基于柑橘葉斑駁病毒的表達載體構建及應用
2022-02-01 02:21:44張琦段玉蘇越蔣琪琪王春慶賓羽宋震
中國農業科學 2022年22期
關鍵詞:煙草
張琦,段玉,蘇越,蔣琪琪,王春慶,賓羽,宋震
基于柑橘葉斑駁病毒的表達載體構建及應用
張琦,段玉,蘇越,蔣琪琪,王春慶,賓羽,宋震
西南大學柑桔研究所/國家柑桔工程技術研究中心,重慶 400712
【目的】構建基于柑橘葉斑駁病毒(citrus leaf blotch virus,CLBV)的表達載體,通過系統表達抗菌肽提高植物抗病性,為柑橘潰瘍病、柑橘黃龍病等病害的防控提供新型技術手段。【方法】基于前期構建的侵染性克隆pCY-CLBV201,在外殼蛋白基因終止子后插入亞基因組啟動子序列及多克隆位點,構建病毒表達載體pCLBV202。在多克隆位點插入綠色熒光蛋白(green fluorescent protein)基因(),通過農桿菌介導接種、熒光觀察驗證pCLBV202-GFP表達GFP的情況。克隆天蠶的抗菌肽(cecropin B,CB)基因并構建重組載體pCLBV202-CB,通過農桿菌介導分別注射接種本氏煙和真空浸潤接種柑橘實生苗,篩選陽性植株并分別注射接種和根灌接種煙草青枯病菌及針刺離體葉片接種柑橘潰瘍病菌,同時設空載體接種植株為對照,通過癥狀觀察、發病率及病情指數評價接種植株的煙草青枯病抗性;通過柑橘葉片的病斑數量、發病率及菌落濃度評價其潰瘍病抗性。【結果】pCLBV202-GFP接種煙草和尤力克檸檬后,均可以在系統新葉上觀察到綠色熒光,在煙草上表現更為明亮,說明基于CLBV的表達載體構建成功。接種青枯病菌后,處理組(pCLBV202-CB)較對照組(pCLBV202)發病時間延遲4 d。在接種后第24天(24 dpi),處理組發病率為14.3%,對照組發病率為100%,差異顯著。處理組相對于對照組的抗性指數為-2.66,抗性評價為高抗,表明利用pCLBV202-CB系統表達CB增強了對煙草青枯病的抗性。尤力克檸檬葉片針刺接種柑橘潰瘍病菌,7 dpi時,處理組病斑數為47個,發病率為43.5%,對照組病斑個數為73個,發病率為67.6%。菌群數變化檢測發現,處理組菌群數小于對照組菌群數,表明利用pCLBV202-CB系統表達CB增強了尤力克檸檬的潰瘍病抗性。【結論】構建了基于CLBV的病毒表達載體pCLBV202。利用pCLBV202在本氏煙和柑橘中系統表達CB可以提高植株對細菌性病害的抗性,這為柑橘細菌性病害的防控提供了新技術。
柑橘葉斑駁病毒;病毒表達載體;抗菌肽;抗病性
0 引言
【研究意義】柑橘是一種重要的經濟作物。近幾十年來,受全球氣候變暖等不利環境因素的影響,病蟲害尤其是黃龍病、潰瘍病對全球柑橘產業構成了嚴重威脅[1-2]。抗病育種是應對病蟲害危害的重要途徑。目前,通過轉基因技術已獲得許多抗病性顯著增強的柑橘轉基因材料[3],導入的抗病基因主要為來自昆蟲的抗菌肽基因和植物的抗病基因。抗菌肽作為天然抗生素可以直接殺死微生物[4],而天蠶素B(cecropin B,CB)是一種滅菌活性很強的抗菌肽,在柑橘轉基因抗病育種和病害防控中均得到了應用[5]。但柑橘轉基因技術難度大、周期長,并受到品種種類的限制。植物病毒表達載體作為一類極具潛力的外源基因表達平臺,可以快速在植物體內表達抗病基因,有望成為一種新型的病害防控手段。