熱應激對豬顆粒細胞蛋白質表達譜的影響
2022-02-05 11:12:16王澤平趙為民付言峰李碧俠任守文程金花
江蘇農業學報 2022年6期
王澤平, 沈 婕, 趙為民, 付言峰, 李碧俠, 任守文, 程金花, 李 輝
(1.江蘇省農業科學院宿遷市農業科學研究所,江蘇宿遷223800;2.江蘇省農業科學院畜牧研究所,江蘇南京210014;3.廣西大學動物科學技術學院,廣西南寧530005)
豬的皮膚較厚,皮下脂肪含量高且汗腺不發達,故調節體溫能力相對較差。研究結果表明,豬最適溫度為12~21 ℃[1],當環境溫度由18 ℃升至29 ℃時,豬體溫將從34.6 ℃升至37.4 ℃[2],而直腸溫度將從39.2 ℃升高至40.0 ℃[3]。因此當外界氣溫超過27 ℃時,母豬體溫也將被動升高而產生熱應激[4],并對豬機體造成不良影響。以母豬為例,當受到熱應激時可表現出發情不明顯、乏情或短促發情等癥狀,經產母豬斷奶7 d內發情率顯著降低,而后備母豬則表現出初情期推遲等現象[5-6]。夏秋時節,中國大部分地區的氣溫超過30 ℃,因此母豬受到熱應激是夏秋季節造成中國養豬業受損的重要因素,隨著全球變暖,熱應激的影響將更加嚴重。然而,目前人們對熱應激影響母豬繁殖性能的機制并不清楚。
顆粒細胞介于膜細胞和卵母細胞之間,是體內唯一與卵母細胞直接接觸的體細胞。它們可以通過分泌類固醇類激素、供給卵母細胞營養物質、接收并傳遞信號等方式參與維持卵泡發育、促進卵母細胞成熟和誘導排卵等過程。顆粒細胞功能受損將影響動物卵泡發育和繁殖活動[7]。養殖過程中,動物常常受到一些因素的影響,導致顆粒細胞功能紊亂,主要表現在類固醇激素分泌能力下降、細胞增殖抑制/凋亡、黃體化受阻等[8-10],其直接后果是抑制動物的繁殖性能,最終影響畜牧業的健康發展。遺憾的是,目前關于熱應激直接影響顆粒細胞的研究相對較少且大多從單一角度或者在某一點上展開研究,例如,前人研究發現熱應激可抑制顆粒細胞中促卵泡素受體(FSHR)表達,并促進細胞凋亡[11]。我們前期研究也發現熱應激處理豬顆粒細胞可顯著抑制雌激素的分泌,造成熱休克蛋白HSP70表達量上調[9, 12]。總體而言,目前在熱應激影響卵泡顆粒細胞功能方面缺乏系統性、整體性的認識。為解決該問題,本研究擬先解析熱應激對豬顆粒細胞蛋白質表達譜的影響。
