氨氮慢性脅迫對凡納濱對蝦肝胰腺氨代謝相關指標及細胞凋亡的影響
2022-02-03 07:57:52黃勇戴習林
南方農業學報 2022年10期
黃勇,戴習林*
(1水產科學國家級實驗教學示范中心(上海海洋大學),上海 201306;2農業農村部淡水水產種質資源重點實驗室(上海海洋大學),上海 201306)
0 引言
【研究意義】自20世紀90年代以來,凡納濱對蝦(Litopenaeus vannamei)產業迅速發展,已成為我國最重要的水產養殖動物之一(滕瑜等,2021)。凡納濱對蝦對環境的適應性強,具備廣溫廣鹽的生理特性,加上養殖技術的進步,養殖范圍已擴展到內陸低鹽地區。氨氮來自含氮有機物的分解,以NH4+和NH3的形態存在,NH3毒性較強;在集約化養殖過程中,受溫度、pH、水交換條件等的限制,水體氨氮濃度難以穩定在較低水平,使甲殼類受到氨氮脅迫,導致組織損傷,免疫力下降,影響甲殼類的生長發育和存活(Koo et al.,2005;Miranda-Filho et al.,2009;黃姝,2017)。因此,研究氨氮慢性脅迫對凡納濱對蝦的影響有助于凡納濱對蝦產業的健康發展。【前人研究進展】氨氮脅迫會擾亂甲殼類的機體代謝,降低免疫力,對組織造成損傷,使其更易被病原所侵襲(葛紅星等,2014)。熊大林等(2020)研究表明,經15 mg/L氨氮脅迫72 h后,凡納濱對蝦鰓組織邊界模糊,鰓絲松散,細胞分解,鰓組織損傷導致生理功能紊亂。方金龍等(2017)在無脅迫和15.6 mg/L氨氮脅迫下,分別對凡納濱對蝦注射白斑綜合征病毒(WSSV)粒子,通過熒光定量PCR檢測白斑綜合征病毒VP28基因表達量,結果顯示72 h后氨氮脅迫組顯著高于無脅迫組,導致WSSV增殖更快的原因在于氨氮脅迫會降低凡納濱對蝦的免疫酶活性。彭軍輝等(2018)研究指出,在氨氮濃度范圍0~140 mg/L對擬穴青蟹(Scylla paramamosain)進行急性毒性脅迫試驗,24和48 h時,死亡率隨氨氮濃度增加而升高;且血清中溶菌酶(LZM)活力在各實驗組中受到抑制,對病原的抵抗能力降低。當甲殼類受到環境中的病原或有毒物質的攻擊時,機體會開啟先天性免疫防御,產生大量的活性氧以抵抗病原入侵,但過量活性氧同樣會損害機體自身(于杰倫,2019)。董學興等(2019)研究表明,羅氏沼蝦(Macrobrachium rosenbergii)幼蝦在0~1.06 mg/L非離子氨脅迫48 h后,體內丙二醛(MDA)含量隨氨氮濃度增加而上升,反映出機體內產生過多的活性氧,活性氧對細胞的攻擊程度與氨氮濃度成正相關。