山地玉米植株及田間草地貪夜蛾幼蟲的空間分布特征
2022-02-03 07:58:14王錕張麗蘭黎斌杜廣祖陳斌
南方農業學報 2022年10期
關鍵詞:玉米田
王錕,張麗蘭,黎斌,杜廣祖,陳斌*
(1云南農業大學植物保護學院/云南生物資源保護與利用國家重點實驗室,云南昆明 650201;2云南省曲靖市師宗縣植保植檢站,云南曲靖 655700)
0 引言
【研究意義】草地貪夜蛾[Spodoptera frugiperda(J.E.Smith)]隸屬于鱗翅目(Lepidoptera)夜蛾科(Noctuidae)貪夜蛾屬(Spodoptera),是一種原產于美洲的雜食性農業害蟲(Day et al.,2017)。該蟲食性廣、適生區域廣、遷飛距離遠、繁殖能力強(孔德英等,2019;雷妍圓等,2020;Xiao et al.,2020),能取食300多種植物,除為害禾本科植物玉米(Zea maysL.)(Montezano et al.,2018;洪鼎剴等,2021)、水稻(Oryza sativaL.)(王磊等,2019)、高粱[Sorghum bicolor(L.)Moench](顧偌鋮等,2020)、小麥(Triticum aestivumL.)(王芹芹等,2020)和甘蔗(Saccharum officinarumL.)(孫東磊等,2019)外,還取食為害甘藍(Brassica oleraceaL.)(劉銀泉等,2019)、馬鈴薯(Solanum tuberosumL.)(趙猛等,2019)、大豆[Glycine max(Linn.)Merr.](白一葦等,2020)、花生(Arachis hypogaeaLinn.)(何莉梅等,2020)和向日葵(Helianthus annuusL.)(吳孔明,2020)等。該蟲自2019年入侵進入云南省普洱市江城縣以來,已在我國多地成功定殖危害,并已成為影響我國玉米產業發展的重大害蟲(姜玉英等,2021),且已對多種化學農藥,如甲萘威、甲基對硫磷、毒死蜱和高效氯氟氰菊酯等產生了較強的抗藥性(李永平等,2019;Guo et al.,2020),加上該蟲能通過遠距離遷飛引起暴發成災,因而對該蟲的監測預警和防控難度較大(郭井菲等,2019;Guo et al.,2020)。昆蟲種群的空間分布和抽樣技術是昆蟲發生危害研究的重要內容,研究昆蟲的空間分布,不僅能揭示昆蟲種群結構及種群動態與環境間的關系,還有助于害蟲的預測預報及綜合防治(榮昌鶴等,2015)。因此,開展草地貪夜蛾幼蟲的空間分布特征研究,可為該害蟲的預測預報、抽樣調查和防治提供重要參考依據。【前人研究進展】種群空間分布格局研究是草地貪夜蛾研究的重要內容(Hernandez-Mendoza et al.,2008;孫小旭等,2019;楊現明等,2020a,2020b;劉詩穎等,2021)。國外Farias等(2001)報道玉米苗期草地貪夜蛾低齡幼蟲呈聚集分布而高齡幼蟲趨向于分散分布;Melo等(2006)報道巴西玉米田草地貪夜蛾幼蟲呈隨機分布;Serra和Trumper(2006)報道阿根廷玉米田草地貪夜蛾幼蟲總體上呈聚集分布;Hernandez-Mendoza等(2008)報道墨西哥玉米田草地貪夜蛾幼蟲呈隨機分布。