1 材料與方法
1.1 試驗材料
將在屠宰場取的豬卵巢,放置于裝有37 ℃生理鹽水(預先加入1%青霉素/鏈霉素)的保溫瓶中盡快拿回實驗室處理。在實驗室超凈臺中清洗消毒卵巢,擦凈殘留液體后,選取直徑大于5 mm卵泡,用一次性注射器抽取其卵泡液,并利用Ficoll-Paque梯度離心法分離出顆粒細胞[13]。分離出的細胞經多次清洗后用DMEM/F12培養液(含10%胎牛血清、1%青霉素/鏈霉素)稀釋后接種于6孔板中,并置于37 ℃,含有5% CO2、95%空氣的培養箱中持續靜止培養24 h后,用溫熱的無Ca2+、Mg2+的磷酸鹽緩沖溶液(PBS)清洗2~3次,洗掉未貼壁細胞,更換新的DMEM/F12培養液(含10%胎牛血清、1%青霉素/鏈霉素)后進行后續處理。
1.2 熱應激處理
將獲得的細胞隨機分為2組。一組為對照組,將細胞放入37 ℃的細胞培養箱中培養3 h;另一組作為熱應激組,將細胞放入41 ℃的細胞培養箱中培養3 h。處理結束后,用溫熱的無Ca2+、Mg2+的PBS徹底清洗細胞,確保無培養基殘留,然后將細胞刮下備用。所有試驗重復3次。
1.3 蛋白質的提取及定量
收集的細胞按照蛋白質組學的標準流程進行處理。處理過程簡述如下:細胞沉淀先用四乙基溴化銨緩沖溶液(TEAB)溶解;裂解后的初始蛋白質溶液經超聲破碎后離心并收集上清液,棄掉細胞碎片。在收集的上清液中加入4倍體積的預冷丙酮[含有10 mmol/L的二硫蘇糖醇(DTT)],充分混勻后放置在4 ℃冰箱中靜置2 h,蛋白質沉淀逐漸析出后通過離心將蛋白質沉淀收集至管底,再用預冷丙酮(含10 mmol/L的DTT)清洗蛋白質沉淀后進行干燥。最終用四乙基溴化銨(TEAB)溶解蛋白質沉淀,并對蛋白質進行定量。
1.4 蛋白質樣品酶解、脫鹽及標記
選取一定質量的蛋白質溶液,調整體積后用NH4HCO3(50 mmol/L)進行稀釋,加入胰蛋白酶溶液,置于37 ℃培養箱中酶解過夜。酶解結束后將酶解液進行脫鹽處理;脫鹽后的蛋白質按照試劑盒說明書進行標記。標記結束后在所有標記樣本中取相同體積進行混合,準備上機分離。
1.5 蛋白質樣品組分分離及液相色譜-質譜/質譜(LC-MS/MS)檢測