活性氧含量過高還會刺激凡納濱對蝦細胞凋亡基因顯著表達,促進細胞凋亡發生(李玲等,2021)。王蕓等(2017)研究表明,中國對蝦(Fenneropenaeus chinensis)在8 mg/L氨氮脅迫下,血淋巴總抗氧化能力在6 h后呈降低趨勢,機體抗氧化平衡被破壞,導致細胞凋亡基因Caspase顯著上調。細胞色素C(CytC)能激活細胞凋亡途徑,最終激活凋亡效應因子Caspase-3,誘導細胞凋亡發生(Hegazi et al.,2010)。為應對氨氮脅迫,甲殼類依賴氨代謝途徑將氨氮排出體外或將氨氮代謝為尿素和谷氨酰胺等低毒、中性物質(Ip et al.,2004;周發林等,2016)。【本研究切入點】甲殼類肝胰腺功能主要是消化、吸收和儲存營養物質,對甲殼類的生長發育、抵抗病菌病毒感染和環境不利因素提供能量有重要作用,也是免疫和氨氮解毒代謝的重要器官(王小剛,2015;楊晨,2020)。在養殖中后期,養殖水體有機物大量累積,底部水體氨氮含量大幅增加,長期氨氮脅迫會對凡納濱對蝦肝胰腺造成不利影響,導致生長緩慢。前人研究氨氮對凡納濱對蝦的影響在氨代謝方面多為急性毒性試驗,缺少慢性氨氮脅迫和細胞凋亡相關試驗。【擬解決的關鍵問題】通過20和40 d氨氮慢性脅迫,比較凡納濱對蝦的生長存活、酶活性、基因表達量和TUNEL檢測結果,研究養殖中后期氨氮對凡納濱對蝦肝胰腺的影響,為凡納濱對蝦免疫機理研究和生產實踐指導提供參考。
1 材料與方法
1.1 試驗動物
試驗用凡納濱對蝦取自上海申漕特種水產開發有限公司,使用曝氣自來水和海水晶調鹽度調至3,水溫為30~31℃,溶解氧為5 mg/L,pH為8.0~8.1,暫養于水泥池中,2周后進行正式試驗。
1.2 試驗方法
通過預試驗得知,凡納濱對蝦氨氮脅迫96 h-LC50約60.0 mg/L,取該濃度的20%(12.0 mg/L)為最高濃度組。設4個氨氮濃度:0(對照組)、0.9、5.2和12.0 mg/L,對應非離子氨濃度為0、0.08、0.49和1.13 mg/L。將480尾初始體質量6.49±0.14 g的凡納濱對蝦分為4個組,6個平行。其中3個平行用于20 d采樣,另外3個平行用于40 d采樣。