西雙版納勐海縣不同生境蛛蜂群落結構及優勢類群動態研究
2022-02-03 07:58:16濮永勝陶玉王金秀趙苓李強馬麗
南方農業學報 2022年10期
關鍵詞:物種
濮永勝,陶玉,王金秀,趙苓,李強,馬麗
(云南農業大學植物保護學院,云南昆明 650201)
0 引言
【研究意義】蛛蜂隸屬于膜翅目(Hymenoptera)細腰亞目(Apocrita)針尾部(Aculeata)胡蜂總科(Vespoidea)蛛蜂科(Pompilidae)。蛛蜂成蟲采食花蜜,幼蟲靠成蟲捕獵蜘蛛喂食(Kurczewski et al.,2020)。蛛蜂在生態環境中擔任著傳粉昆蟲與天敵昆蟲的角色,是將動物與植物、動物與動物聯系起來的一個重要類群(Evans,1997)。蛛蜂種類繁多,種群數量龐大,廣泛分布于多種生態環境中,其種類和種群數量隨生態環境的變化而變化。西雙版納勐海縣屬熱帶、亞熱帶季風氣候,終年暖熱,冬無嚴寒、夏無酷暑,是我國生物多樣性最豐富的地區之一。選取西雙版納勐海縣賀開村4種不同生境開展不同生境蛛蜂群落研究,可為探討蛛蜂的生態功能及其保護與利用提供科學依據。【前人研究進展】目前世界已記錄蛛蜂5個亞科5000余種,我國已知3亞科240余種。蛛蜂種類多,其屬間的生物學習性差異較大,性喜陽光,廣泛分布于沙地、果園、耕地、林木和草地等多種生態環境中。蛛蜂社會行為不發達,多為獨居蜂,筑巢、捕獵、飼育幼蟲等活動多數情況下由雌蜂單獨進行,尚未有蛛蜂具社會性的報道。筑巢方式多種多樣,巢穴筑在土中、腐木上,或石塊縫隙間,部分利用其他動物廢棄的巢穴,或昆蟲的蛀道和有隧道植物的莖干等,很少在遠離沙土的地方筑巢(Howard and Robert,1973;Christophe and James,2012)。就目前所知,大部分蛛蜂的巢室為單個,部分為多巢室,每室1頭蛛蜂。在長期的進化過程中,蛛蜂形成了自己獨特的捕獵行為和生態環境適應性,生物學習性復雜,在膜翅目的系統發育中代表了一個特殊分支。蛛蜂以寄生、捕食蜘蛛或盜寄生其他蛛蜂的巢穴為生,體型小到大型,性喜陽光,行動敏捷。常在沙丘、干濕地、溪流和池塘附近、林中的空地和小徑上活動,利用細長的腿沿著低的植被,在地上快速移動,不易被發現和捕捉。該科成蟲主要以花蜜為生,壽命不足1年。雌蜂產卵很少,每隔2~3周產卵1次,1次僅產1枚;當雌性蛛蜂“卵巢”中的卵孕育到一定程度時,便外出搜尋獵殺蜘蛛,利用蟄針刺蟄并麻痹獵物,搬回巢穴產卵,幼蟲一般在2~3 d內孵出,孵出后即開始取食被麻痹的蜘蛛(Andrietti et al.,2008)。目前國內蛛蜂研究多為種類及區系研究(紀曉玲等,2015;蔣力等,2016;江鑫等,2016;朱健等,2016;李穎等,2019),蛛蜂與生態環境之間關系的研究較少,僅王金秀等(2021)對西雙版納地區不同生態環境演替階段的蛛蜂群落進行了研究,發現蛛蜂物種多樣性和群落特征與生態環境演替密切相關,人為因素是造成蛛蜂群落差異的重要原因。