將上述混合后的樣本用Thermo DINOEX Ultimate 3000 BioRS分成12個組分,并利用AB SCIEX nano LC-MS/MS(Triple TOF 5600 plus)對每個組分依次進行質譜檢測。
1.6 蛋白質的鑒定、差異表達蛋白質的功能注釋和通路分析
以可信度95%為標準對所得的肽段進行篩選,可信度大于95%的肽段用搜索引擎ProteinpilotTMV4.5進行搜索。當差異表達倍數≥1.5倍(即上調≥1.5倍或下調≤0.67倍)且P值≤0.05時,視為顯著差異表達蛋白質,并利用在線分析系統BLAST2GO (http://www.Blast2GO.com/)對所有差異表達蛋白質進行GO功能注釋,隨后利用通路數據庫KEGG進行通路分析。
2 結果與分析
2.1 熱應激后豬顆粒細胞的蛋白質總體鑒定情況
利用同位素標記相對和絕對定量(iTRAQ)技術對受熱應激處理的豬顆粒細胞的蛋白質進行定量分析,共鑒定到5 451個蛋白質。鑒定肽段數為35 936個,其中至少包含2個Unique肽段的蛋白質數為4 448個。本研究鑒定到的肽段的長度主要分布在7.00~20.00 aa,其中大多數肽段的長度為11.00 aa,平均長度為14.57 aa,處于肽段長度合理范圍(圖1A)。對可信度≥95%的肽段覆蓋度分布的分析結果顯示,覆蓋度介于0~10.0%的蛋白質數量占總鑒定蛋白質數量的38.49%(圖1B)。
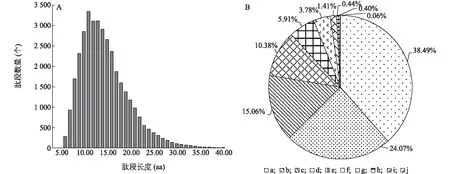
A:肽段長度分布;B:蛋白質鑒定覆蓋度分布。a:0~10.0%; b:10.1%~20.0%; c:20.1%~30.0%; d:30.1%~40.0%; e:40.1%~50.0%; f:50.1%~60.0%; g:60.1%~70.0%; h:70.1%~80.0%; i:80.1%~90.0%; j:90.0%~100.0%。圖1 肽段長度分布和蛋白質覆蓋度Fig.1 Distribution of peptides length and protein sequence coverage
2.2 差異表達蛋白質分析