試驗在75.5 cm×55.0 cm×45.5 cm的養殖箱中進行,水體體積100 L,每日定時(6:00、14:00、22:00)、定量(每餐投喂量占蝦體質量1.5%)投喂,攝食1.5 h后,清出含氮廢物,參考王華等(2013)的方法確定和調整氨氮濃度,每天換水97%,使用Na2CO3溶液維持pH穩定,用NH4Cl配制成10 g/L母液,稀釋成所需濃度。于慢性氨氮脅迫20和40 d時采樣,樣品采集前一天停止投喂,各濃度組隨機選取3尾凡納濱對蝦,采集血淋巴保存于抗凝管中。將凡納濱對蝦肝胰腺組織剖出,用預冷PBS清洗再吸干后,放入離心管中,于-80℃冰箱中保存。同時取肝胰腺組織用組織固定液(波恩氏液)保存,用于制作石蠟切片和細胞凋亡染色。
1.3 生長指標和存活率測定
體質量增長率(WGR)和存活率(SR)分別按如下公式計算:
WGR(%)=(WT-W0)/W0×100
式中,WT為終末體質量,W0為初始體質量,T為時間(20和40 d)。
SR(%)=(N0-NT)/N0×100
式中,N0為初始數量,NT為試驗結束時的數量。
1.4 生理生化指標測定
采用血氨、尿素氮、谷氨酸脫氫酶(GDH)、谷氨酰胺合成酶(GS)、總超氧化物歧化酶(T-SOD)和過氧化氫酶(CAT)試劑盒(南京建成生物工程研究所)檢測血淋巴和肝胰腺生理生化指標。
1.5 基因表達量測定
采用TRIzol法提取肝胰腺RNA,天根生化科技(北京)有限公司反轉錄試劑盒進行反轉錄。在GenBank中查詢凡納濱對蝦Cyt C(KX096890.1)、Caspase-3(EU421939.1)和β-actin(JF288784.1),使用Primer Premier 5.0設計引物,50.0 μL體系進行PCR擴增,將瓊脂糖凝膠有特異性條帶的PCR產物送測驗證[生工生物工程(上海)股份有限公司],得到特異性引物(表1)。完成實時熒光定量PCR檢測后,采用2-△△Ct法計算基因相對表達量。

表1 實時熒光定量PCR擴增引物序列Table 1 Real-time fluorescence quantitative PCR amplification primer sequences
1.6 TUNEL檢測
根據上海碧云天生物技術有限公司TUNEL細胞凋亡檢測試劑盒(顯色法)說明書,對凡納濱對蝦肝胰腺組織進行石蠟包埋、切片、生物素標記及樣品顯色后封片觀察。
1.7 統計分析
試驗數據采用SPSS 25.0進行單因素方差分析(One-way ANOVA)和Duncan’s多重比較。