此外,國內許多學者對西雙版納其他昆蟲的多樣性進行了研究,發現不同生境不同地域分布不同的昆蟲種類,如不同類群的蝴蝶分布在不同的生態環境中(陳明勇,2001)、不同的薊馬物種分布在不同的樹冠層中(魯雯敏,2020)。西雙版納昆蟲物種豐富,擁有其他區域未分布的物種種類,如蝽類(章士美和林毓鑒,1990)及部分螞蟻(徐正會,1999),這些研究均充分表明昆蟲群落與生態環境之間有著密切的聯系,研究西雙版納昆蟲與環境之間的關系有助于當地生物多樣性的保護與利用。【本研究切入點】蛛蜂作為傳粉昆蟲與天敵昆蟲在生態系統中有著重要地位,與生態環境之間有著密切聯系,但前人對蛛蜂與生態環境之間關系的研究較少,且對其他昆蟲主要著眼于大范圍的調查研究,側重于小區域的調查相對較少。【擬解決的關鍵問題】通過馬氏網及黃盤誘集法對西雙版納幾種不同生境的蛛蜂群落進行調查,以摸清蛛蜂在不同生境下的群落結構及其差異、不同類型生境下蛛蜂優勢類群及其動態變化,為蛛蜂與生態環境協同演化及傳粉昆蟲和天敵昆蟲的保護利用研究提供參考。
1 材料與方法
1.1 調查區域與樣地設置
調查區域位于西雙版納勐海縣賀開村(21°49.692′N,100°28.848′E),是一個典型的高原山間盆地,即海拔1200 m處地形較平坦,主要種植水稻、甘蔗、花生和茶樹等作物,是當地居民主要聚居地;海拔1500 m處地形陡峭,不適宜人類居住,多為當地百年以上普洱茶產地,及分布一些原始的落葉闊葉林。2種地形為西雙版納當地具有代表性的地形,不同地形的生態環境、植被類型相差很大。
于2018年4月—2019年5月,在西雙版納勐海縣賀開村采用樣地調查法,根據賀開村區域的生境多樣性設置4塊不同生態環境的樣地,分別為農田區(樣地Ⅰ)、農田與茶園交界區(農田與茶園混合生境)(樣地Ⅱ)、茶園區(樣地Ⅲ)、次生落葉闊葉林區(樣地Ⅳ),樣地概況如表1所示。

表1 4種樣地生境特征及植被組成Table 1 Habitat characteristics and vegetation composition of four sample plots
1.2 試驗方法
1.2.1 蛛蜂群落調查在每個樣地安裝1個Townes型馬氏網用于蛛蜂誘集,收集瓶內裝有95%酒精約400 mL,每個月換1次收集瓶。每次更換收集瓶的同時進行田間觀察和調查,在每個馬氏網周圍3~10 m內分別放置10個黃盤,黃盤中放置2/3的洗潔劑液用于蛛蜂補充收集,每次黃盤放置時間為24 h。將采集的蛛蜂標本放入95%酒精瓶中帶回實驗室進行標本制作,采用形態分類的方法進行分類鑒定,并將蛛蜂鑒定到種級分類階元,如不能鑒定到種則采用sp.表示,并記錄數據。