國內胡慧芬等(2019)對云南飼料玉米田草地貪夜蛾幼蟲空間分布型進行了調查研究,發現在飼料玉米田該蟲幼蟲呈典型的聚集分布;孫小旭等(2019)在云南普洱市江城縣鮮食玉米田調查發現,鮮食玉米田草地貪夜蛾幼蟲呈聚集分布,且聚集度隨密度的增加而升高;馬明勇等(2019)報道湖南玉米苗期草地貪夜蛾幼蟲呈聚集分布;楊紫涵等(2020)報道廣東玉米田玉米苗期草地貪夜蛾幼蟲整體上呈聚集分布;楊現明等(2020a,2020b)在云南省大麥和小麥田調查發現,大麥田草地貪夜蛾幼蟲呈聚集分布,而在小麥田中其低齡幼蟲呈聚集分布,高齡幼蟲呈均勻分布;劉詩穎等(2021)報道海南玉米田草地貪夜蛾3齡幼蟲呈聚集分布,5~6齡幼蟲呈均勻分布。綜合以上國內外玉米田草地貪夜蛾幼蟲空間分布型的研究報道,發現草地貪夜蛾幼蟲在玉米田的空間分布特征隨著地理環境與氣候、玉米種植制度及玉米品種類型的不同而存在差異,且即使在同一省范圍內,不同區域玉米田草地貪夜蛾幼蟲的空間分布特征也不相同,如馬明勇等(2019)在湖南調查發現,永州、長沙和岳陽3地玉米田草地貪夜蛾幼蟲空間分布格局均為聚集分布;劉詩穎等(2021)研究發現,草地貪夜蛾幼蟲在海南東部地區玉米田為聚集分布,在中部、北部、南部玉米田的空間分布為均勻分布。【本研究切入點】云南省曲靖市師宗縣位于云南省東南部山區,是云南省玉米主要產區,草地貪夜蛾于2019年4月進入龍慶鄉,已在該區成功定殖,嚴重為害玉米并造成了巨大的經濟損失(張瓊等,2020)。作為一種新入侵進入該地區的害蟲,目前對其在該區域內的發生危害特點及空間分布特征尚不清楚,影響了對該蟲的科學預測預報及綜合防治。【擬解決的關鍵問題】采用聚集指標、Iwao回歸分析法和Taylor冪法則等方法,研究云南山地玉米種植制度下草地貪夜蛾幼蟲的空間分布類型及抽樣技術,為山地玉米田草地貪夜蛾的調查取樣、監測預警及選擇防治適期提供指導,也為提高草地貪夜蛾的防控效果、降低其危害提供理論依據。
1 材料與方法
1.1 調查地點
調查地位于云南省曲靖市師宗縣龍慶鄉扯寨村委會大山腳村(海拔1340.25 m,東經104°4′54″,北緯24°29′28″)。玉米田為山地玉米田,土壤為粘土。
1.2 調查作物
試驗田前茬作物為小麥,玉米于2021年5月28日播種,株行距為30 cm×40 cm。玉米品種為師單8號。整個生育期間未施用任何殺蟲劑。
1.3 試驗方法
選擇玉米田3塊,每塊田面積400 m2。從2021年6月17日玉米齊苗后開始,每塊田選取5個小樣方,采用W型五點調查取樣法(趙雪晴等,2019),在每個小樣方內定點定株標記玉米并分別調查10株。每3 d調查1次,如遇下雨則調查時間順延。每次分別調查記錄玉米不同生育期(齊苗期、拔節期、小喇叭口期、大喇叭口期、抽雄期、散粉期、吐絲期、灌漿期、乳熟期和蠟熟期)與不同組織(心葉、莖/葉、雌穗和雄穗)草地貪夜蛾幼蟲數量及齡期(1~3齡為低齡,4~6齡為高齡),如齊苗期到抽雄期,調查玉米心葉和莖葉(展開葉)的正面和背面;玉米進入吐絲期到蠟熟期后調查玉米的雌穗和雄穗,同時計算雌穗上的百穗蟲量。