2 結果與分析
2.1 氨氮慢性脅迫下凡納濱對蝦生長存活的變化
由表2可知,氨氮脅迫20和40 d,0.9、5.2和12.0 mg/L組凡納濱對蝦體質量增長率和存活率均顯著低于對照組(P<0.05,下同),且體質量增長率和存活率均呈隨氨氮濃度增加而降低的變化趨勢。
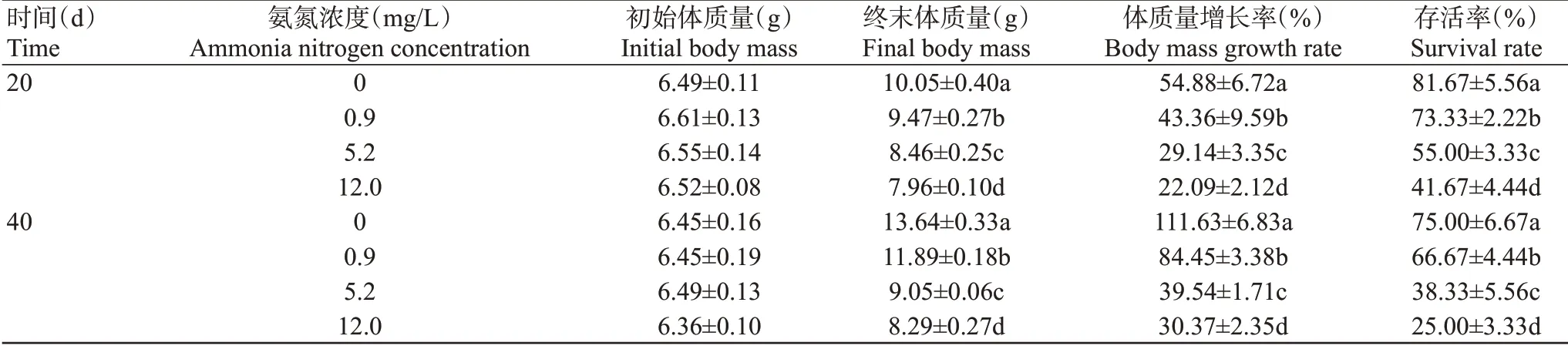
表2 氨氮慢性脅迫對凡納濱對蝦生長存活的影響Table 2 Effects of chronic ammonia nitrogen stress on growth and survival of L.vannamei
2.2 氨氮慢性脅迫下凡納濱對蝦氨代謝的變化
2.2.1 血淋巴氨氮和尿素氮含量變化.由圖1可知,氨氮脅迫20 d,各氨氮濃度組血淋巴氨氮含量分別為177.75、253.27、435.15和886.99 μmol/L,氨氮脅迫40 d,各氨氮濃度組血淋巴氨氮含量分別為145.80、317.88、649.73和1265.30 μmol/L。血淋巴氨氮和尿素氮含量均呈隨氨氮濃度增加而升高的變化趨勢,且各脅迫組顯著高于對照組。其中,在氨氮脅迫20 d,5.2 mg/L組與12.0 mg/L組的血淋巴尿素氮含量無顯著差異(P>0.05,下同),而氨氮含量差異顯著。氨氮脅迫40 d時0.9~12.0 mg/L組血淋巴氨氮含量比20 d高,氨氮脅迫20 d后血淋巴氨氮含量在體內積累,而尿素氮含量在2個時間點變化不明顯。
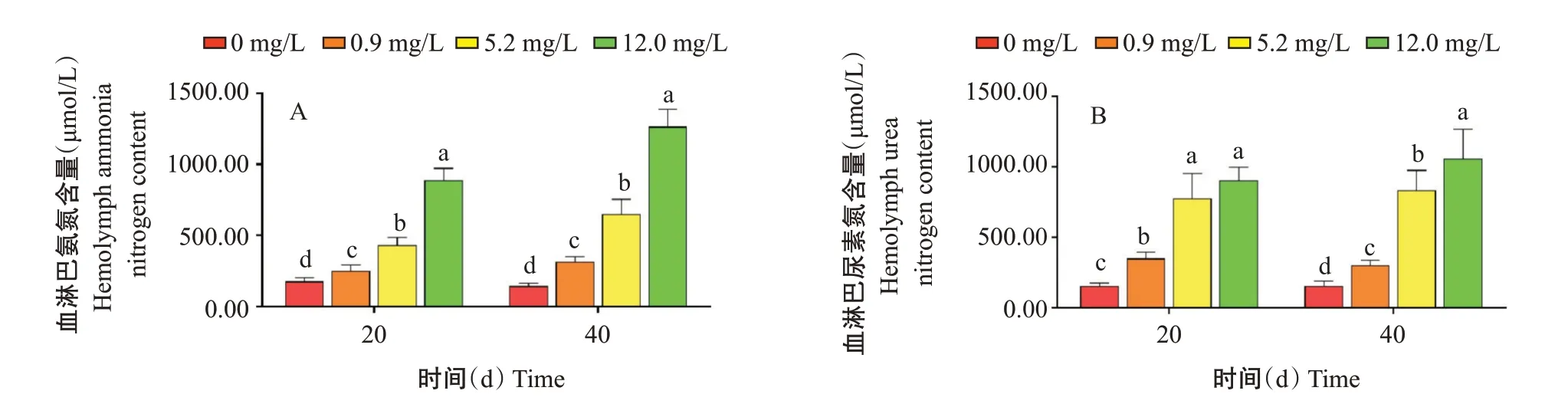
圖1 氨氮慢性脅迫對凡納濱對蝦血淋巴的影響Fig.1 Effects of chronic ammonia nitrogen stress on hemolymph of L.vannamei
2.2.2 肝胰腺GDH和GS活性變化.由圖2可知,氨氮脅迫20 d,GDH和GS活性隨氨氮濃度增加整體呈升高趨勢,0.9、5.2和12.0 mg/L組顯著高于對照組,5.2 mg/L組與12.0 mg/L組的GDH活性無顯著差異。氨氮脅迫40 d,GDH和GS活性隨氨氮濃度增加呈先升高后降低的變化趨勢;0.9和5.2 mg/L組的GDH活性顯著高于對照組,12.0 mg/L組顯著低于對照組;0.9 mg/L組的GS活性顯著高于對照組,5.2和12.0 mg/L組均顯著低于對照組。說明氨氮脅迫40 d,當氨氮濃度超過5.2 mg/L,機體合成谷氨酸和谷氨酰胺的能力受到抑制,去除氨氮的能力降低。