1.2.2 蛛蜂群落結構分析.根據蛛蜂物種個體數占群落總個體數的百分比將物種劃分為5個類型:≥10.00%為優勢類群;5.00%~9.99%為常見類群;1.00%~4.99%為較常見類群;0.10%~0.99%為較稀有類群;<0.10%為稀有類群(熊忠平等,2021)。
1.2.3 蛛蜂群落多樣性指標
Shannon-Wiener多樣性指數:H′=-∑(Ni/N)ln(Ni/N)
Berger-Parker優勢度指數:D=Nmax/N
Margalef豐富度指數:R=(S-1)/lnN
Pielou均勻度指數:J=H'/lnS
式中,Ni為第i種的個數;N為總個體數量;Nmax為優勢種群數量;S為物種數。
1.2.4 蛛蜂群落相似性
Jaccard相似性系數:q=c(/a+b-c)
式中,a為A群落類群數,b為B群落類群數,c為兩群落共有類群數。0.75<q≤1.00為極相似;0.50<q≤0.75為中等相似;0.25<q≤0.50為中等不相似;0.00<q≤0.25為極不相似。
1.3 統計分析
采用Excel 2010對試驗數據進行統計;使用SPSS 20.0對數據進行單因素方差分析;采用Origin 2018繪圖。
2 結果與分析
2.1 不同生境蛛蜂群落結構
本次調查在西雙版納勐海縣賀開村共采集到蛛蜂標本801號,隸屬于2亞科、33屬112種,其中包括2種優勢類群、4種常見類群、15種較常見類群和12種較稀有類群,未發現稀有類群;奧溝蛛蜂屬(Auplopus)的物種數量最多,為20種,占比17.86%;扁腹溝蛛蜂屬(Minagenia)個體數最多,為190頭,占比23.72%;狹鱗溝蛛蜂屬(Leptodialepis)、刺溝蛛蜂屬(Phanagenia)和Euplaniceps屬均僅有1種1頭個體,物種數量和個體數量分別占比0.89%和0.12%;各樣地蛛蜂物種數量排序為樣地Ⅰ>樣地Ⅲ>樣地Ⅱ>樣地Ⅳ,個體數量排序為樣地Ⅰ>樣地Ⅳ>樣地Ⅲ>樣地Ⅱ;同一個屬在不同生境中的物種數和個體數不同,其中4種生境中共有的屬有9個;Poecilopompilus、副灣蛛蜂屬、鐵色蛛蜂屬、童蛛蜂屬、盜蛛蜂屬和隱唇溝蛛蜂屬僅分布于農田生境,刺溝蛛蜂屬僅分布于農田與茶園混合生境,狹鱗溝蛛蜂屬僅分布于落葉闊葉林生境(表2)。結果表明4種不同生境中蛛蜂群落組成存在明顯差異。
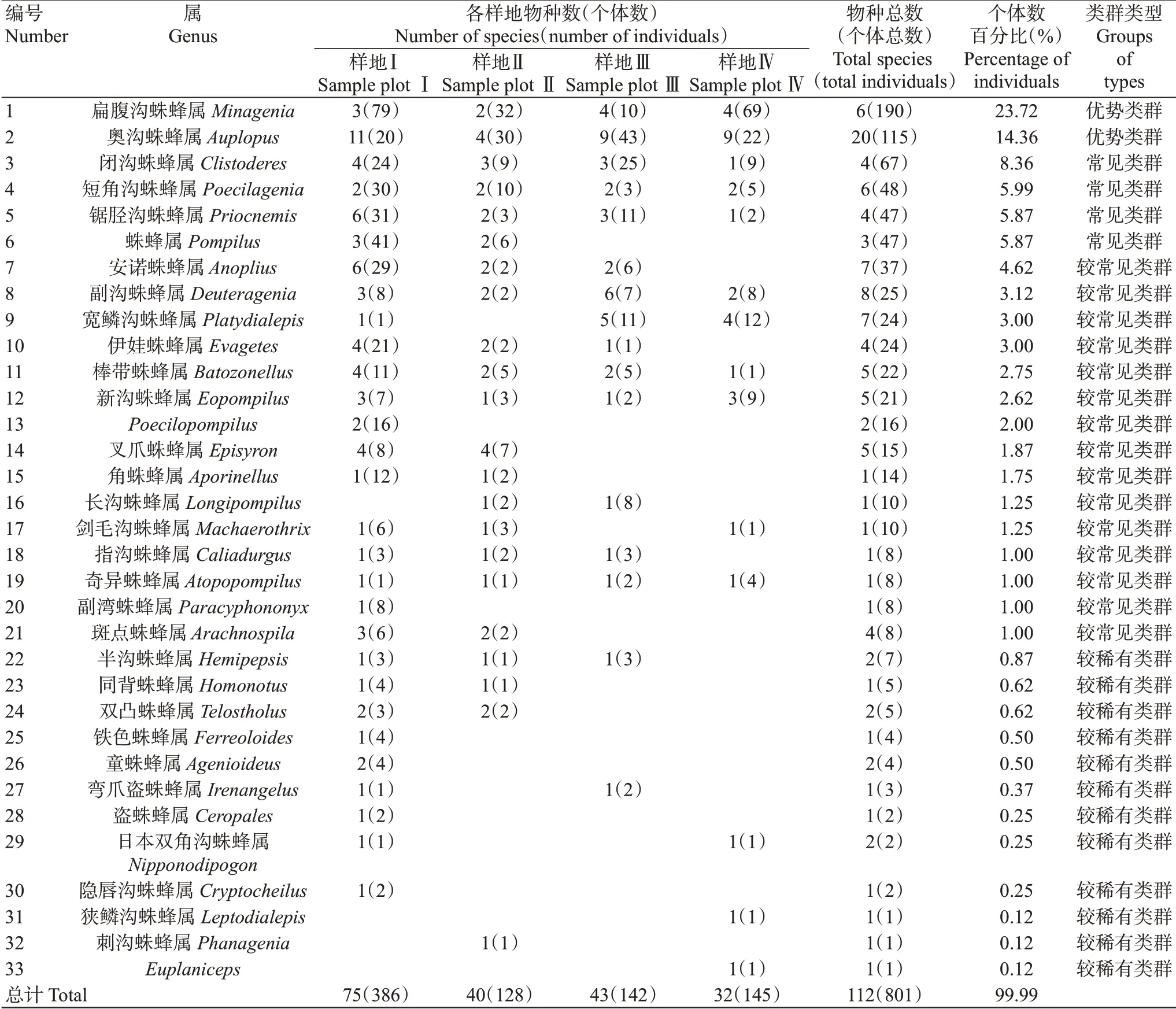
表2 不同生境蛛蜂群落結構Table 2 Spider wasp community structure in different habitats
2.2 不同生境蛛蜂群落多樣性特征