1.4 相關計算方法及公式
采用Taylor冪法則、Iwao回歸分析法及聚集度指標法(孫小旭等,2019)對草地貪夜蛾幼蟲空間分布型進行分析。
1.4.1 聚集度指標.每個樣本每次調查玉米植株上草地貪夜蛾幼蟲數量,計算每個樣本方差(S2)和蟲口密度(m),再根據公式計算其空間分布型的各個參數值。Lloyd平均擁擠度(m*)(m*=m+S2/m-1)、Morisita的擴散指數(C)(C=S2/m)、聚塊指數(m*/m),3個值分別小于1時種群為均勻分布,等于1時為隨機分布,大于1時為聚集分布;David & Moor的叢生指數(I)(I=S2/m-1)、Kuno的久野指數(Ca)[Ca=(I=S2/m-1)/m]、Water的負二項分布(K)[K=m(/S2/m-1)],3個值分別等于0時為隨機分布,小于0時為均勻分布,大于0時為聚集分布。
1.4.2 Iwaom*-m回歸分析法Iwao回歸方程:m*=α+βm。式中,α為個體,β為個體群的分布情況;當β<1時種群呈均勻分布,β趨近于1時種群呈隨機分布,β>1時種群呈聚集分布。Taylor冪法則:lgS2=lga+blgm,其中,b值為聚集度指標,當b<1時種群呈均勻分布,b趨近于1時呈隨機分布,b>1時呈聚集分布。
1.4.3 抽樣技術.理論抽樣數計算模型:
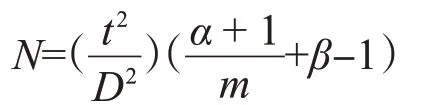
式中,N為最適的理論抽樣數;t為在一定置信度下的分布值;D為允許誤差,α為Iwaom*-m回歸方程中的截距,β為斜率。
復合序貫抽樣技術模型:
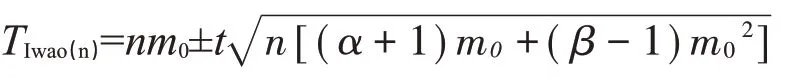
式中,用減號計算出下限T2(n)的值,用加號計算出上限T1(n)的值,m0為防治指標,n為田間抽樣數,t為相應置信度下的分布值,α、β同理論抽樣計算模型。若取樣時蟲量大于上限表示種群密度高于防治指標,蟲量小于下限表示種群密度低于防治指標,需繼續進行抽樣。
1.5 統計分析
使用Excel 2010對試驗數據進行整理分析,利用DPS 9.01對試驗數據進行單因素方差分析,Duncan’s新復極差法比較不同處理間的差異顯著性,利用DPS 9.01進行空間分布格局分析。
2 結果與分析
2.1 草地貪夜蛾幼蟲在玉米植株上的分布特征
2.1.1 玉米不同生育期草地貪夜蛾不同齡期幼蟲的組成結構.由表1可看出,不同生育期玉米植株上草地貪夜蛾各齡期幼蟲的組成不同。在玉米齊苗期、拔節期、大喇叭口期和吐絲期,草地貪夜蛾低齡幼蟲(1~3齡)數量比例較高,分別為100.00%、71.45%、63.39%和78.07%,其中2齡幼蟲數量比例較高,分別占所有幼蟲數量的82.02%、38.20%、42.51%和51.56%。抽雄期、散粉期、灌漿期、乳熟期和蠟熟期草地貪夜蛾高齡幼蟲(4~6齡)數量比例較高,分別為70.01%、83.55%、81.35%、95.94%和95.56%。此外,在玉米小喇叭口期,除1齡幼蟲比例占2.61%外,2~6齡幼蟲數量比例均在20.00%左右。在乳熟期和蠟熟期,6齡老熟幼蟲占優勢(57.48%和62.22%)。