通過對熱應激處理的豬顆粒細胞中蛋白質表達的分析,共鑒定到289個差異表達蛋白質,其中上調表達蛋白質162個,下調表達蛋白質127個(部分差異表達蛋白質見表1和表2)。從表1中可以看出,熱休克蛋白HSP70、HSP90等與熱應激相關的標志性蛋白質上調表達。這些蛋白質的上調表達也可被視為熱應激處理成功的標志。同時,肽基脯氨酸順反異構酶、鈣網蛋白、鈣調理蛋白等蛋白質上調表達(表1)。而組蛋白H4、載脂蛋白E、芳香化酶3、雌二醇17β-脫氫酶、三磷酸腺苷合成酶α亞基等蛋白質表達顯著下調(表2)。
我們進一步對對照組和熱應激組樣品的分析結果進行聚類分析,從圖2可以看出,3個生物學重復之間重復性良好,且差異表達蛋白質的聚類清晰。
2.3 差異表達蛋白質的直系同源簇(COG)注釋
為預測和系統了解所鑒定到的蛋白質的潛在生物學功能,我們對所有鑒定到的蛋白質與COG數據庫進行了比對分析和分類統計。圖3是本次鑒定結果比對到COG數據庫的統計結果。
2.4 差異表達蛋白質的GO分類和KEGG分析
為解析差異表達蛋白質的生物學功能,本研究對所有的差異表達蛋白質進行了注釋。如圖4所示,本研究所鑒定到的差異表達蛋白質參與了細胞過程、代謝過程、生物調控、應激反應等相關功能的調控。

表1 部分上調表達的差異蛋白質
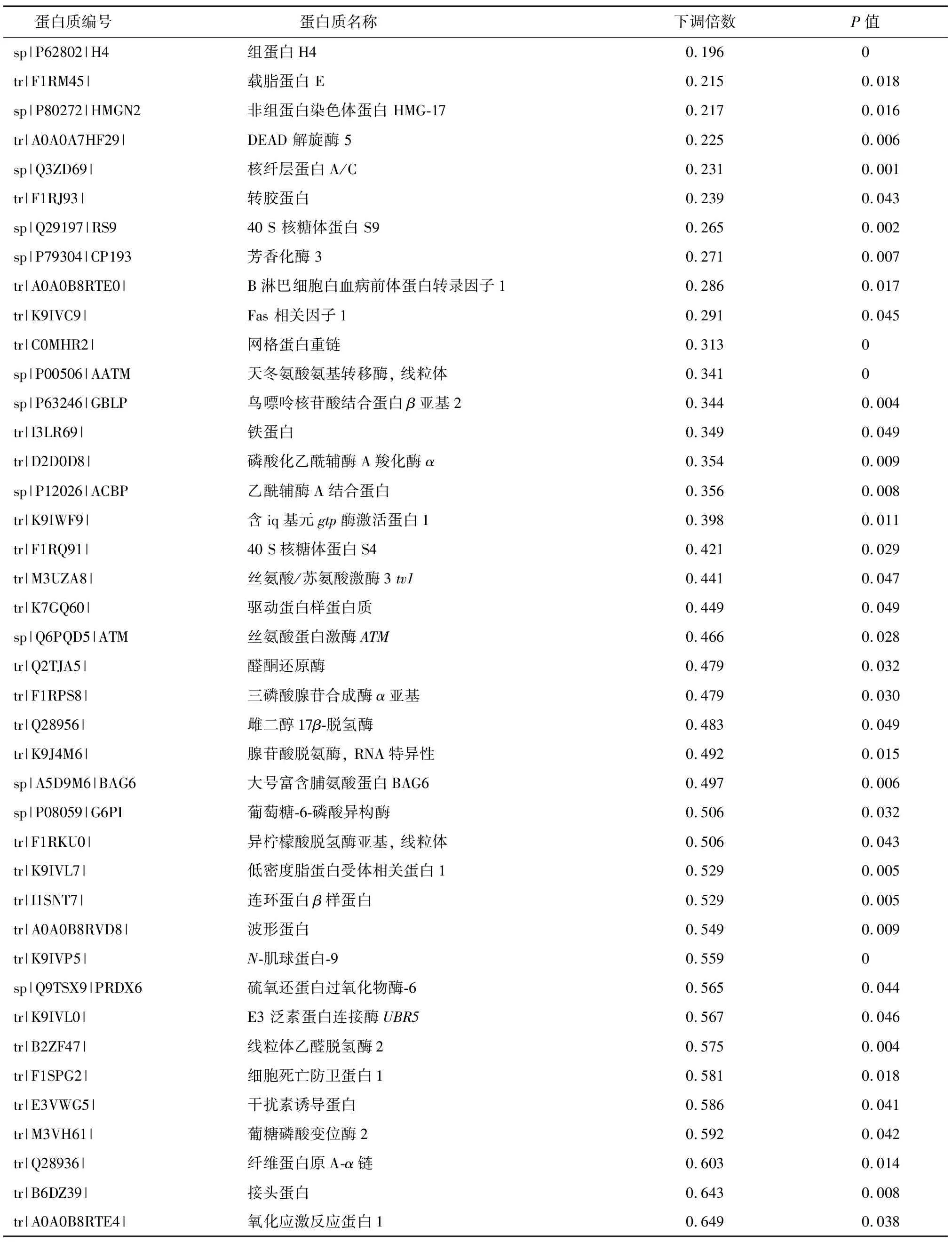
表2 部分下調表達的差異蛋白質
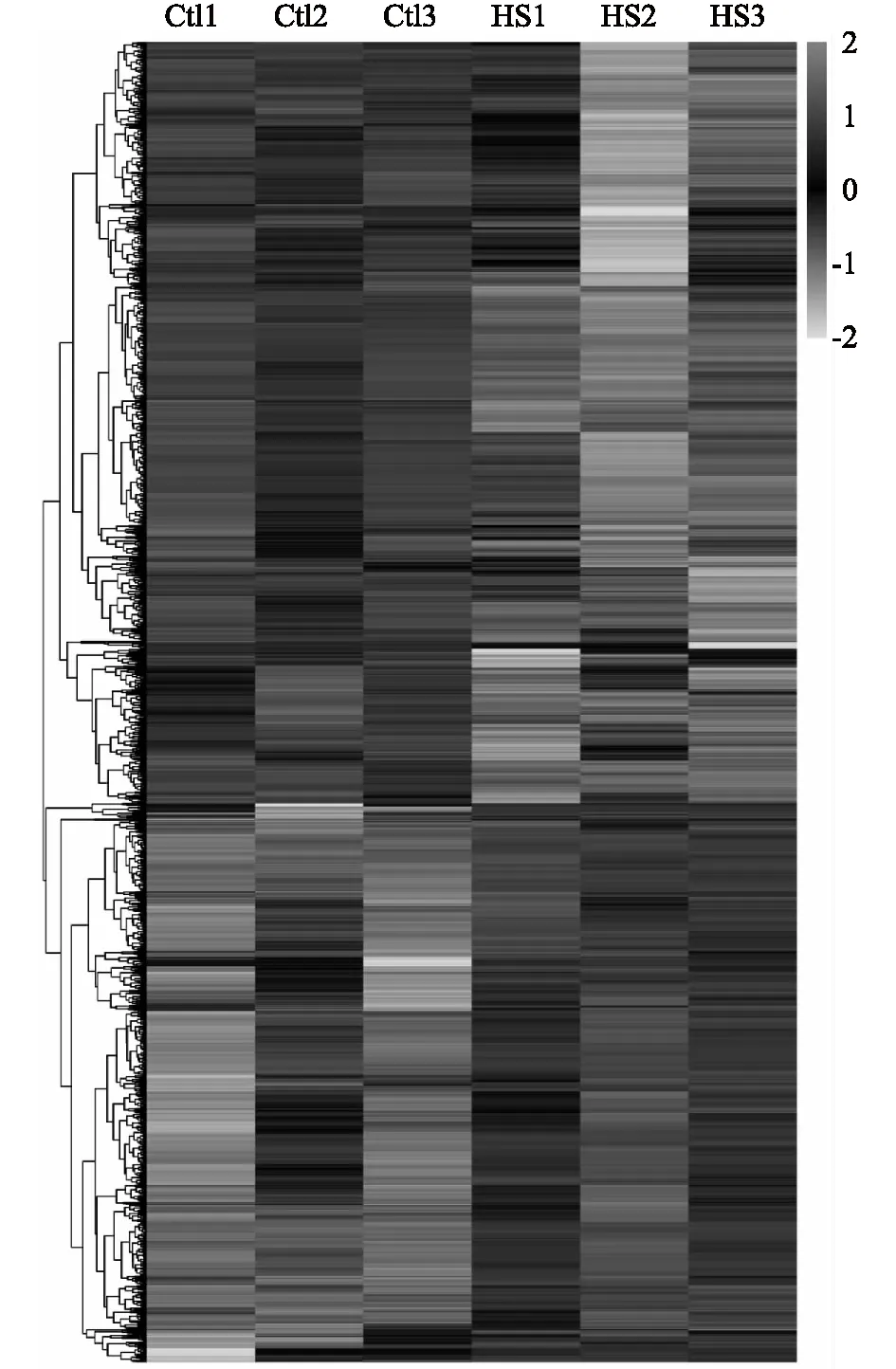
Ctl:對照;HS: 熱應激。圖2 豬顆粒細胞受熱應激過程中差異表達蛋白質的聚類分析Fig.2 Clustering results of differentially expressed proteins in porcine granulosa cells during heat stress
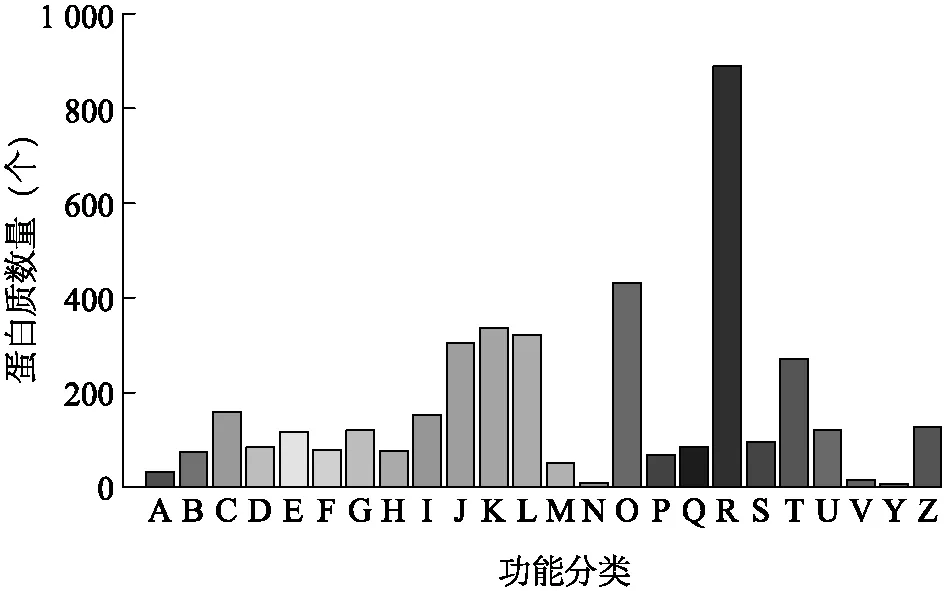
A:RNA加工和修飾; B:染色質結構和動態變化; C:能量產生和變化; D:細胞周期控制,細胞分裂,染色體分割; E:氨基酸運輸和代謝; F:核苷酸轉移和代謝; G:碳水化合物轉移和代謝; H:輔酶轉移和代謝; I:脂類轉移和代謝; J:翻譯,核糖體結構和生物發生; K:轉錄; L:復制,重組和修復; M:細胞壁/膜/殼生成; N:細胞運動; O:轉錄后修飾,蛋白質折疊,分子伴侶; P:無機鹽離子轉運和代謝; Q:次級代謝產物生成,轉運和代謝; R:功能預測; S:未知功能; T:信號轉導機制; U:胞內交通,分泌和微囊轉運; V:防御機制; Y:核結構; Z:細胞骨架。