【前人研究進展】目前,抗菌肽基因在柑橘中的表達研究取得了一定進展,已將其成功導入多個品種的柑橘植株[6-7]。劉琦琦[8]研究表明,通過轉基因表達CB的柑橘植株能顯著提高其對柑橘潰瘍病的抗病能力。果樹病毒載體如柑橘衰退病毒(citrus tristeza virus,CTV)[9]、蘋果潛隱球形病毒(apple latent spherical virus,ALSV)[10]等能侵染果樹等多年生的植物,可用于在果樹等多年生植物中系統表達抗病基因[11]。CTV載體在柑橘上表達外源基因具有表達量高、穩定性強等優點[12-13],但其會通過蚜蟲傳播,存在病毒病擴散的風險。柑橘碎葉病毒(citrus tatter leaf virus,CTLV)已被構建成侵染性克隆[14-15]。但CTLV侵染以枳及其雜種為砧木的柑橘植株后引起嫁接部位腫大及褐色壞死,導致樹體生長衰弱和產量降低[16-17]。柑橘葉斑駁病毒(citrus leaf blotch virus,CLBV自然寄主廣泛,不僅能侵染大多數柑橘品種,也可侵染獼猴桃、牡丹、桑葚等其他物種[18-20],并且在柑橘多數栽培品種中不引發明顯癥狀,無昆蟲傳播媒介,適宜于植物病毒表達載體的開發[21]。目前,國外學者確定了CLBV外殼蛋白基因亞基因組RNA啟動子位點[22-23]。Vives等將CLBV的全長cDNA插入改進后的載體中,獲得了CLBV的全長侵染性克隆,并構建了病毒表達載體[10,24]。國內西南大學崔甜甜[25]構建了適用于農桿菌介導的CLBV侵染性克隆。【本研究切入點】在前期構建獲得CLBV侵染性克隆的基礎上,通過在其外殼蛋白(coat protein,CP)基因終止子后插入亞基因組啟動子序列及多克隆位點構建表達載體并利用該載體在本氏煙()和柑橘中系統表達抗菌肽,進而進行抗病性評價。【擬解決的關鍵問題】構建基于CLBV的病毒表達載體pCLBV202,通過在煙草與柑橘中系統表達CB,提高植株抗病性,為柑橘細菌性病害的防控提供新方法。
1 材料與方法
試驗于2019—2021年在西南大學柑桔研究所脫毒課題組完成。
1.1 試驗材料
尤力克檸檬()、本氏煙、柑橘葉斑駁病毒的侵染性克隆(pCY-CLBV201)、煙草青枯病菌()、柑橘潰瘍病菌(subsp.,)均由西南大學柑桔研究所國家柑桔苗木脫毒中心提供。
1.2 引物設計與基因合成
根據CLBV201侵染性克隆酶切位點的特性,利用Primer 5.0軟件設計擴增亞基因組啟動子序列的特異性引物92bpf1/92bpr1,以及用于插入載體目的片段檢測、擴增與測序的引物(3+1)F/(3+1)R。CLBV的通用檢測引物參考段玉等[26]的CLBV1F/CLBV5R。CB基因由上海英濰捷基貿易有限公司合成。利用引物5-F/5-R和3-F/3-R分別擴增I和I雙酶切pCY-CLBV201所獲小片段的5′端(1 904 bp)和3′端(658 bp)序列。gfpf1/gfpr1用于的擴增,引物序列如表1所示。

表1 試驗所用引物序列
1.3 pCLBV202表達載體構建