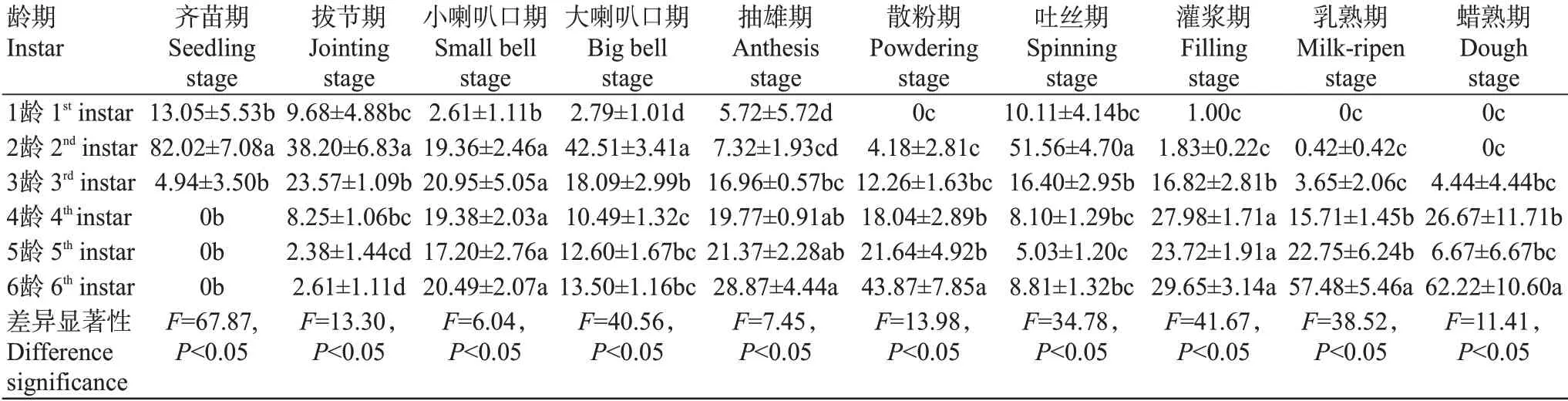
表1 玉米不同生育期草地貪夜蛾各齡期幼蟲數量比例(%)Table 1 Proportion of larvae S.frugiperda of various instars at the different corn growth stages(%)
2.1.2 玉米植株不同部位草地貪夜蛾幼蟲的分布表2結果顯示,除草地貪夜蛾1齡幼蟲在雄穗上沒有分布外,各齡期幼蟲在玉米不同部位上均有分布;除1齡幼蟲外,各齡期幼蟲在玉米心葉、莖/葉、雄穗和雌穗等不同部位間的數量比例均存在顯著差異(P<0.05,下同),其1~6齡幼蟲主要分布于玉米心葉和雌穗上,數量比例均超過24.00%;1~3齡幼蟲在莖/葉上的數量比例均超過20.00%,分別為25.26%、21.85%和20.53%,而4~6齡幼蟲在莖/葉上的數量比例均低于20.00%;1~6齡幼蟲在雄穗上的數量比例均低于20.00%。玉米植株上不同部位草地貪夜蛾幼蟲總的數量分布為:心葉>雌穗>莖/葉>雄穗。
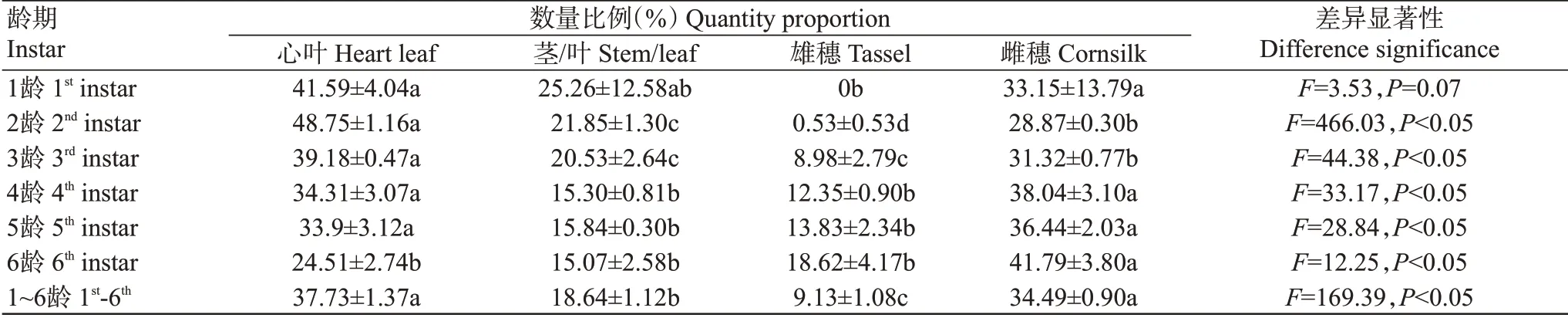
表2 草地貪夜蛾幼蟲在玉米植株不同部位上的分布Table 2 Distribution of larvae S.frugiperda on different parts of corn plants
2.1.3 玉米雌穗上草地貪夜蛾種群密度由玉米不同生育期雌穗上草地貪夜蛾幼蟲的種群密度調查結果(表3)可看出,除1齡幼蟲外,各齡期幼蟲在玉米不同生育期雌穗上的百穗蟲量均存在顯著差異。幼蟲在雌穗上的百穗蟲量隨著玉米的發育逐漸增多,至灌漿期達到峰值,灌漿期后,隨著玉米雌穗籽粒的成熟,百穗蟲量逐漸減少,到乳熟期時降至42.22頭/100穗,到蠟熟期僅為2.56頭/100穗。表明在玉米雌穗上灌漿期的草地貪夜蛾幼蟲數量最多。
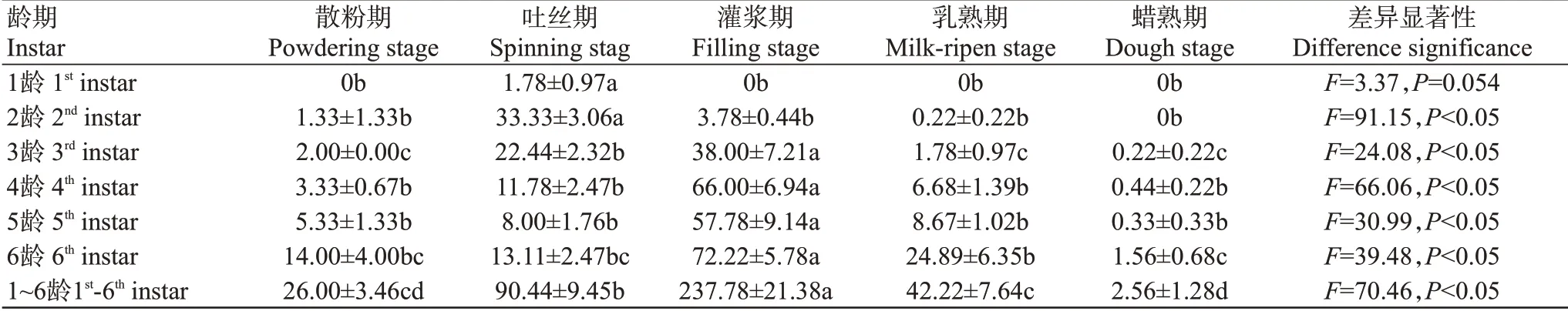
表3 玉米雌穗上草地貪夜蛾蟲口密度(頭/100穗)Table 3 Density of larvae S.frugiperda on corn ears(S.frugiperda/100 ears)
2.2 山地玉米田草地貪夜蛾幼蟲的空間分布格局