圖3 蛋白質的直系同源簇注釋分析Fig.3 COG annotation analysis of proteins

1:細胞過程; 2:代謝過程; 3:生物調控; 4:應激反應; 5:生物過程調控; 6:細胞組分配置或生物發生; 7:發育過程; 8:多細胞生物過程; 9:定位; 10:定位建立; 11:信號; 12:死亡; 13:免疫系統過程; 14:正調控生物過程; 15:負調控生物過程; 16:細胞增殖; 17:運動; 18:繁殖; 19:繁殖過程; 20:多物種過程; 21:生長; 22:病毒繁殖; 23:生物黏附; 24:細胞; 25:細胞組分; 26:細胞器; 27:細胞器組分; 28:大分子復合物; 29:膜封閉腔; 30:胞外域; 31:部分胞外域; 32:結合; 33:催化活性; 34:結構分子活動; 35:轉運活性; 36:能量調控活性; 37:蛋白質結合轉錄因子活性; 38:抗氧化活性; 39:受體調控活性。圖4 差異表達蛋白質的GO功能注釋結果Fig.4 GO function annotation results of differentially expressed proteins
為進一步了解差異表達蛋白質所富集的通路,進行了KEGG通路分析。如圖5所示,表達量上調的蛋白質主要富集在剪接體以及內質網的蛋白質加工、代謝通路、胞吞、RNA降解和氧化磷酸化等信號通路中。而下調表達蛋白質則顯著富集在代謝通路、次級代謝產物的生物合成、嘌呤代謝、泛素化蛋白質降解、糖酵解/糖異生以及胞間緊密連接等信號通路中。

A:上調表達蛋白質富集的通路及數量占比;B:下調表達蛋白質富集的通路及數量占比。a1:剪接體; b1:內質網蛋白質加工; c1:代謝通路; d1:胞吞; e1:帕金森病; f1:亨廷頓癥; g1:抗原加工和遞呈; h1:RNA降解; i1:氧化磷酸化; j1:阿爾茨海默病。