1.3.1 pCLBV202表達載體構建原理 采用I和I雙酶切pCY-CLBV201,將酶切下的小片段2 563 bp,自位點1 904處分成兩段,分別命名為5′端(1 904 bp)和3′端(658 bp)。利用引物5-F、5-R和3-F、3-R分別擴增5′端和3′端。CLBV外殼蛋白亞基因組啟動子序列大小92 bp,位于CLBV基因組的6 789—6 880,參照Agüero等[24]設計引物92bpf1/92bpr1。利用設計的帶同源臂的引物gfpf1/gfpr1從本實驗室的TuMV-GFP擴增回收基因片段。在設計引物時,除了同源臂外,在的兩端添加I酶切位點,便于目的基因的更換,具體如圖1所示。
1.3.2 CLBV201質粒酶切 將陽性的侵染性克隆菌株pCY-CLBV201,按照TIANprep Mini Plasmid Kit試劑盒的質粒提取步驟提取質粒,酶切體系:pCY- CLBV201質粒1.0 μg,10×Cutsmart Buffer 5 μl,I和I各1 μl,加入無菌水補足50 μl,25℃溫育30 min。酶切產物用0.8%的瓊脂糖凝膠電泳分離,進行切膠回收。
1.3.3 亞基因組啟動子序列、載體酶切片段、、片段的擴增與重疊 PCR擴增體系:2 μl模板,2×PrimeStar Max反應液10 μl,上下游引物各0.2 μl,加入無菌水7.2 μl。將5個片段單獨擴增出來,通過2次重疊PCR連接4個片段。92 bp+gfp重疊PCR:以92 bp亞啟動子和gfp膠回收產物各1 μl為模板,2×PrimeStar Max反應液10 μl,加入無菌水6 μl,94℃預變性5 min,94℃變性5 s,60℃解鏈30 s,72℃延伸90 s,10個循環后立即將反應液置于冰上,向反應體系中加入引物92bpf1/gfpr1各1 μL,置于PCR儀,94℃預變性5 min,94℃變性30 s,62℃解鏈30 s,72℃延伸90 s,35個循環,72℃再延伸5 min,電泳驗證后切膠回收。5′端+92 bp+gfp +3′端重疊PCR:以92 bp+gfp、5′端片段、3′端片段各1 μl,2×PrimeStar Max反應液10 μl,加入無菌水6 μl,94℃預變性5 min,94℃變性5 s,60℃解鏈30 s,72℃延伸90 s,10個循環后立即將反應液置于冰上,向反應體系中加入引物5-F/3-R各1 μL,置于PCR儀,94℃預變性5 min,94℃變性30 s,62℃解鏈30 s,72℃延伸90 s,35個循環,72℃再延伸5 min,電泳驗證后切膠回收。
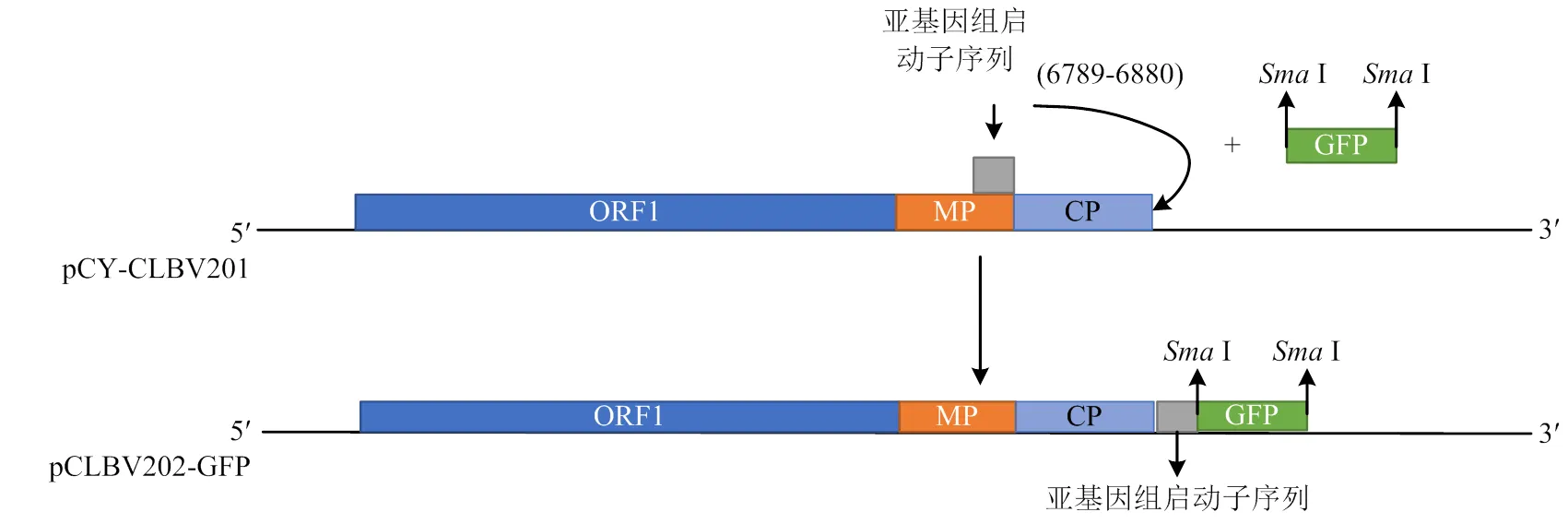
圖1 pCLBV202-GFP表達載體結構圖
1.3.4 pCLBV202-GFP、pCLBV202-CB重組表達載體的構建 目的片段100 ng,pCY-CLBV201酶切質粒200 ng,5×In-Fusion HD Enzyme Premix反應液2 μL,加入無菌水補足10 μl,在50℃中反應50 min,置于冰上,再將連接產物轉化大腸桿菌(),將測序成功的大腸桿菌提取質粒,電轉農桿菌。菌液PCR體系:菌液2 μl,2×Taq Master Mix反應液10 μl,上下游引物CLBV1F/CLBV5R各0.4 μl,無菌水7.2 μl。菌液PCR程序:94℃預變性5 min,94℃變性30s,55℃解鏈30 s,72℃延伸35 s,35個循環,72℃再延伸5 min。