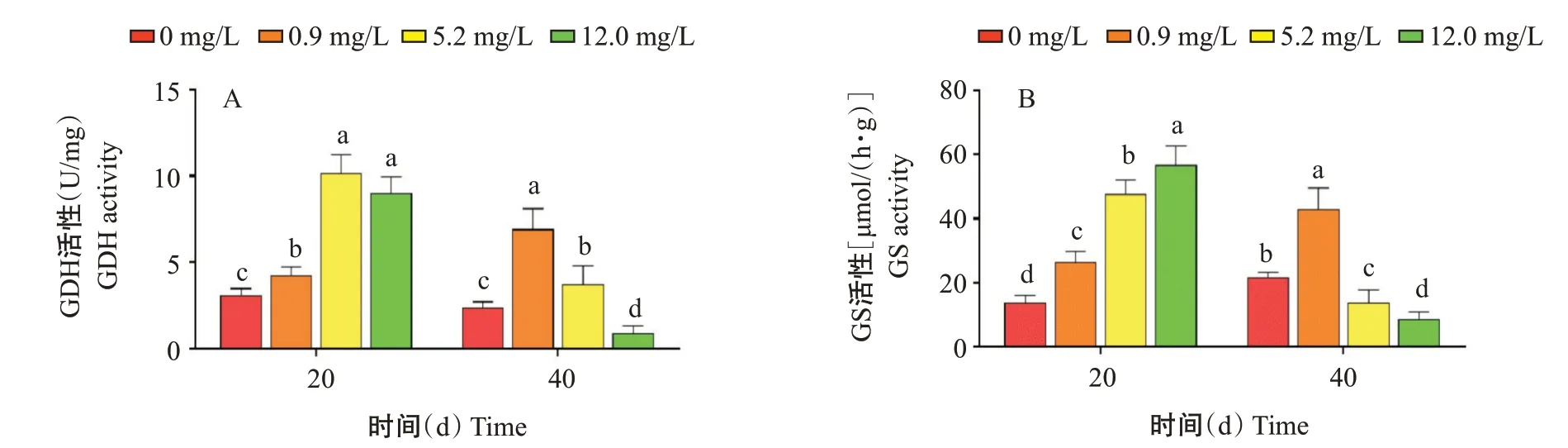
圖2 氨氮慢性脅迫對凡納濱對蝦肝胰腺GDH和GS活性的影響Fig.2 Effects of chronic ammonia nitrogen stress on GDH activity and GS activity in hepatopancreas of L.vannamei
2.3 氨氮慢性脅迫下凡納濱對蝦肝胰腺抗氧化酶活性的變化
由圖3可知,氨氮脅迫20 d,T-SOD活性隨氨氮濃度增加呈先升高后降低的變化趨勢,0.9和5.2 mg/L組顯著高于對照組,12.0 mg/L組顯著低于對照組;氨氮脅迫40 d,0.9 mg/L組顯著高于對照組,5.2和12.0 mg/L組顯著低于對照組。而氨氮脅迫20 d,CAT活性隨氨氮濃度增加整體呈升高趨勢,各脅迫組顯著高于對照組,其中5.2 mg/L組與12.0 mg/L組無顯著差異;氨氮脅迫40 d,0.9 mg/L組的CAT活性顯著高于對照組,5.2和12.0 mg/L組顯著低于對照組。說明氨氮脅迫40 d,當氨氮濃度超過5.2 mg/L,機體的抗氧化能力受到抑制。
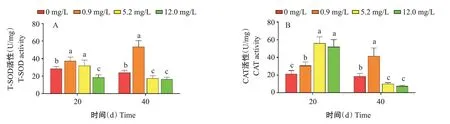
圖3 氨氮慢性脅迫對凡納濱對蝦肝胰腺抗氧化能力的影響Fig.3 Effects of chronic ammonia nitrogen stress on antioxidant activity in hepatopancreas of L.vannamei
2.4 凡納濱對蝦肝胰腺細胞凋亡基因表達量變化
由圖4可知,氨氮脅迫20 d,Cyt C基因相對表達量隨氨氮濃度增加而升高,各脅迫組顯著高于對照組,5.2 mg/L組與12.0 mg/L組無顯著差異;氨氮脅迫40 d,Cyt C基因相對表達量隨氨氮濃度增加呈先升高后降低的變化趨勢,5.2和12.0 mg/L組顯著高于對照組。氨氮脅迫20 d,Caspase-3基因相對表達量隨氨氮濃度增加而升高,0.9、5.2和12.0 mg/L組顯著高于對照組;氨氮脅迫40 d,Caspase-3基因相對表達量隨氨氮濃度增加整體呈升高趨勢,各脅迫組顯著高于對照組,5.2 mg/L組與12.0 mg/L組無顯著差異。表明受到氨氮脅迫后,各濃度組機體發生不同程度的細胞凋亡。
