基于eDNA宏條形碼技術(shù)的喀斯特高原人工湖泊魚類多樣性分析
2022-02-03 07:57:40許龍飛姚鄧雕楊原偉郭星辰李君軼姜海波安苗董響紅邵儉
南方農(nóng)業(yè)學(xué)報(bào) 2022年10期
許龍飛,姚鄧雕,楊原偉,郭星辰,李君軼,姜海波,安苗,董響紅,邵儉*
(1貴州大學(xué)動(dòng)物科學(xué)學(xué)院/高原山地動(dòng)物遺傳育種與繁殖教育部重點(diǎn)實(shí)驗(yàn)室,貴州貴陽 550025;2中國水產(chǎn)科學(xué)研究院長江水產(chǎn)研究所/農(nóng)業(yè)農(nóng)村部淡水生物多樣性保護(hù)重點(diǎn)實(shí)驗(yàn)室,湖北武漢 430223)
0 引言
【研究意義】我國以云貴高原為中心的西南地區(qū)喀斯特地貌豐富,是世界三大喀斯特集中分布區(qū)域之一,其中貴州境內(nèi)61.9%的國土面積為喀斯特地貌(李秋華等,2013;阮玉龍等,2013)。西南喀斯特地區(qū)是我國珍稀物種、瀕危物種、特有物種分布最集中、種類最豐富的地區(qū)之一(王波等,2018);但喀斯特生態(tài)系統(tǒng)也是典型的脆弱生態(tài)系統(tǒng),土層淺薄,土壤承載力差,生態(tài)地質(zhì)環(huán)境脆弱(賀忠權(quán)等,2021)。近年來,人工筑壩導(dǎo)致的河道破碎、工農(nóng)業(yè)污染引起的水體富營養(yǎng)化及不合理引種造成的外來物種生物入侵等,致使我國喀斯特地區(qū)土著魚類種群受到不同程度的脅迫。因此,亟待摸清喀斯特高原魚類群落現(xiàn)狀,為保護(hù)西南喀斯特地區(qū)魚類多樣性提供科學(xué)依據(jù)。【前人研究進(jìn)展】紅楓湖和阿哈湖是典型的喀斯特高原人工湖泊。紅楓湖位于貴州省清鎮(zhèn)市,是貓?zhí)犹菁?jí)水電站攔河而成的人工湖泊,屬長江流域?yàn)踅担ㄔA獻(xiàn)等,2020);阿哈湖位于貴州省貴陽市西南郊8 km,流域面積180.3 km2,總庫容0.87億m3,同屬長江流域?yàn)踅担S軼婧等,2020)。至今,有關(guān)紅楓湖和阿哈湖的研究多圍繞水質(zhì)評(píng)價(jià)(郭云等,2015;韓翠紅等,2020;金祖雪等,2020;費(fèi)志軍等,2021),涉及魚類資源調(diào)查的研究較少,對(duì)于二者的魚類種群現(xiàn)狀尚不清楚。紅楓湖和阿哈湖作為長江上游重要的生態(tài)屏障,研究其魚類多樣性是維持當(dāng)?shù)厮鷳B(tài)系統(tǒng)的關(guān)鍵,對(duì)于維持和補(bǔ)充長江魚類資源同樣具有重要意義(de Paula Gutiérrez and Agudelo,2020;Wang et al.,2021)。環(huán)境DNA(Environmental DNA,eDNA)是指生物散落在環(huán)境中的DNA片段,通常來源于生物的皮膚碎屑、配子、黏液及糞便等(Bohmann et al.,2014)。Ogram等(1987)利用eDNA研究土壤沉積物中的微生物,并首次提出eDNA的概念。eDNA宏條形碼技術(shù)是利用環(huán)境中的DNA結(jié)合高通量測序技術(shù)完成對(duì)環(huán)境樣本總DNA測序,通過生物信息學(xué)分析,以及與公開物種序列數(shù)據(jù)庫進(jìn)行比對(duì),最終得到環(huán)境樣本中物種組成的技術(shù)(Shu et al.,2020;楊海樂等,2021)。