2.2.1 低齡幼蟲的空間分布型.由表4可知,玉米在齊苗期、拔節期、大喇叭口期和吐絲期時的低齡幼蟲蟲口密度較大,平均密度(m)均大于1.00頭/株,其余生育期的低齡幼蟲蟲口平均密度均小于1.00頭/株。低齡幼蟲在玉米各個生育期的聚集指標中大于1的有平均擁擠度(m*)、聚塊指數(m*/m)、久野指數(Ca)和擴散指數(C);大于0的叢生指數(I)和負二項分布(K)。綜合各聚集度分布標準分析表明,草地貪夜蛾低齡幼蟲在山地玉米田間的空間分布類型為聚集分布。根據Iwao的回歸分析方法,建立m與m*的線性回歸方程為m*=2.26+1.28744m(R=0.62),由方程可知,α=2.26>1及β=1.29>1,說明低齡幼蟲種群呈聚集分布。將Taylor冪法則公式S2=amb轉換成lgS2=lga+blgm,并將表4中的S2和m值代入公式得到回歸方程lgS2=1.18lgm+0.55(R=0.9720),a=0.55>0,b=1.18>1。聚集度指標檢驗結果與Iwao回歸分析結果相一致。因此,可判定草地貪夜蛾低齡幼蟲在山地玉米田間呈聚集分布,且聚集度存在密度依賴性。
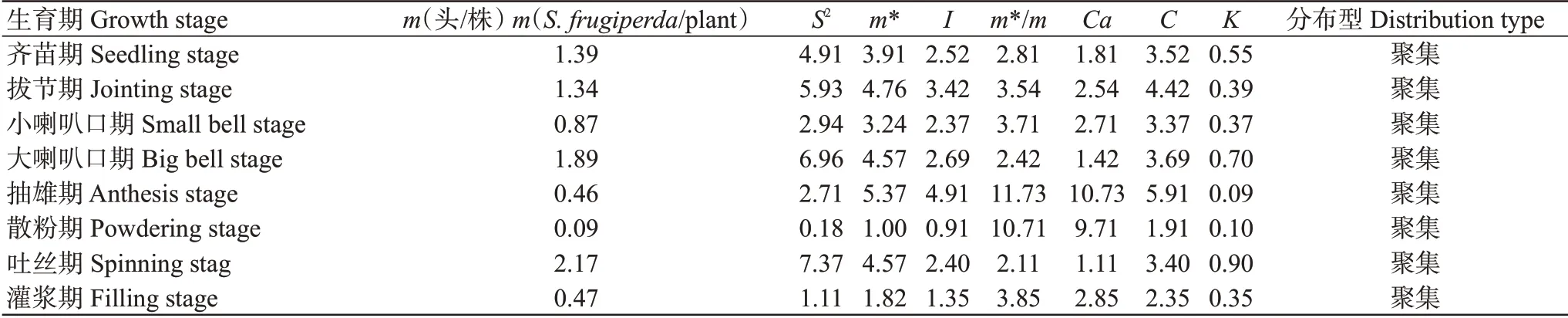
表4 玉米田草地貪夜蛾低齡幼蟲密度及聚集指標Table 4 Density and aggregation index of young larvae of S.frugiperda in corn fields
2.2.2 高齡幼蟲的空間分布型.由表5可知,玉米生育期為小喇叭口期、大喇叭口期、抽雄期、灌漿期時高齡幼蟲的蟲口密度較大,m均大于1.00頭/株;拔節期、散粉期和吐絲期時高齡幼蟲的蟲口m均小于1.00頭/株;苗期未發現有高齡幼蟲。高齡幼蟲在拔節期和吐絲期的m*/m、C和K等聚集指標均大于1;灌漿期的m*也大于1;而大喇叭口期的m*=1;其余各生育期的聚集指標均小于1。綜合各聚集度分布標準分析表明,草地貪夜蛾高齡幼蟲在山地玉米田間的空間分布類型主要為均勻分布。根據Iwao的回歸分析方法,利用m與m*建立線性回歸方程m*=0.01+0.94m(R=0.98),獲得α=0.01<1及β=0.94<1,說明高齡幼蟲種群呈均勻分布。將Taylor冪法則公式S2=amb轉換成lgS2=lga+blgm,并將表5中的S2和m值代入公式得到回歸直線方程lgS2=1.18lgm+0.55(R=0.9825),a=-0.03<0,b=0.93<1。聚集度指標檢驗結果與Iwao回歸分析結果一致。因此,判定草地貪夜蛾高齡幼蟲在山地玉米田間主要呈均勻分布。
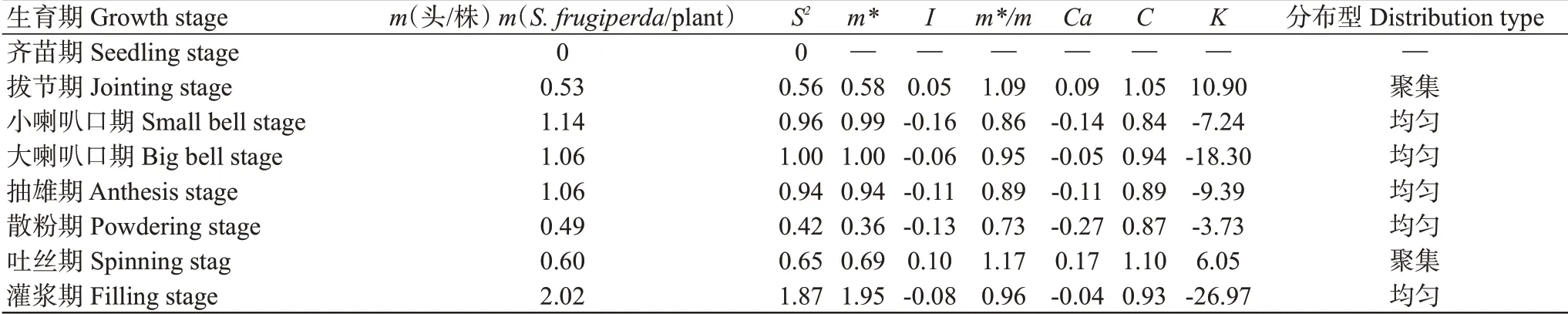
表5 玉米田草地貪夜蛾高齡幼蟲密度及聚集指標Table 5 Density and aggregation index of aged larvae of S.frugiperda in corn fields
2.3 山地玉米田草地貪夜蛾幼蟲的空間分布型
由表6結果可得,草地貪夜蛾幼蟲在山地玉米散粉期的各聚集指標均小于1,其余各生育期的m*、m*/m和C均大于1;小喇叭口期和灌漿期的I及齊苗期和拔節期的K均小于1,其余各生育期的I和K均大于1;此外,Ca僅在齊苗期和拔節期大于1。綜合上述結果表明,草地貪夜蛾幼蟲在山地玉米田間的空間分布呈聚集分布。根據m與m*進行回歸模型分析,建立的線性回歸方程為m*=0.47+1.34m(R=0.82),獲得α=0.47,β=1.34>1。將Taylor冪法公式S2=amb轉換成lgS2=lga+blgm,并將表6中的S2和m值代入公式得到回歸直線方程lgS2=1.39lgm+0.21(R=0.93),a=0.21>0,b=1.39>1。聚集度指標檢驗結果與Iwao回歸分析結果一致,表明草地貪夜蛾幼蟲在山地玉米田間的空間分布呈聚集分布,且聚集度具密度依賴性。