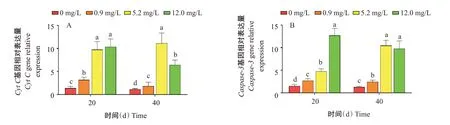
圖4 氨氮慢性脅迫對凡納濱對蝦肝胰腺Cyt C和Caspase-3基因相對表達量的影響Fig.4 Effects of chronic ammonia nitrogen stress on relative expression level of Cyt C gene and Caspase-3 gene in hepatopancreas of L.vannamei
2.5 凡納濱對蝦肝胰腺TUNEL檢測結果
由圖5可知,氨氮脅迫20和40 d,對照組凋亡細胞數量極少,肝小管飽滿,排列整齊,邊界清晰,星狀管腔形狀完整。隨氨氮濃度增加,凋亡細胞數量增加,肝小管變形,排列紊亂,邊界模糊,星狀管腔變形,氨氮脅迫40 d較20 d細胞凋亡情況更嚴重。
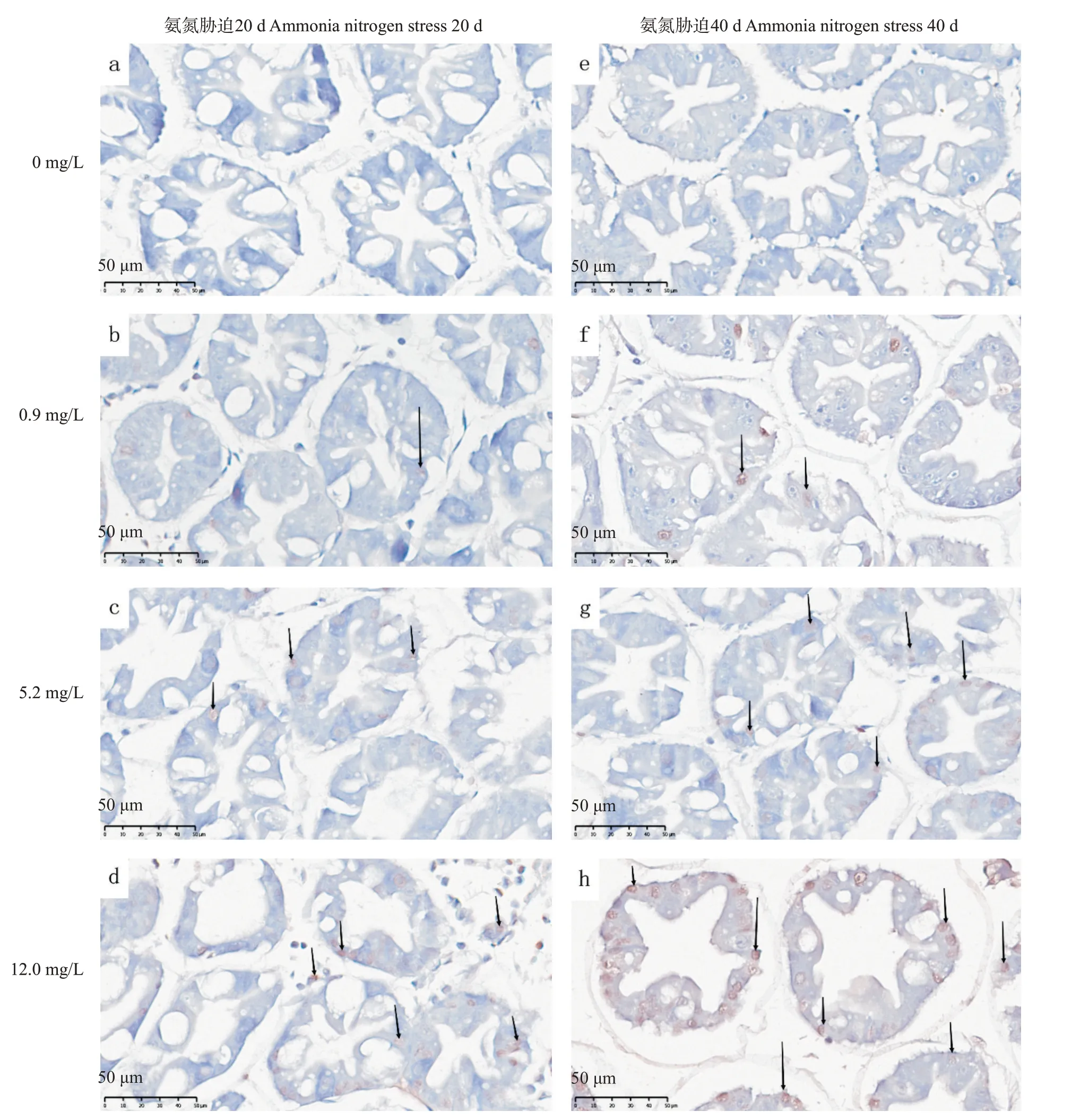
圖5 氨氮慢性脅迫下凡納濱對蝦肝胰腺TUNEL檢測結果Fig.5 TUNEL results of L.vannamei hepatopancreas under chronic ammonia nitrogen stress
3 討論
經過氨氮脅迫20和40 d后,凡納濱對蝦體質量增長率和存活率隨氨氮濃度增加而降低,隨時間和氨氮濃度的增加,凡納濱對蝦體質量增長越慢,存活率越低。通過TUNEL檢測結果顯示,肝胰腺凋亡細胞數量隨氨氮濃度增加而增加,且氨氮脅迫40 d凋亡細胞數量較20 d更多。李志輝等(2018)研究表明,脊尾白蝦(Exopalaemon carinicauda)養殖密度越高,養殖水體中的氨氮含量越高,特定生長率和存活率受到氨氮脅迫的抑制。王夢杰等(2021)研究表明,氨氮脅迫濃度越高,臺灣泥鰍(Paramisgurnusdabryanusssp.Taiwan)幼魚的增重率和特定生長率越低,且處理組的肝組織細胞核發生偏移、溶解,細胞邊界模糊。由此可知,氨氮慢性脅迫后,對凡納濱對蝦生長和存活產生不利影響,與肝胰腺受損密切相關。