1.3.5 農桿菌介導接種本氏煙和尤力克檸檬 農桿菌菌液在含Rif+和Kan+的平板上劃線,28℃培養2 d。挑取單克隆于Rif+和Kan+的LB培養液中,28℃,220 r/min振蕩培養2 d。按照1﹕50的比例將活化后的農桿菌菌液接種于農桿菌擴繁培養基中,相同條件下培養菌液至OD600=0.9—1.1。8 000 r/min離心8 min使菌液富集,用接種緩沖液懸浮(10 mmol·L-1MgCl2,10 mmol·L-1MES,200 μmol·L-1As),使其OD600為0.8—1.0,同時加入沉默抑制子P19表達克隆,使樣品OD600≈1.0,沉默抑制子OD600≈0.5,黑暗靜置2 h 備用。農桿菌注射接種:從葉背面注射本氏煙葉片,至直徑約為1 cm。每菌株至少注射3株植物的3個葉片。農桿菌根灌接種:用7 mL離心管沿著本氏煙的根部澆灌5 mL農桿菌。農桿菌真空浸潤:將清洗干凈的柑橘幼苗放入接種菌液并置于真空干燥儀中,抽真空至壓力-1.0—-0.8 kg·cm-2,保持5 min后快速釋放壓力并立即用無菌水沖洗,將植株移栽于土床中,28℃黑暗處理2 d,即可移栽于光照培養箱中。培養箱參數設置:光照時長16 h,23℃;黑暗8 h,20℃。待長出2—4片新葉后便可取樣檢測。
1.4 pCLBV202-CB對煙草青枯病的防治效果
1.4.1 pCLBV202-CB和pCLBV202接種本氏煙 本氏煙在長出第5片針葉后進行農桿菌注射接種,處理組pCLBV202-CB接種9株,對照組pCLBV202接種9株,具體接種步驟詳見1.3.5。
1.4.2 煙草青枯菌活化與接種 將存于水中的青枯病菌涂布于TTC平板上,28℃培養48 h;挑取單克隆菌落接種于2 mL TTC培養基中,28℃250 r/min培養,監測菌液OD值,在OD值達到0.8之前停止培養細菌;轉移1 mL菌液至1.5 mL離心管,8 000 r/min離心2 min,去上清,集沉淀;用10 mmol·L-1的MgCl2洗滌沉淀3次,離心,最后重懸青枯病菌于10 mmol·L-1MgCl2,OD=0.001備用。在處理組和處理組18 dpi時,接種煙草青枯病菌。每株進行注射葉片接種及根灌接種農桿菌菌液5 mL。
1.4.3 煙草青枯病病情統計 發病率(%)=(發病株數/調查總株數)×100;病情指數(DI)=[Σ(病情級數×此級菌株數)/(總級數×總株數)]×100;青枯病病情分級標準參照《煙草病蟲害分級及調查方法》(GB/T 23222—2008)[27]。抗性指數RI=ln[DI/(100-DI)]-ln[DO/(100-DO)]。其中,DI指對照組病情指數;DO指處理組病情指數。RI≤0,表示高抗,用HR表示;0<RI≤1.00為抗病,用R表示;1.00<RI≤1.49為感病,用S表示,RI≥1.50為高感,用HS表示。
1.5 pCLBV202-CB對柑橘潰瘍病的防治效果
1.5.1 pCLBV202-CB和pCLBV202接種尤力克檸檬 在播種檸檬長至8—10 d時,通過真空浸潤處理組pCLBV202-CB 30株,對照組pCLBV202 30株,方法參照1.3.5。待檸檬植株長至30 d時,通過檢測篩選pCLBV202-CB、pCLBV202陽性植株。