目前,eDNA宏條形碼技術(shù)已廣泛應(yīng)用于監(jiān)測魚類空間結(jié)構(gòu)(Zhang et al.,2020)、魚類物種組成與種群豐度(王夢等,2022)及魚類生物入侵現(xiàn)狀(Zhang et al.,2022)等領(lǐng)域。eDNA宏條形碼技術(shù)的優(yōu)點(diǎn)包括:(1)不受科研人員的主觀判斷影響;(2)避免了長期的魚類樣本采集,極大提高了物種監(jiān)測效率;(3)eDNA宏條形碼技術(shù)不入侵環(huán)境,對(duì)監(jiān)測物種及其棲息地幾乎不產(chǎn)生影響(Antognazza et al.,2019;Aglieri et al.,2021)。【本研究切入點(diǎn)】隨著國際動(dòng)物保護(hù)公約、動(dòng)物福利及生態(tài)生物多樣性保護(hù)等法規(guī)措施的執(zhí)行,eDNA宏條形碼技術(shù)無疑更符合當(dāng)下生物多樣性監(jiān)測的需求。【擬解決的關(guān)鍵問題】利用eDNA宏條形碼技術(shù)探究喀斯特高原人工湖泊魚類多樣性,綜合評(píng)估紅楓湖和阿哈湖魚類群落結(jié)構(gòu)及環(huán)境因子對(duì)喀斯特高原人工湖泊魚類分布的影響,以期為喀斯特地區(qū)水域魚類多樣性評(píng)估提供新方法、新思路,也為紅楓湖和阿哈湖魚類保護(hù)管理措施的制定積累基礎(chǔ)資料。
1 材料與方法
1.1 樣點(diǎn)設(shè)置及樣品采集
紅楓湖和阿哈湖樣品采集時(shí)間為2021年11月25日,采樣點(diǎn)設(shè)置參考SC/T 9402—2010《淡水浮游生物調(diào)查技術(shù)規(guī)范》,具體如下:紅楓湖分為南湖和北湖,在南、北湖湖心處、湖泊入水口、出水口設(shè)置采樣點(diǎn),結(jié)合紅楓湖水域特點(diǎn),環(huán)湖布置采樣點(diǎn),南、北湖各設(shè)5個(gè)采樣點(diǎn),共計(jì)10個(gè)采樣點(diǎn);在阿哈湖湖心處、湖泊入水口、出水口設(shè)置采樣點(diǎn),結(jié)合阿哈湖水域特點(diǎn),環(huán)湖布置采樣點(diǎn),共計(jì)7個(gè)采樣點(diǎn)(圖1)。樣品采集全部在船上完成,使用采水器分水體上(1 m)、中(5 m)、下層(10 m)采樣混合后,每個(gè)采樣點(diǎn)留取2 L水樣低溫避光保存,送回實(shí)驗(yàn)室立即抽濾,實(shí)驗(yàn)人員操作時(shí)每個(gè)采樣點(diǎn)更換1次手套。采樣過程記錄采樣點(diǎn)的經(jīng)緯度,使用黑白盤測定水體透明度(Transparency,Tra),并以YSI便攜式水質(zhì)儀測定水溫(Water temperature,WT)、總?cè)芙夤腆w(Total dissolved solids,TDS)、鹽度(Salinity,Sal)、pH、溶解氧(Dissolved oxygen,DO)、離子氨(Ionic ammonia,NH4+)等環(huán)境因子(表1)。
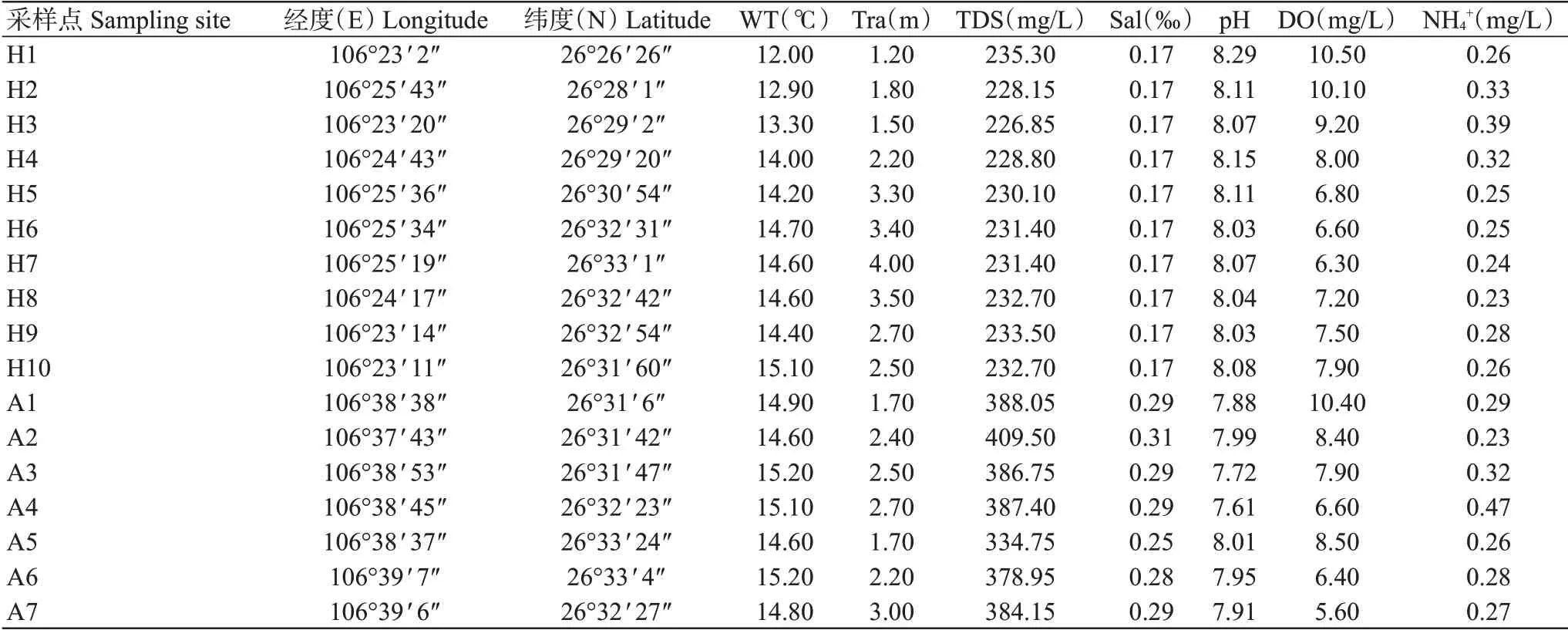
表1 eDNA樣品采集點(diǎn)地理位置及其環(huán)境因子Table 1 Geographic locations and environmental factors of eDNA sampling sites