在水體中,氨氮以NH4+和NH3形式存在,NH3可直接進入細胞膜,破壞膜結構,影響蛋白質代謝和損傷DNA(Martins Dutra et al.,2016)。為應對氨氮脅迫,甲殼類依賴氨代謝途徑將氨氮主動排出體外或代謝為尿素和谷氨酰胺等低毒、中性物質(Ip et al.,2004;周發林等,2016)。Hong等(2007)研究指出,中華絨螯蟹(Eriocheir sinensis)經48 h氨氮脅迫后,血淋巴氨氮含量顯著升高。王蕓(2011)研究發現,中國對蝦(Fenneropenaeus chinensis)經96 h氨氮脅迫,2、4、6和8 mg/L組血淋巴氨氮和尿素氮含量呈上升趨勢,為減少氨氮累積,凡納濱對蝦機體開啟氨代謝解毒機制,加速尿素氮生成。本研究結果中,凡納濱對蝦經氨氮脅迫20 d后,各濃度組血淋巴氨氮含量分別為177.75、253.27、435.15和886.99 μmol/L;氨氮脅迫40 d,各濃度組血淋巴氨氮含量分別為145.80、317.88、649.73和1265.30 μmol/L。血淋巴尿素氮含量呈隨氨氮濃度增加而升高的趨勢,各濃度組血淋巴氨氮含量均高于各組環境的氨氮濃度,且氨氮脅迫40 d血淋巴氨氮含量較20 d多,說明環境氨氮會擴散并積累在血淋巴中。機體隨氨氮脅迫時間越長,對血淋巴氨氮清除能力會降低,而尿素氮含量在20和40 d水平相差不明顯,反映出機體對氨氮轉化為尿素氮水平在20 d達最高水平。血淋巴連接各個組織,氨氮通過血淋巴影響肝胰腺組織代謝。為代謝多余的氨氮,肝胰腺GDH和GS聯合發揮作用,將氨和谷氨酸轉化為谷氨酰胺,從而達到降低氨氮毒性保護機體代謝的作用(李志輝等,2018)。何玉英等(2016)使用16 mg/L氨氮脅迫中國對蝦72 h,肝胰腺GDH基因相對表達量隨時間增加而升高,氨代謝途徑被誘導發生,加速合成谷氨酰胺以保護機體。邱立國(2017)研究表明,氨氮濃度在3.4~24.6 mg/L,凡納濱對蝦肌肉GDH和GS活性在5~10 d時受到抑制,過高濃度的氨氮超過肌肉代謝范圍,導致代謝紊亂。而本研究中,氨氮脅迫20 d后,GDH和GS活性顯著高于對照組,氨氮脅迫40 d后,氨氮濃度高于5.2 mg/L時,谷氨酸代謝途徑受到抑制,說明長期受氨氮脅迫,機體受到損傷,導致氨代謝途徑紊亂,谷氨酰胺合成能力受阻,造成血淋巴氨氮含量大量累積。
據報道,氨氮脅迫會刺激肝胰腺產生過多的活性氧,對抗氧化系統造成影響,為應對氧化應激產生的活性氧損傷,機體可通過抗氧化酶來平衡氧化還原反應,從而起到保護機體的功能正常運轉和適應脅迫環境的作用(何玉英等,2016)。T-SOD和CAT聯合作用清除活性氧和過氧化氫的抗氧化作用是甲殼類先天性免疫中至關重要的一部分(于杰倫,2019)。