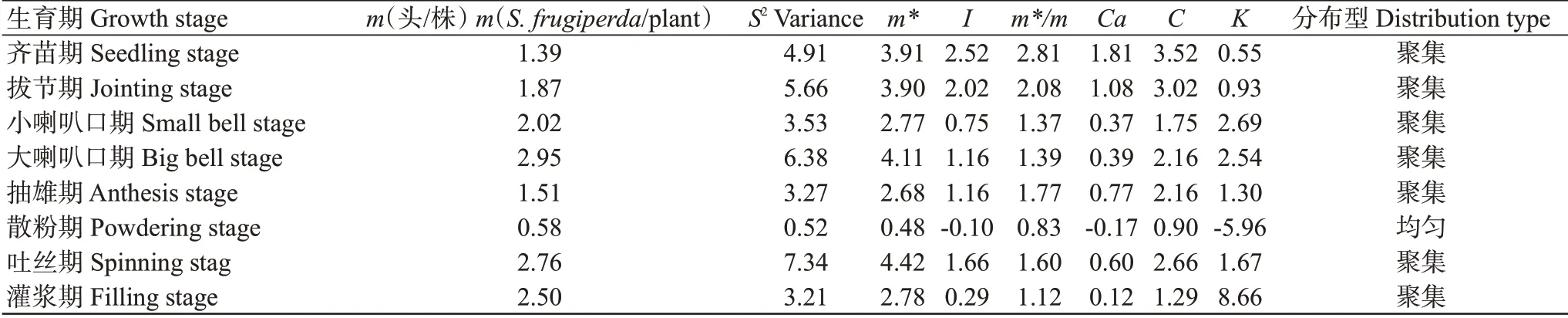
表6 玉米田草地貪夜蛾幼蟲密度及聚集指標Table 6 Density and aggregation index of S.frugiperda larvae in corn fields
2.4 山地玉米田草地貪夜蛾幼蟲的理論抽樣模型及技術
將山地玉米田草地貪夜蛾幼蟲低齡、高齡以及全部樣本Iwaom*-m回歸模型系數α=2.26,β=1.29;α=0.01,β=0.94;α=0.47,β=1.34分別代入公式,得到最佳理論抽樣數模型(條件:概率為0.95,t=1.96)。草地貪夜蛾低齡幼蟲為:;高齡幼蟲為;全部幼蟲樣本為由各個最佳理論抽樣數模型公式可計算出不同誤差條件下(D=0.1,0.2,0.3)草地貪夜蛾低齡、高齡及全部幼蟲樣本在相應密度(m)的理論最佳抽樣數(N)。由表7可知,在允許誤差固定時,低齡、高齡及全部幼蟲樣本的理論抽樣數均隨著幼蟲平均密度的增大而呈減少的趨勢,且減少的幅度逐漸縮小。此外,在幼蟲平均密度固定時,低齡、高齡及全部幼蟲樣本的理論抽樣數也隨著允許誤差的增大而減少,減少的幅度亦相應逐漸縮小。
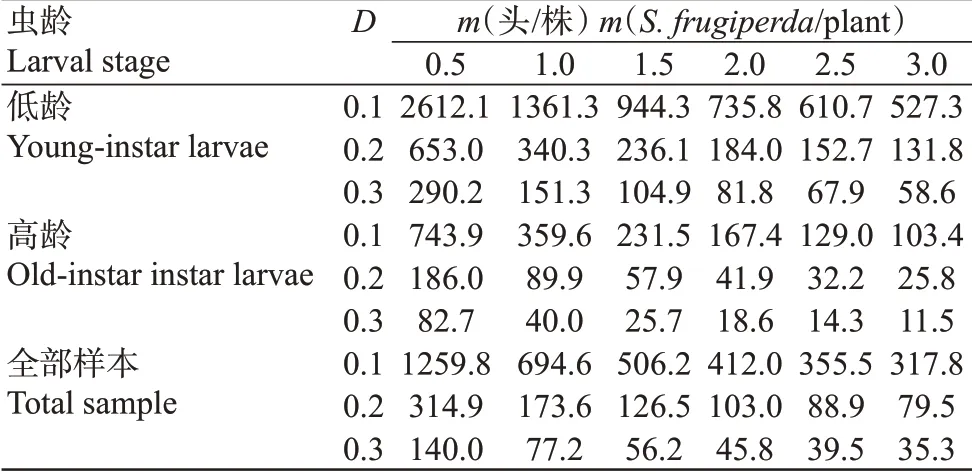
表7 玉米田草地貪夜蛾低齡、高齡及全部幼蟲樣本不同密度下的理論抽樣數Table 7 Theoretical sampling number of young-instar,old-instar and all larvae of S.frugiperda in corn field under different densities
根據前人研究結果,玉米田間草地貪夜蛾幼蟲的為害率達0.33頭/株時就應采取防控措施(孫小旭等,2019)。將草地貪夜蛾幼蟲低齡、高齡及全部樣本的Iwaom*-m回歸模型系數α=2.26,β=1.29;α=0.01,β=0.94;α=0.47,β=1.34分別代入序貫抽樣模型,獲得低齡幼蟲的序貫 抽樣模型為:TIwao(n)=0.33n±1.96;高齡幼蟲的序貫抽樣模型為:TIwao(n)=;全部幼蟲樣本的序貫抽樣模型為
計算不同抽樣株數(n=60、70、80、…、200)時將幼蟲的序貫抽樣區間分別列入表8。抽樣調查時,若累積的幼蟲數高于表中上限的數量T1(n)時,表明種群密度高于防治指標,需要采取防控措施;而累積的幼蟲數低于下限的數量T2(n)時,表明種群密度低于防治指標,不需要防治;若累積的幼蟲數在上、下限的數量之間時,表明還要繼續抽樣。