a:代謝通路; b:次級代謝產物的生物合成; c:不同環境下微生物的代謝; d:嘌呤代謝; e:泛素化蛋白質降解; f:糖酵解/糖異生; g:胞間緊密連接; h:磷酸戊糖途徑; i:淀粉和糖代謝; j:氨糖和核糖代謝。圖5 差異表達蛋白質的KEGG通路分析結果Fig.5 KEGG pathway analysis results of differentially expressed proteins
3 討論
熱應激可導致雌性動物短促發情或乏情、降低受胎率、早期胚胎死亡和流產等問題[12]。前人研究發現,熱應激可經下丘腦-垂體-性腺軸對動物的內分泌產生不良影響進而造成上述問題[14]。熱應激導致動物內分泌紊亂主要表現在性激素[促性腺激素釋放激素(GnRH)、促卵泡激素(FSH)和促黃體激素(LH)等]分泌量下降、抑制促性腺激素受體基因的表達[15-17]等。此外,通過下丘腦-垂體-性腺軸,熱應激還可調控其他激素的分泌,例如抑制促甲狀腺激素[18]、甲狀腺激素[19]以及促腎上腺皮質激素釋放激素[20]等激素的合成和分泌,從而增加腎上腺皮質激素分泌量,造成黃體提早溶解,引起早期胚胎死亡或流產[21]。
熱應激通過下丘腦-垂體-性腺軸對顆粒細胞功能產生影響,是其重要的內分泌調控路徑。然而,就顆粒細胞所處的位置來講,其位于動物腹腔最深處,受熱應激時,豬機體深處的體溫,即核心體溫顯著升高[22],升高的核心溫度必然直接作用于顆粒細胞而影響其功能。遺憾的是,當前對于熱應激直接影響顆粒細胞的研究較少。因此本研究通過體外培養的方式,將顆粒細胞所處的卵泡微環境、下丘腦-垂體-性腺軸以及內分泌環境的影響剝離,純粹在細胞水平上對熱應激調控顆粒細胞功能的分子機理開展研究。我們前期在體外模擬了顆粒細胞受到熱應激的狀態:將顆粒細胞分別在37 ℃(對照組)和41 ℃(熱應激組)培養箱中培養,結果發現,顆粒細胞合成雌激素和孕酮的能力隨著溫度的升高顯著下降,且與之相關的CYP11A1、CYP19A1和FSHR基因表達量顯著下調,但HSP70的表達量顯著上調[9]。上述結果說明熱應激可直接對顆粒細胞的功能產生影響。