如表3所示,4個樣地蛛蜂群落的Margalef豐富度指數(R)排序為:樣地Ⅰ(12.42)>樣地Ⅲ(8.47)>樣地Ⅱ(8.04)>樣地Ⅳ(6.23);4個樣地的Shannon-Wiener多樣性指數(H')排序為:樣地Ⅰ(3.50)>樣地Ⅲ(3.26)>樣地Ⅱ(3.01)>樣地Ⅳ(2.46);Berger-Parker優勢度指數(D)排序為:樣地Ⅳ(0.44)>樣地Ⅱ(0.24)>樣地Ⅰ(0.20)>樣地Ⅲ(0.15);Pielou均勻度指數(J)排序為:樣地Ⅲ(0.87)>樣地Ⅰ(0.81)>樣地Ⅱ(0.80)>樣地Ⅳ(0.71)。表明蛛蜂物種數和個體數隨樣地的植被類型變化而變化,樣地Ⅰ農田植被的蛛蜂物種數和個體數均較豐富,樣地Ⅲ茶園生境各種類的蛛蜂分布較均勻,無明顯的蛛蜂優勢種,樣地Ⅳ的落葉闊葉林生境中有著明顯的優勢物種。

表3 不同生境蛛蜂群落多樣性特征Table 3 Community characteristics of spider wasp communities in different habitats
2.3 不同生境蛛蜂群落相似性分析結果
如表4所示,樣地Ⅰ與樣地Ⅱ的相似性系數為0.39,為中等不相似;其他樣地之間的相似性系數均≤0.25,表明其他樣地的蛛蜂群落間均為極不相似。樣地Ⅰ與樣地Ⅱ間的蛛蜂群落相似性相對最高,樣地Ⅲ與樣地Ⅳ的次之,且距離越遠相似性越低。4個樣地的蛛蜂群落均不相似,不同樣地的蛛蜂群落結構特點不同,說明蛛蜂群落結構及群落特征與生態環境密切相關。
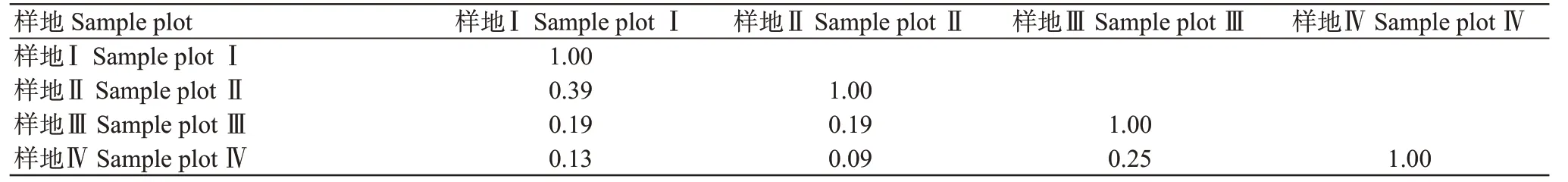
表4 4種不同生境蛛蜂群落相似性系數Table 4 Similarity coefficient of spider wasp community in four different habitats
2.4 不同季節蛛蜂優勢類群個體數量分析結果
西雙版納地區氣候一年三季,分別為:干熱季(3—5月)、濕熱季(6—10月)、霧涼季(11—次年2月)(朱華等,2015)。奧溝蛛蜂屬和扁腹溝蛛蜂屬為4種生境下的優勢類群。由圖1和圖2可看出,2個優勢類群的個體數量在4個樣地中均表現為干熱季最多、濕熱季和霧涼季數量相對較少;奧溝蛛蜂屬在樣地Ⅰ中個體數量霧涼季比濕熱季多,其余樣地的奧溝蛛蜂屬和扁腹溝蛛蜂個體數量的季節變化均是濕熱季大于霧涼季,在霧涼季大量減少,甚至下降至接近0的水平。說明氣候變化對蛛蜂的群落結構產生影響,蛛蜂更適應高溫和低濕環境的干熱季,而濕度較大的濕熱季和溫度較低的霧涼季環境中蛛蜂個體數大量減少。
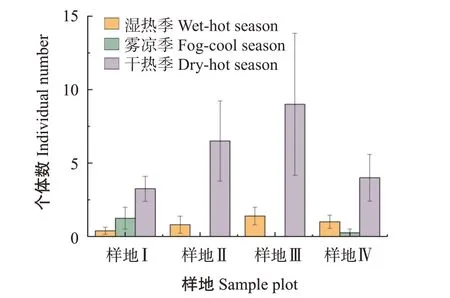
圖1 4個樣地中奧溝蛛蜂屬個體數量Fig.1 Individual numbers of Auplopus in four sample plots