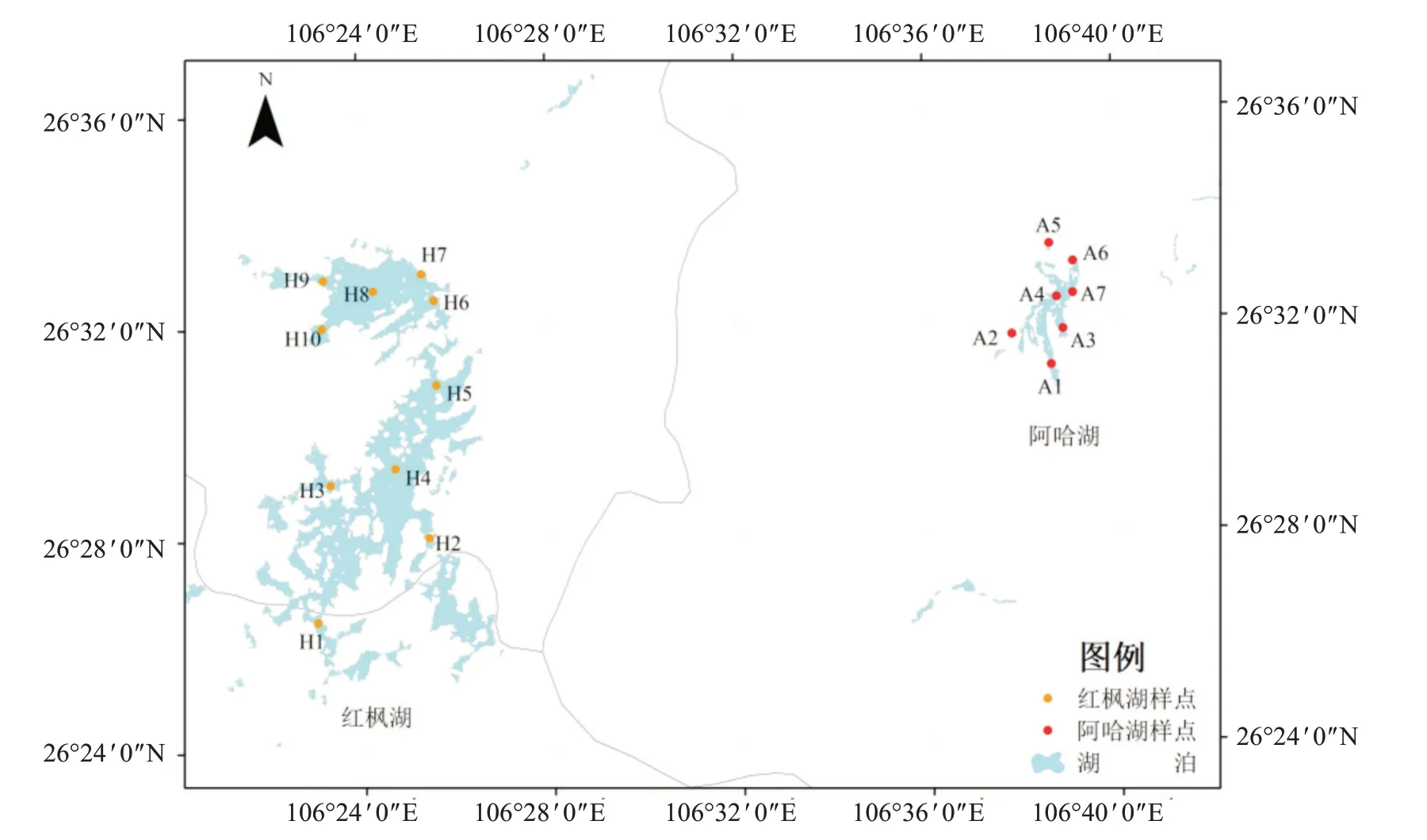
圖1 紅楓湖和阿哈湖采樣點(diǎn)設(shè)置分布情況Fig.1 Sampling site distribution of Hongfeng Lake and Aha Lake
1.2 eDNA提取及擴(kuò)增
使用隔膜真空泵和砂芯玻璃過濾裝置抽濾水樣,選用0.45 μm孔徑的玻璃纖維濾膜。抽濾時(shí)操作環(huán)境充分通風(fēng),每過濾1份樣品均用1%次氯酸溶液處理實(shí)驗(yàn)操作臺(tái)面及過濾裝置,實(shí)驗(yàn)人員更換1次手套。設(shè)1個(gè)空白對(duì)照,使用同樣濾膜抽濾2 L的ddH2O檢測有無環(huán)境污染。抽濾完成后將濾膜置于1.5 mL離心管中,-20℃保存?zhèn)溆谩J褂肙MEGA公司的Water DNA Kit D5525試劑盒提取水體eDNA,選用Tele 02硬骨魚類通用引物(Tele 02-F:5'-AAA CTCGTGCCAGCCACC-3';Tele 02-R:5'-GGGTATC TAATCCCAGT TTG-3')進(jìn)行PCR擴(kuò)增(Taberlet et al.,2018)。對(duì)提取的eDNA進(jìn)行凝膠電泳檢測,質(zhì)檢合格的樣本上機(jī)測序,測序平臺(tái)為Illumina NovaSeq 6000。
1.3 測序及生物信息學(xué)分析
設(shè)10 bp的窗口,若窗口內(nèi)平均質(zhì)量值低于20,則從窗口開始截去后端堿基,過濾質(zhì)控后50 bp以下的序列(Trimmomatic v0.3),根據(jù)雙端測序的overlap關(guān)系,將成對(duì)序列拼接成1條序列,最小overlap長度為10 bp,overlap區(qū)允許的最大錯(cuò)配比率為0.2,篩除不符合序列(FLASH v1.2.7),按照標(biāo)簽和引物序列進(jìn)行拆分并調(diào)整序列方向,得到每個(gè)樣本的優(yōu)質(zhì)序列。對(duì)優(yōu)質(zhì)序列按照98%相似性進(jìn)行OTU聚類(Zhang et al.,2020),在聚類過程中去除嵌合體,生成OTUs表格(perl v5.18.2,Usearch v10,qiime v1.9.1)。將生成的OTUs表格在基于NCBI網(wǎng)站nt數(shù)據(jù)庫自建的淡水魚類數(shù)據(jù)庫中進(jìn)行物種基因注釋(建庫時(shí)間2021年10月),得到魚類物種信息,魚類分類依據(jù)參考Fishbase數(shù)據(jù)庫。
1.4 魚類多樣性數(shù)據(jù)處理分析
為確保鑒定結(jié)果更準(zhǔn)確,對(duì)鑒定結(jié)果進(jìn)行如下校正:(1)將低豐度序列(序列樣本平均讀數(shù)低于1)訂正為未識(shí)別;(2)將鑒定到的雜交種進(jìn)行校正,若雜交種父母本隸屬于同屬魚類A屬,則將該雜交種訂正為Asp.,否則將該雜交種序列訂正為未識(shí)別。使用Excle 2020分析含魚類物種信息注釋的OTUs表格,并繪制物種組成餅圖;基于魚類代表OTUs序列讀數(shù),繪制屬分類水平魚類分布熱圖,為消除數(shù)據(jù)差異過大對(duì)熱圖的影響,先將序列讀數(shù)進(jìn)行均一化處理,再使用R語言繪制熱圖。同時(shí),使用R語言進(jìn)行PerMANOVA分析,明確紅楓湖南、北湖魚類群落結(jié)構(gòu)差異性;利用R語言vegan包進(jìn)行冗余分析(Redundancy analysis,RDA),將魚類代表OTUs序列讀數(shù)進(jìn)行Hellinger轉(zhuǎn)化,環(huán)境因子數(shù)據(jù)進(jìn)行均一化以消除量綱影響,RDA分析前需對(duì)環(huán)境因子進(jìn)行共線性分析,剔除方差膨脹因子過大的環(huán)境因子。
