起著重要作用,它的功能是參與植物代謝物的糖基化。糖基化過程在調節次生代謝產物的有效性方面起著重要的作用,可以增強分子的穩定性和水溶性,從而改變其在細胞內的運動,進而改變生物活性[94]。糖基化是由糖基轉移酶催化的一種常見的后修飾過程[95],能增加代謝物在植物體內的穩定性[96],有利于代謝物在體內積累和貯存[97]。
UGT基因家族的C末端含有1個由44個氨基酸組成的保守序列PSPG(plant secondary product glycosyltransferase)基序,該保守序列被認為是與UDP-糖供體結合的區域[98],但是僅憑序列信息通常無法可靠地預測物種間糖基化次生代謝產物的結構變化和功能[98],相反,高同源性的UGT可以作用于不同的底物[99—100]。
UGT分布廣泛,雙子葉植物與單子葉植物的基因外觀模式存在差異。然而,雙子葉植物和單子葉植物在各物種間所占基因數量上差異不大。本研究使用ClustalX v2.0對擬南芥121個[94]、水稻178個[94]以及蒺藜苜蓿243個[101]UGT氨基酸序列進行多序列比對,使用MEGA 7軟件利用鄰接法(neighbor-joining method,NJ)構建系統進化樹(圖2),bootstrap為1000。來自3種模式植物的UGT至少可以形成17個不同的組,其中擬南芥14個組(A~N),水稻15個組,蒺藜苜蓿15個組。水稻中缺失F組,說明F組是雙子葉植物特有的組,擬南芥中缺少O和P組,蒺藜苜蓿中缺少C和K組,Q組在3種模式植物中均缺失,R組只有蒺藜苜蓿存在。各組的個體數量各不相同,在D和E組中的UGT數量最多,其次是A、L和G組,這與Caputi等[94]的研究結果一致,在高等植物的進化過程中,每個物種的這5個類群比任何其他類群的擴張都要大。D組中蒺藜苜蓿比其他兩種植物擴張明顯。在I和G組中,擬南芥的UGT數量為1和7,水稻和蒺藜苜蓿分別為9、20和5、39,發現I和G組在水稻和蒺藜苜蓿中有明顯的擴張。根據文獻報道,大豆A組中GmSGT 2和GmSGT 3參與大豆皂苷I的生物合成;擬南芥D和E組功能相似,參與萜類化合物、類黃酮、苯甲酸鹽和油菜素類固醇生物合成;L組可以催化苯丙烷類代謝產物羧基的葡萄糖酯的形成;G組參與萜類化合物合成;C、I、J、K、M、N組的功能尚未明確[94],因此預測D、E和L組最有可能與香豆素合成相關。

圖2 UG T基因家族在模式植物中的進化分析Fig.2 Phylogenetic analysis of the UGT gene family in three model plants
5 結語與展望
本研究以香豆素作為切入點,對香豆素生物合成途徑中的關鍵酶基因研究進展進行綜述。雖然目前大量研究已對香豆素生物合成相關基因進行了定位,并對生物合成途徑中諸多基因進行克隆、鑒定及部分功能經過驗證,然而更多的研究是針對香豆素合成途徑中單個基因的功能研究,基因表達量的上調和下調與香豆素含量之間的關系仍有待進一步探討;且由于該代謝網絡復雜,關鍵基因是單一地參與香豆素的生物合成,還是與上下游的關鍵酶基因形成調控網絡參與到香豆素的生物合成,目前暫無明確定論;且結構基因和轉錄因子在香豆素生物合成途徑的調控網絡尚較模糊,多基因以及多轉錄因子間蛋白互作及其對香豆素合成相關結構基因的調控機制尚未挖掘清楚,具體調控的靶基因及有關轉錄因子的作用機制還不透徹。另外,香豆素含量高的草木樨等植物由于遺傳轉化體系不成熟等因素,低香豆素含量品系的選育一直沒有取得明顯突破。
結合當前研究存在的問題,今后對香豆素生物合成的研究應集中在以下幾點:1)進一步在香豆素生物合成的下游途徑進行探索;2)利用轉錄組學、蛋白組學、代謝組學等現代生物技術發掘更多與香豆素生物合成的關鍵基因,確定關鍵基因之間的關系,基于多組學的網絡構建和完善香豆素生物合成調控網絡;3)克隆出相關調控香豆素合成基因后,通過轉基因技術轉入模式植物中進行功能驗證,并進一步構建非模式植物自身轉基因體系進行功能驗證;4)基因過表達或敲除以及基因編輯技術已逐步取得突破,從遺傳學角度高效準確地深入研究并驗證香豆素生物合成過程中不同基因的生物學功能,這對將來通過基因工程進行作物的改良育種具有重要意義。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