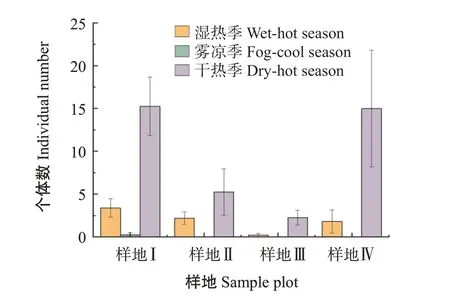
圖2 4個樣地中扁腹溝蛛蜂屬個體數量Fig.2 Individual numbers of Minagenia in four sample plots
3 討論
3.1 不同生境蛛蜂群落結構
不同生境蛛蜂群落結構不同,主要體現在蛛蜂物種數量、個體數量和特殊物種的差異上。農田生境中蛛蜂的屬和個體數量均多于其他生境,茶園與農田交界區蛛蜂個體數量最少,次生落葉闊葉林區中蛛蜂物種數量最少;蛛蜂的屬和個體數在不同生態區分布不均勻,蛛蜂不同屬之間個體數量差異較大。蛛蜂捕獵蜘蛛,是一類捕食性天敵昆蟲,這種高度的不均勻性或與農業生產中暴發的蟲害有關。人為干擾會改變生態環境中植被的多樣性,而植被多樣性是影響蛛蜂多樣性的重要原因。在田間調查中發現,農田區域環境受耕作等人為干擾程度相對較大,人們會選擇種植種類豐富的農作物,農田生境中各類植食性昆蟲相對農田與茶園交界區、茶園區和次生落葉闊葉林區環境均較豐富,其獵物蜘蛛相對較多,蛛蜂種類和個體數量也相對較大,茶園生境中蛛蜂物種數僅次于農田生境,與宋揚等(2014)關于優勢類群個體及其所占比例在一定程度上反映人為干擾程度的結論基本相符。另外,本次調查未采集到蛛蜂稀有類群,稀有類群在一定程度上反映了所在生境保護的必要性(徐正會等,2001)。
3.2 不同生境蛛蜂群落多樣性
生態環境中多樣的植被能影響蛛蜂的群落多樣性,農田生態環境中蛛蜂群落穩定性較高,結構較復雜,對外來干擾適應能力較強;茶園區蛛蜂種類分布較均勻,無明顯的優勢物種;次生林區蛛蜂物種數量少,存在明顯的優勢種。這種人為干擾越嚴重、蛛蜂多樣性及豐富度越高的格局與左自途等(2008)、張紅葉等(2011)、Chen等(2011)、李欣蕓等(2020)、王金秀等(2021)的研究報道相似,但與李志剛等(2010)、徐志峰等(2020)的研究結果存在差異,分析其原因為:蛛蜂是一類傳粉昆蟲,生態環境中植物種類尤其是顯花植物越豐富、傳粉昆蟲種類也越豐富(Senapathi et al.,2021)、種群數量也更大,該生境中物種多樣性和物種多度就越高(Zamorano et al.,2020)。當生態環境中顯花植物種類越單一,該生境中傳粉昆蟲的種類及數量也會變少,則相對容易產生優勢物種(韋小平等,2020)。同時,蛛蜂作為一類捕食性天敵昆蟲具有重要的生態功能。有研究認為多樣的作物種植能顯著增加天敵昆蟲的數量以抑制植食性昆蟲的暴發(尤士駿等,2019;Skarbek et al.,2020)。相對于單一化的種植模式,多樣化的種植能夠為天敵昆蟲提供更好的生存條件,能在多個時段為天敵昆蟲提供多種多樣的食物資源并增強其繁殖能力,并且當主要害蟲減少時,有可替代的食物源使得天敵繼續留在生境內(董文霞等,2016)。