2 結(jié)果與分析
2.1 PCR擴(kuò)增產(chǎn)物質(zhì)量檢測結(jié)果
紅楓湖10個(gè)eDNA樣本和阿哈湖7個(gè)eDNA樣本的PCR擴(kuò)增產(chǎn)物質(zhì)量檢測目的條帶大小正確,均滿足測序條件,而空白對(duì)照組無任何條帶,說明試驗(yàn)過程中無污染。
2.2 高通量測序結(jié)果
紅楓湖10個(gè)eDNA樣本的物種累積曲線如圖2-A所示,隨著測序深度的增加,觀測到的OTUs數(shù)量趨于穩(wěn)定,說明eDNA樣本數(shù)量足以代表紅楓湖魚類多樣性。阿哈湖7個(gè)eDNA樣本的物種累積曲線如圖2-B所示,隨著樣本數(shù)量的增加,觀測到的OTUs數(shù)量趨于穩(wěn)定,也說明eDNA樣本數(shù)量足以反映阿哈湖魚類多樣性。
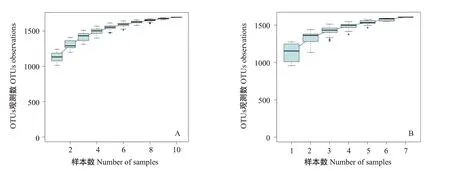
圖2 紅楓湖(A)和阿哈湖(B)的eDNA樣本物種累積曲線Fig.2 Cumulative curve of species of eDNA samples from Hongfeng Lake(A)and Aha Lake(B)
2.3 魚類基因注釋結(jié)果
經(jīng)人工校正后,在紅楓湖鑒定出魚類32種,隸屬于5目11科27屬,有2種 魚 類 鑒 定 到 屬[鱖 屬(Siniperca)],有11種魚類在歷史數(shù)據(jù)庫中得到匹配(表2)。其中,鯉形目(Cypriniformes)魚類18種,鲇形目(Siluriformes)魚類5種,日鱸目(Centrarchiformes)魚類5種,胡瓜魚目(Osmeriformes)魚類2種,鰕虎魚目(Gobiiformes)魚類2種(圖3-A)。在阿哈湖鑒定到魚類33種,隸屬于6目12科28屬,有3種魚類鑒定到屬(表2)。其中,鯉形目魚類18種,鲇形目魚類5種,日鱸目魚類5種,胡瓜魚目魚類2種,鰕虎魚目魚類2種,攀鱸目(Anabantiformes)魚類1種(圖3-B)。

圖3 紅楓湖(A)和阿哈湖(B)的魚類物種組成結(jié)構(gòu)Fig.3 Fish species composition of Hongfeng Lake(A)and Aha Lake(B)

表2 基于eDNA宏條形碼技術(shù)在紅楓湖和阿哈湖各采樣點(diǎn)的魚類檢出情況Table 2 Detection of fish from each sampling site in Hongfeng Lake and Aha Lake based on eDNA metabarcoding technology
在屬分類水平上,紅楓湖魚類在各采樣點(diǎn)的分布情況如圖4顯示。紅楓湖魚類代表OTUs(屬分類水平)序列讀數(shù)排名前10的分別是鱯屬(Hemibagrus)、鯉屬(Cyprinus)、鱖屬、黃顙魚屬(Tachysurus)、鰱屬(Hypophthalmichthys)、屬(Hemiculter)、新銀魚屬(Neosalanx)、草魚屬(Ctenopharyngodon)、青魚屬(Mylopharyngodon)和特鯰屬(Tatia)。阿哈湖屬分類水平魚類在各采樣點(diǎn)的分布情況如圖5所示,阿哈湖魚類代表OTUs(屬分類水平)序列讀數(shù)排名前10的分別是鰱屬、鱯屬、鱖屬、鯉屬、黃顙魚屬、屬、新銀魚屬、草魚屬、吻鰕虎魚屬(Rhinogobius)和鳑鲏屬(Rhodeus)。