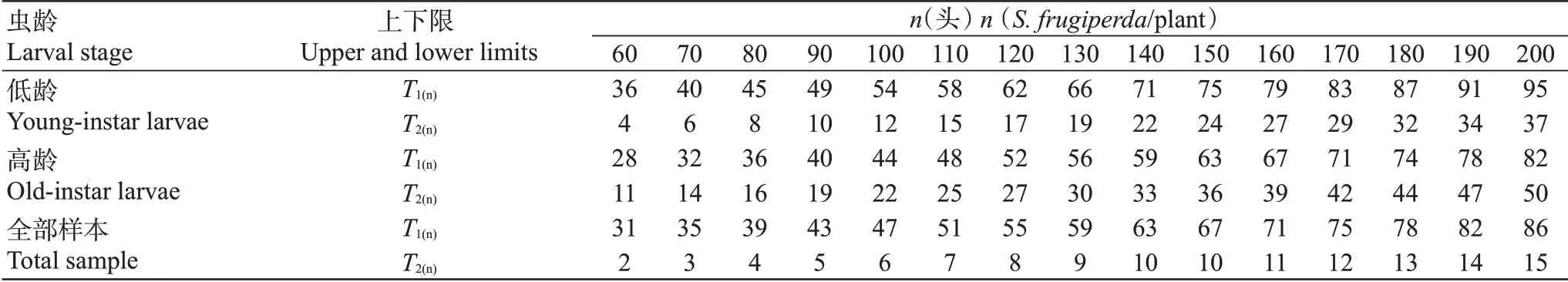
表8 玉米田草地貪夜蛾低齡、高齡及全部幼蟲樣本的序貫抽樣數Table 8 Sequential sampling number of young-instar,old-instar and all larvae of S.frugiperda in corn field
3 討論
草地貪夜蛾自2019年1月進入我國云南南部普洱市江城縣以來即迅速擴散,現已成為危害我國玉米生產的重要因素(姜玉英等,2019)。本研究對云南省曲靖市師宗縣山地玉米田玉米植株上草地貪夜蛾幼蟲的分布特點進行調查研究,發現在山地玉米的整個生育期,草地貪夜蛾幼蟲在玉米植株上的分布表現為心葉>雌穗>莖/葉>雄穗,表明在玉米生育期該蟲的危害部位是心葉和雌穗,這可能也是該蟲危害造成重大損失的重要原因之一。各生育期玉米植株上草地貪夜蛾各齡期幼蟲的組成不同,玉米齊苗期、拔節期、大喇叭口期和吐絲期草地貪夜蛾低齡蟲的數量比例較高,與草地貪夜蛾成蟲以卵塊形式產卵的習性相吻合;在抽雄期、散粉期、灌漿期、乳熟期和蠟熟期草地貪夜蛾高齡幼蟲數量比例較高,綜上得到草地貪夜蛾幼蟲在玉米田間取食具有趨嫩性,與唐慶峰等(2020)研究發現玉米雌穂及心葉等幼嫩器官和組織對草地貪夜蛾具有較高的適合度的結論相符。因此,應加強對玉米苗期草地貪夜蛾的預測預報,在苗期加強對低齡幼蟲的防治,以減輕對苗期的危害和植株生長發育的影響;同時,應加強對玉米心葉和雌穗中草地貪夜蛾的防治,以提高防治效果。本研究調查發現,玉米雌穗吐絲后,草地貪夜蛾幼蟲即可危害玉米花絲,花絲部分或全部被咬斷,之后隨花絲通道進入穗尖,取食籽粒,導致其啃食后的玉米籽粒發生霉變,應加強對玉米吐絲前草地貪夜蛾的監測和預防,以減少進入玉米雌穗的幼蟲數量,從而減少對玉米雌穗與籽粒的危害,與韓海亮等(2020)研究發現草地貪夜蛾幼蟲拔節前主要為害心葉,吐絲后主要為害雌穗,防治關鍵期為苗期保苗和后期保穗的結論相吻合。
本研究發現,山地玉米田草地貪夜蛾低齡幼蟲呈聚集分布,與Farias等(2001)和阿根廷學者Serra和Trumper(2006)的研究結果一致,但與巴西學者Melo(2014)和墨西哥學者Hernandez-Mendoza(2008)報道的玉米田草地貪夜蛾幼蟲呈隨機分布的結論存在差異,可能由于地理氣候、土壤類型、玉米品種及種植方式與本研究不同所致。本研究結果與楊紫涵等(2020)在廣州研究報道的玉米苗期和喇叭口初期草地貪夜蛾幼蟲為聚集分布,而大喇叭口期為均勻分布的結果一致,究其原因可能與草地貪夜蛾習性和寄主植物的發育特點相關。本研究發現,在山地玉米田間,草地貪夜蛾低齡幼蟲的空間分布型呈聚集分布,而高齡幼蟲呈均勻分布,與國內馬明勇等(2019)在湖南、孫小旭等(2019)在云南德宏州、劉詩穎等(2021)在海南的玉米田草地貪夜蛾幼蟲呈聚集分布的結果也存在差異,可能與本研究中山地玉米生境特征及玉米品種有關,但具體原因還有待進一步探究。但該結果與楊現明等(2020a)在小麥田研究發現的草地貪夜蛾低齡幼蟲呈聚集分布,高齡幼蟲為均勻分布的結果一致,雖然寄主植物不同,但在不同的寄主植物上草地貪夜蛾高齡幼蟲均具有互相殘殺的習性,可能由于這種習性導致高齡幼蟲在玉米田呈均勻分布的空間分布特征。此外,本研究發現低齡幼蟲與高齡幼蟲的空間分布型存在差異的特點,可為生產防治提供有利條件,如在幼蟲低齡時應用藥劑集中防治可取得更好的防效。