3.3 不同生境蛛蜂群落相似性
本研究結果顯示,一個生境會與周圍生境共有部分的蛛蜂種類,但4個生境中蛛蜂群落均不相似,且生境之間距離越遠,相似水平越低;相似的物種大多集中于當地的優勢類群、常見類群及較常見類群,較稀有類群則少量單一分布于部分生境;4種生境均不是孤立的生境,各個生境中都有其他生境相似或相同的植被,相近的生境之間蛛蜂種類交流相對較多,蛛蜂相似性較高。相近的不同生境會成為彼此的天敵蟲源地(楊屾等,1999;張鑫等,2012),其生境內的昆蟲個體會擴散到周圍環境中,從而增加臨近地區的物種豐富度。半自然的棲息地能為周圍農田的天敵昆蟲提供越冬場所或食物(Rand et al.,2006)。雖然不同樣地不同生境下的蛛蜂群落各有特點,但距離相對接近的農田生境及農田邊界的茶園生境、茶園生境及次生林生境中的蛛蜂群落相似性較高,表明蛛蜂的活動能力強,能在相鄰或相似的生境中擴散活動。因此,在傳粉昆蟲的保護和利用方面,可采取在耕作區域周圍栽種顯花或誘集植物來對蛛蜂等傳粉昆蟲進行保護和補充。
3.4 不同季節蛛蜂優勢類群個體數量
蛛蜂的種群動態受當地季節變化(溫度、濕度等氣候因素)影響。西雙版納獨特的氣候綜合作用于當地蛛蜂群落,2個優勢蛛蜂類群奧溝蛛蜂屬和扁腹溝蛛蜂屬的種群動態均形成了獨特的變化特點。西雙版納一年中干濕季分明,一年可分為干熱、濕熱、霧涼3季。干熱季氣溫較高、降水量少、濕度低;濕熱季氣溫高、降水量多、濕度高;霧涼季降水較少、溫度較低、早晚有濃霧(朱華等,2015)。高溫低濕的季節有利于蛛蜂的生長繁殖,導致降水少和溫度較高的干熱季蛛蜂的個體數較其他季節多,降水多但溫度較高的濕熱季次之,降水少且溫度低的霧涼季蛛蜂個體數最少。昆蟲是變溫動物,對體內溫度的調節能力較弱,溫度的變化會影響昆蟲的發育與繁殖(史彩華等,2017);溫度的改變會影響昆蟲的性別比例和生長發育從而影響昆蟲種群密度及動態(原建強和李欣,2008)。環境濕度會對昆蟲體內水分產生直接的負面影響,從而影響昆蟲個體發育及群體發生(常曉娜等,2008)。溫度、濕度等氣候因素通常會綜合作用于昆蟲群落,從而對昆蟲發生及種群數量產生直接影響。因此,蛛蜂能更好地適應溫度較高、濕度較低的環境,對于高濕度、低溫度的環境則不能很好適應,環境因素綜合作用于蛛蜂群落,影響蛛蜂的發生與種群數量的變化。
4 結論
西雙版納勐海縣蛛蜂屬種豐富,但不同生境中蛛蜂的群落組成和特征存在差異。蛛蜂有較強的活動能力,在相鄰的生境中共享了部分種類。蛛蜂優勢類群個體數主要隨季節的變化而變化,高溫、低濕的季節更適于蛛蜂生存。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