為深入了解熱應激對豬卵泡顆粒細胞功能的影響,并最終為解決受熱應激的母豬繁殖性能降低的問題提供理論依據,本研究利用iTRAQ技術對受熱應激處理的豬顆粒細胞的蛋白質表達差異進行了系統研究,共鑒定到5 451個蛋白質。這是目前已知鑒定到的相對較多的豬卵泡顆粒細胞蛋白質組表達譜,并篩選出了289個差異表達蛋白質,其中上調表達蛋白質162個,下調表達蛋白質127個。通過對這些差異表達蛋白質的GO分類和KEGG通路分析,初步闡明其分類屬性和富集的功能通路。我們推測這些差異表達蛋白質在細胞抵抗熱應激和減輕熱應激對細胞造成損傷方面起到重要作用。例如,熱休克蛋白HSP70和HSP90是細胞受熱應激的標志性蛋白質,作為分子伴侶,它們參與蛋白質折疊,在抵御不利因素造成的應激或凋亡過程中發揮了重要作用[23],其表達量的升高也標志著細胞正在抵御熱應激帶來的損害[24]。
在對差異表達蛋白質富集的KEGG通路分析中我們發現了參與內質網蛋白質加工、RNA降解以及氧化磷酸化等過程的重要通路,說明這些通路的激活參與顆粒細胞抵抗熱應激的過程。以氧化磷酸化為例,該過程是還原型煙酰胺腺嘌呤二核苷酸(NADH)通過氧化制造細胞代謝的重要能量載體——三磷酸腺苷(ATP)的過程[25],其詳細過程為:電子經電子傳遞鏈從供體轉移至載體上,最后傳遞到受體,同時釋放能量以便ATP的合成[26]。Yin等[27]也發現熱應激處理可促進卵丘顆粒細胞中ATP的合成,在我們前期研究中發現脂多糖(LPS)處理亦可上調豬顆粒細胞中ATP的合成[28]。這些結果充分說明,細胞提高胞內ATP的合成可能是為了抵御受到的不良刺激。然而合成的ATP如何參與抵御不良刺激,目前尚不清晰,這需要我們通過進一步的研究加以闡明。
4 結論
本研究利用iTRAQ技術對受熱應激處理的豬顆粒細胞的蛋白質表達譜進行了研究,初步篩選出289個差異表達蛋白質,并分別富集到蛋白質的泛素化降解、蛋白質加工、胞吞、RNA降解、氧化磷酸化、嘌呤代謝、次生代謝產物的生物合成、糖酵解以及胞間緊密連接等信號通路中,也預示這些差異表達蛋白質和信號通路參與了熱應激對豬顆粒細胞功能影響的過程。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國科技論壇(2017年7期)2017-07-25 08:49:53
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