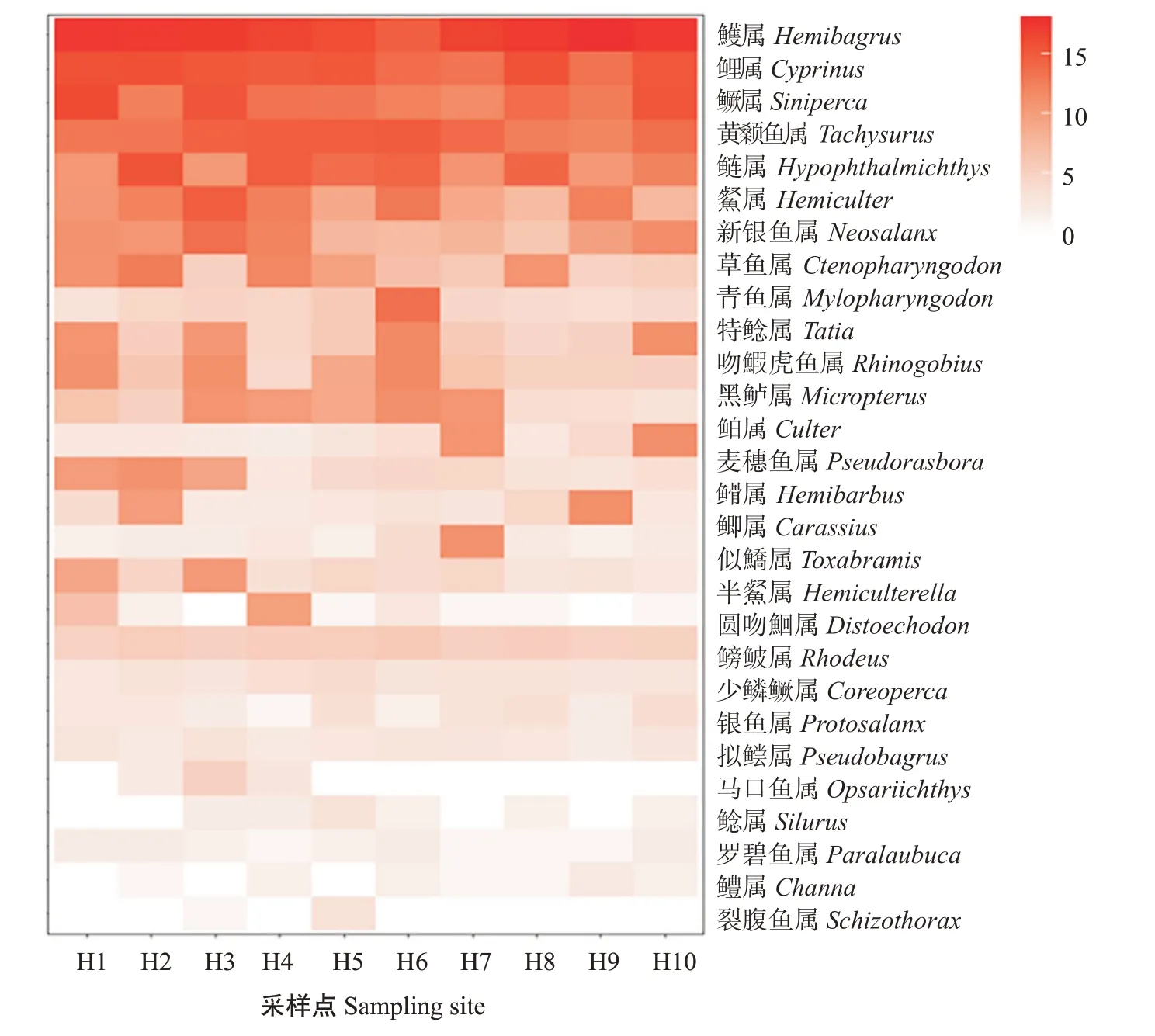
圖4 基于紅楓湖各采樣點(diǎn)魚類代表OTUs(屬分類水平)序列讀數(shù)的魚類分布熱圖Fig.4 Heat map of the distribution of fish OTUs(genus classification level)sequence reads based on sampling sites in Hongfeng Lake

圖5 基于阿哈湖各采樣點(diǎn)魚類代表OTUs(屬分類水平)序列讀數(shù)的魚類分布熱圖Fig.5 Heat map of the distribution of fish OTUs(genus classification level)sequence reads based on sampling sites in Aha Lake
2.4 紅楓湖和阿哈湖魚類多樣性
紅楓湖各采樣點(diǎn)魚類的Chao1指數(shù)顯示,在H6采樣點(diǎn)觀察到的OTUs數(shù)量最低(1205.08),在H10采樣點(diǎn)觀察到的OTUs數(shù)量最高(1434.26);在Shannon指數(shù)方面,H7采樣點(diǎn)群落多樣性最高,H9采樣點(diǎn)群落多樣性最低;Simpson指數(shù)的變化趨勢與Shannon指數(shù)一致(表3)。在監(jiān)測到的32種魚類中,有27種魚類在所有采樣點(diǎn)均能監(jiān)測到,紅楓湖全部采樣點(diǎn)均無特有魚類,各采樣點(diǎn)檢出魚類總數(shù)差異不明顯,保持在28~30種。紅楓湖南湖監(jiān)測到32種魚類,其中2種為南湖獨(dú)有魚類;北湖監(jiān)測到30種魚類,無北湖獨(dú)有魚類(表2)。PerMANOVA分析結(jié)果顯示,紅楓湖南、北湖魚類群落結(jié)構(gòu)差異不顯著(R2=0.08,P=0.562>0.05)。

表3 紅楓湖魚類α多樣性指數(shù)Table 3 α diversity index of fish in Hongfeng Lake
阿哈湖各采樣點(diǎn)魚類的Chao1指數(shù)顯示,在A7采樣點(diǎn)觀察到的OTUs數(shù)量最低(1051.95),在A5采樣點(diǎn)觀察到的OTUs數(shù)量最高(1435.24);在Shannon指數(shù)方面,A5采樣點(diǎn)群落多樣性最高,A3采樣點(diǎn)群落多樣性最低;Simpson指數(shù)則顯示以A6采樣點(diǎn)群落多樣性最高、A3采樣點(diǎn)群落多樣性最低(表4)。在監(jiān)測到的33種魚類,有26種魚類在所有采樣點(diǎn)中均能監(jiān)測到,阿哈湖全部采樣點(diǎn)均無獨(dú)有魚類,各采樣點(diǎn)檢出魚類總數(shù)差異不明顯,保持在28~32種(表2)。

表4 阿哈湖魚類α多樣性指數(shù)Table 4 α diversity index of fish in Aha Lake
2.5 魚類多樣性與環(huán)境因子的相關(guān)性