王蕓(2011)對中國對蝦進行96 h氨氮脅迫,肝胰腺CAT活性在0~48 h受氨氮刺激顯著升高,在48~96 h活性氧含量過高,CAT活性受到抑制。熊大林等(2020)用15 mg/L濃度氨氮脅迫凡納濱對蝦72 h,脅迫組鰓T-SOD活性顯著升高,活性氧含量顯著降低。較低濃度的氨氮可刺激抗氧化酶活性上升,而高濃度氨氮會抑制抗氧化酶活(Sun et al.,2014)。本研究結果中,氨氮脅迫20 d,0.9和5.2 mg/L組T-SOD活性顯著高于對照組,12.0 mg/L組顯著低于對照組;而氨氮脅迫40 d,5.2和12.0 mg/L組T-SOD活性受到抑制;這可能是高濃度或長期氨氮脅迫導致血淋巴氨氮大量積累,對機體造成嚴重損傷,抑制T-SOD活性。在氨氮脅迫20 d,CAT活性隨氨氮濃度的增加而升高;而在氨氮脅迫40 d,5.2和12.0 mg/L組的CAT活性顯著低于對照組。說明氨氮濃度高于5.2 mg/L脅迫40 d后,機體內大量活性氧難以被有效清除,對機體造成氧化損傷。
機體抗氧化能力不足以對抗氧化應激時,過多的活性氧會增加細胞凋亡率(邱立國,2017)。CytC能激活Caspase-3,而Caspase-3是線粒體凋亡途徑和死亡受體途徑中的常見效應子,Caspase-3的激活象征著細胞進入不可逆的凋亡階段(Jung et al.,2014)。李玲等(2021)用10.5 mg/L鎘(Cd)刺激凡納濱對蝦,24 h后凡納濱對蝦體內產生大量的活性氧,Caspase-3基因表達量顯著上升。賈旭穎(2014)研究發現凡納濱對蝦受高濃度非離子氨脅迫后,抗氧化酶活性被抑制,產生過量活性氧誘導肝胰腺CytC含量和Caspase-3活性升高。在本研究中,凡納濱對蝦受氨氮脅迫20和40 d后,肝胰腺CytC和Caspase3基因相對表達量均隨氨氮濃度增加呈升高趨勢。氨氮脅迫會使活性氧在肝胰腺內大量產生,增加CytC的表達量,大量的細胞色素C釋放,從而調節Caspase-3表達量升高,誘導細胞凋亡發生,對組織造成損傷,導致凡納濱對蝦氨代謝紊亂,機體對氨氮清除能力下降。
4 結論
經慢性氨氮脅迫后,隨氨氮濃度增加和脅迫時間延長,凡納濱對蝦血淋巴氨氮累積越多,尿素氮含量隨之增加,肝胰腺谷氨酰胺合成途徑受到抑制,氨代謝受到阻礙,應激產生的活性氧超出抗氧化系統調節范圍,對肝胰腺造成損傷,誘導細胞凋亡發生,對凡納濱對蝦生長存活造成不利影響。