抽樣技術是指導開展害蟲監測與防治工作的基礎(孫小旭等,2019;楊紫涵等,2020)。本研究通過對云南山地玉米田草地貪夜蛾幼蟲空間分布調查及抽樣技術的研究,建立了理論抽樣數模型和序貫抽樣模型。在田間調查過程中,防治標準可根據序貫抽樣技術來確定。當調查樣本的累積幼蟲量T0(n)大于公式計算的蟲量上限,說明種群密度已高于防治指標,需要進行防治;當低于下限時,則不需要防治;若處于上、下限之間,仍需要繼續抽樣,直至最大抽樣數,再按照序貫抽樣表調查蟲量,決定是否需要噴藥防治。理論抽樣數模型和序貫抽樣模型可對山地玉米田草地貪夜蛾的防治提供有效的參考依據。
昆蟲空間分布型受多種因素的影響,如地理環境、寄主品種、寄主生長發育階段、昆蟲種群密度和種群內部競爭等(Serra and Trumper,2006;Melo et al.,2014;楊紫涵等,2020)。云南省曲靖市師宗縣龍慶鄉是云南省玉米種植重要區域之一,地形均為山地,田塊連片種植少,且農戶自主種植玉米品種較多,草地貪夜蛾已在山地玉米田間定殖、發生和危害,已成為該地區玉米生產的重要害蟲之一。本研究僅從玉米師單8號及該品種的不同生育期和不同部位研究了草地貪夜蛾幼蟲的空間分布特征,并未開展幼蟲在不同氣候條件、土壤、種植密度、玉米多品種等各因素下的空間分布型研究,本研究得到的各種模型未必能適用于指導其他玉米種植區的害蟲監測與防治。因此,在今后的研究中應加強分析草地貪夜蛾幼蟲在更多因素條件下的空間分布特征及抽樣調查技術,對已得到的模型進行驗證或優化,以發展適合當地的玉米田草地貪夜蛾的抽樣調查技術,從而更好地指導草地貪夜蛾防控。
4 結論
云南山地玉米田草地貪夜蛾低齡幼蟲呈聚集分布,高齡幼蟲呈均勻分布,整個生育期田間草地貪夜蛾幼蟲總體呈聚集分布。可根據模型計算山地玉米田草地貪夜蛾幼蟲最佳理論抽樣數和序貫抽樣區間,以提高預測預報的準確性。
猜你喜歡
現代農業科技(2016年23期)2017-04-06 21:38:29
現代農業(2016年4期)2016-02-28 18:42:10
雜草學報(2015年2期)2016-01-04 14:57:53
江蘇農業科學(2015年9期)2015-10-20 14:37:12
營銷界(2015年23期)2015-02-28 22:06:18
營銷界(2015年23期)2015-02-28 22:06:17
現代農業(2015年5期)2015-02-28 18:40:47
現代農業(2015年5期)2015-02-28 18:40:42
現代農業(2015年3期)2015-02-28 18:40:23
安徽農學通報(2014年13期)2014-09-19 08:57:03