對(duì)紅楓湖魚類多樣性與環(huán)境因子進(jìn)行RDA分析前,在共線性分析中剔除了Tra和Sal 2個(gè)環(huán)境因子,剩余5個(gè)環(huán)境因子(表5)對(duì)引起紅楓湖各采樣點(diǎn)魚類群落結(jié)構(gòu)差異的解釋度為69.38%。RDA分析結(jié)果(圖6-A)顯示,紅楓湖各采樣點(diǎn)分布較散,無聚集趨勢。在RDA1軸上,WT呈正相關(guān),其余環(huán)境因子呈負(fù)相關(guān);在RDA2軸上,pH、DO和NH4+呈正相關(guān),其余環(huán)境因子呈負(fù)相關(guān)。對(duì)阿哈湖魚類多樣性與環(huán)境因子進(jìn)行RDA分析前,在共線性分析中剔除了Sal和pH 2個(gè)環(huán)境因子,剩余5個(gè)環(huán)境因子(表5)對(duì)引起阿哈湖各采樣點(diǎn)魚類群落結(jié)構(gòu)差異的解釋度為68.74%。RDA分析結(jié)果(圖6-B)顯示,阿哈湖各采樣點(diǎn)分布較散,也無聚集趨勢,其中A3、A5采樣點(diǎn)與其余采樣點(diǎn)距離較遠(yuǎn),說明這2個(gè)采樣點(diǎn)與其余采樣點(diǎn)魚類群落相似度更低。在RDA1軸上,NH4+、DO和WT呈正相關(guān),其余環(huán)境因子呈負(fù)相關(guān);在RDA2軸上,5個(gè)環(huán)境因子均呈正相關(guān)。
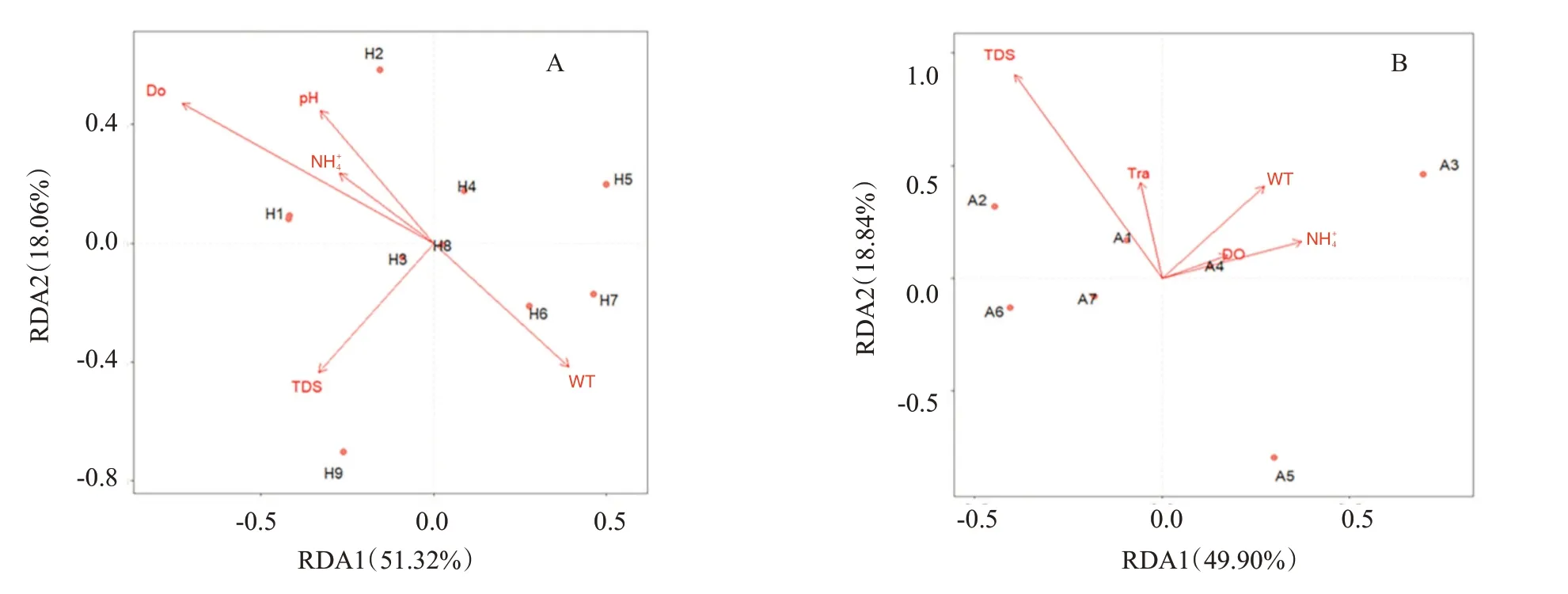
圖6 紅楓湖(A)和阿哈湖(B)魚類多樣性與環(huán)境因子的RDA分析結(jié)果Fig.6 RDA results of fish diversity and environmental factors of Hongfeng Lake(A)and Aha Lake(B)

表5 RDA分析中各環(huán)境因子的決定系數(shù)及其相關(guān)性Table 5 Determination coefficients and correlation of each environmental factor in RDA
3 討論
3.1 紅楓湖和阿哈湖魚類組成
本研究在紅楓湖設(shè)10個(gè)采樣點(diǎn),通過eDNA宏條形碼技術(shù)共監(jiān)測到32種魚類,其中有11種魚類在歷史數(shù)據(jù)庫(牟洪民等,2012)中得到驗(yàn)證。紅楓湖有21種魚類沒有在歷史數(shù)據(jù)庫中得到驗(yàn)證,主要是由于歷史數(shù)據(jù)庫為2010—2011年采集樣本,對(duì)于目前紅楓湖現(xiàn)有魚類群落組成尚不清楚,說明選取的歷史數(shù)據(jù)庫不能完全代表紅楓湖現(xiàn)階段魚類群落結(jié)構(gòu)。此外,利用公用魚類線粒體12S序列文庫進(jìn)行物種信息注釋可能會(huì)對(duì)個(gè)別親緣近的物種出現(xiàn)偏差(Lamy et al.,2021;de Santana et al.,2021)。本研究監(jiān)測到鰱(H.molitrix)、鳙(H.nobilis)(H.leucisculus)、南方馬口魚(O.uncirostris)、棒花魚(A.rivularis)、鯉(C.carpio)、草魚(C.idella)等紅楓湖歷史優(yōu)勢魚類,但1982和2010年2次紅楓湖魚類資源調(diào)查中監(jiān)測到的紅鰭鲌(Culter erythropterus)、長春鳊(Parabramis pekinensis)、三 角 魴(Magalobrama tarminalis)、泥鰍(Misgurnus anguillicaudatus)、黃鱔(Monopterus albus)等魚類(牟洪民等,2012)在本研究中均未監(jiān)測到,說明這些魚類種群極度萎縮或已消失。本研究在紅楓湖監(jiān)測到鯉形目魚類18種(占56.25%),還包括日鱸目大口黑鱸(M.salmoides)、中國少鱗鱖(C.whiteheadi)、鱖(S.chuatsi)及鱖屬(Sinipercasp.)未定種魚類2種,胡瓜魚目太湖新銀魚(N.taihuensis)、中國大銀魚(P.chinensis),鲇形目斑鱯(H.guttatus)、南方鯰(S.meridionalis)等常見非紅楓湖土著經(jīng)濟(jì)養(yǎng)殖魚類。阿哈湖并無傳統(tǒng)的魚類資源調(diào)查數(shù)據(jù)作為歷史數(shù)據(jù)庫,本研究在阿哈湖監(jiān)測到鯉形目魚類18種(占54.55%),另有鱧屬未定種魚類1種,日鱸目大口黑鱸、中國少鱗鱖、鱖及鱖屬未定種魚類2種,胡瓜魚目太湖新銀魚、中國大銀魚,鲇形目斑鱯、南方鯰等常見經(jīng)濟(jì)養(yǎng)殖魚類。可見,現(xiàn)階段的紅楓湖和阿哈湖魚類組成以鯉形目魚類為主,但存在以日鱸目為主的養(yǎng)殖魚類生態(tài)入侵。
3.2 高通量測序序列讀數(shù)與魚類種群豐度的關(guān)系
本研究結(jié)果顯示,在OTUs(屬分類水平)序列讀數(shù)排名中,紅楓湖魚類以鱯屬排名第一、特鯰屬排名第十,阿哈湖魚類以鱯屬排名第一、黃顙魚屬排名第五、特鯰屬排名第十三。鱯屬僅監(jiān)測到斑鱯1種魚類,斑鱯隸屬于鲇形目鲿科(Bagridae),俗稱芝麻劍,是我國傳統(tǒng)的名貴養(yǎng)殖魚類,主要分布在錢塘江、九龍江和珠江等水系(劉偉,2016),而紅楓湖和阿哈湖屬于長江水系。在紅楓湖監(jiān)測到斑鱯eDNA信號(hào)可能是由于2021年更新的《國家重點(diǎn)保護(hù)野生動(dòng)物名錄》將斑鱯野生種群列為國家二級(jí)保護(hù)動(dòng)物(養(yǎng)殖群體除外),貴州多地開展斑鱯人工繁育及養(yǎng)殖所造成,但斑鱯是否成為紅楓湖和阿哈湖魚類的優(yōu)勢種尚有待進(jìn)一步考證。黃顙魚隸屬于鲇形目鲿科,在我國長江、黃河及珠江等各大水系均有分布,是常見的經(jīng)濟(jì)養(yǎng)殖魚類(王凌宇等,2020)。有研究表明,大壩的建成通常導(dǎo)致原有江段魚類多樣性下降,繁殖魚類群落結(jié)構(gòu)趨于單一,其中條等小型魚類等易成為優(yōu)勢種(李世健等,2011;常濤等,2021),但鮮見黃顙魚成為優(yōu)勢種的報(bào)道。特鯰屬魚類監(jiān)測到中間特鯰(T.intermedia),中間特鯰隸屬于鲇形目頸鰭鯰科(Auchenipteridae),俗名為銀河豹鯨,在南美洲廣泛分布(Pereira et al.,2017),F(xiàn)ishbase數(shù)據(jù)庫標(biāo)記我國無分布,但在公開網(wǎng)絡(luò)上可檢索到國內(nèi)已將其作為觀賞魚進(jìn)行人工繁育的記錄。故推測中間特鯰是因遺棄、放生等方式進(jìn)入我國自然水域,但其為熱帶魚類,紅楓湖和阿哈湖的環(huán)境并不利于中間特鯰發(fā)展成為優(yōu)勢種。
既往研究顯示,序列豐度讀數(shù)和生物量存在一定的相關(guān)性(Kelly et al.,2014;Evans et al.,2016)。陳世靜(2020)研究表明,在魚類組成相對(duì)簡單的水族箱中,魚類生物量與序列讀數(shù)呈正相關(guān)。在PCR擴(kuò)增過程中,通用引物對(duì)水環(huán)境中不同物種的擴(kuò)增效率也不同,是由于通用引物對(duì)個(gè)別物種存在擴(kuò)增偏好性,而導(dǎo)致個(gè)別物種序列讀數(shù)產(chǎn)生數(shù)量級(jí)差異(Elbrecht and Leese,2015;Pi?ol et al.,2015)。因此,黃顙魚屬、鱯屬、特鯰屬魚類代表OTUs序列讀數(shù)較高,也有可能是通用引物的擴(kuò)增偏好性所造成。
3.3 紅楓湖和阿哈湖魚類多樣性與環(huán)境因子的關(guān)系
紅楓湖環(huán)境因子與魚類群落結(jié)構(gòu)差異的決定系數(shù)(R2)排序?yàn)镈O>TDS=WT>pH>NH4+。DO(P=0.034)是引起紅楓湖各采樣點(diǎn)魚類群落結(jié)構(gòu)差異的主要環(huán)境因子。DO是影響魚類生存最重要的環(huán)境因子,當(dāng)水體DO在2.0~3.0 mg/L時(shí)會(huì)影響魚類攝食等生命活動(dòng),而DO高于5.0 mg/L時(shí)魚類攝食生長完全正常(申玉春,2008)。本研究的采樣均在日出時(shí)間,紅楓湖各采樣點(diǎn)水體DO在6.3~10.5 mg/L,可能是由于日出時(shí)間湖中生產(chǎn)者正在進(jìn)行光合作用,因此水體DO均高于5.0 mg/L;在日落時(shí)間因光合作用停止,紅楓湖部分區(qū)域水體DO降至5.0 mg/L以下,致使DO成為紅楓湖魚類群落分布的主要制約因子。阿哈湖環(huán)境因子與魚類群落結(jié)構(gòu)差異的決定系數(shù)(R2)排序?yàn)門DS>W(wǎng)T>Tra>NH4+>DO。TDS(P=0.005)是引起阿哈湖各采樣點(diǎn)魚類群落結(jié)構(gòu)差異的主要環(huán)境因子。據(jù)李美霞(2021)的研究顯示,TDS和浮游植物群落呈顯著正相關(guān)。本研究中,阿哈湖以鰱和鳙的OTUs序列豐度最高,且二者都是以浮游生物為主要食物來源的濾食性魚類。這可能也是TDS成為引起阿哈湖各樣點(diǎn)魚類群落結(jié)構(gòu)差異主要環(huán)境因子的原因。
4 結(jié)論
現(xiàn)階段的紅楓湖和阿哈湖魚類組成以鯉形目魚類為主,但存在以日鱸目為主的養(yǎng)殖魚類生態(tài)入侵。eDNA宏條形碼技術(shù)在喀斯特高原水域魚類多樣性評(píng)估方面具有較好的適應(yīng)性,可快速監(jiān)測到魚類群落結(jié)構(gòu)和空間分布情況,但通用引物可能對(duì)某些魚類存在擴(kuò)增偏好性;此外,不同人工湖泊水質(zhì)理化因子特異性較強(qiáng),研究環(huán)境因子與湖泊魚類群落的關(guān)系時(shí)應(yīng)對(duì)每個(gè)湖泊單獨(dú)進(jìn)行評(píng)估。
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學(xué)學(xué)報(bào)(自然科學(xué)版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀(jì)智能(英語備考)(2019年12期)2020-01-13 06:07:18
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