1.5.2 潰瘍病菌離體針刺接種 取大小相近、生長狀況相似的尤力克檸檬葉片,處理組3片,對照組3片。發病率(%)=(病斑個數/接種個數)×100;菌群數變化檢測的具體操作參照Duan等[28]的方法。
2 結果
2.1 pCLBV202-GFP與pCLBV202-CB重組表達載體的構建
pCLBV202-GFP重組表達載體轉入大腸桿菌DH10B后,進行菌液PCR檢測,結果見圖2-A,檢測片段與目標片段大小一致。序列分析顯示,在CP基因終止子后插入了92 bp的亞基因組啟動子序列及其控制下的完整GFP開放讀碼框,GFP開放讀碼框兩端分別包含一個I酶切位點,證明pCLBV202-GFP重組表達載體構建成功。使用I雙酶切pCLBV202-GFP質粒構建pCLBV202-CB,轉化大腸桿菌DH10B。菌液PCR檢測結果如圖2-B所示,獲得大小約為732 bp的特異條帶,符合目標基因片段大小,經序列分析后,確定pCLBV202-CB重組載體構建成功。
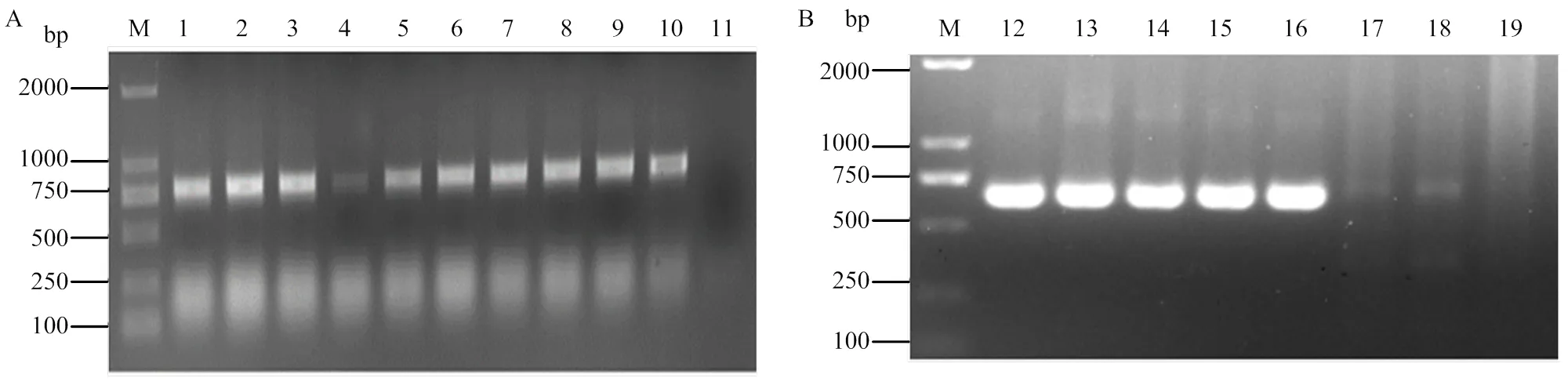
A:pCLBV202-GFP單克隆檢測pCLBV202-GFP monoclonal detection;B:pCLBV202-CB單克隆檢測pCLBV202-CB monoclonal detection。M:2000 bp標準分子量2000 bp molecular marker;1—10:pCLBV202-GFP轉化大腸桿菌單克隆pCLBV202-GFP transformed E. coli clones;12—18:pCLBV202-CB轉化大腸桿菌單克隆pCLBV202-CB transformed E. coli clones;11、19:水對照Water control
2.2 本氏煙和尤力克檸檬的熒光觀察
本氏煙經注射接種含pCLBV202-GFP重組表達載體的農桿菌后,11 dpi時,莖和根部出現綠色熒光,在15 dpi時,系統新葉出現強烈的綠色熒光,表明pCLBV202-GFP在本氏煙植株中完成系統侵染。隨后,在花中觀察到綠色熒光,說明構建的pCLBV202-GFP能夠在本氏煙植株中穩定表達外源的綠色熒光蛋白(圖3-A—3-E)。
尤力克檸檬經真空浸潤接種pCLBV202-GFP,30 dpi時通過共聚焦顯微鏡對系統葉進行綠色熒光觀察。結果表明,與空載體對照相比,處理組檸檬葉片有微弱的綠色熒光顆粒。表明構建的pCLBV202-GFP在尤力克檸檬體內也具有表達功能,能夠表達外源的綠色熒光蛋白(圖3-F)。
2.3 pCLBV202-CB對煙草青枯病的防治效果
處理組pCLBV202-CB和對照組pCLBV202接種煙草青枯病菌后,處理組發病時間為7 dpi,對照組發病時間為3 dpi,處理組比對照組發病晚4 d。在接種青枯病菌24 dpi后,處理組發病率為14.3%,對照組發病率為100%,處理組發病率遠低于對照組發病率。處理組相對于對照組的抗性指數為-2.66,抗性評價為高抗。處理組相較于對照組,癥狀出現明顯減輕,具體結果如圖4所示。這表明利用pCLBV202在煙草中系統表達CB增強了對煙草青枯病的抗性。
2.4 pCLBV202-CB對柑橘潰瘍病的防治效果
通過RT-PCR檢測CB蛋白基因是否在尤力克檸檬系統葉中存在轉錄,篩選陽性植株進行抗性評價。結果如圖5所示,共獲得了4株pCLBV202-CB陽性尤力克檸檬植株。
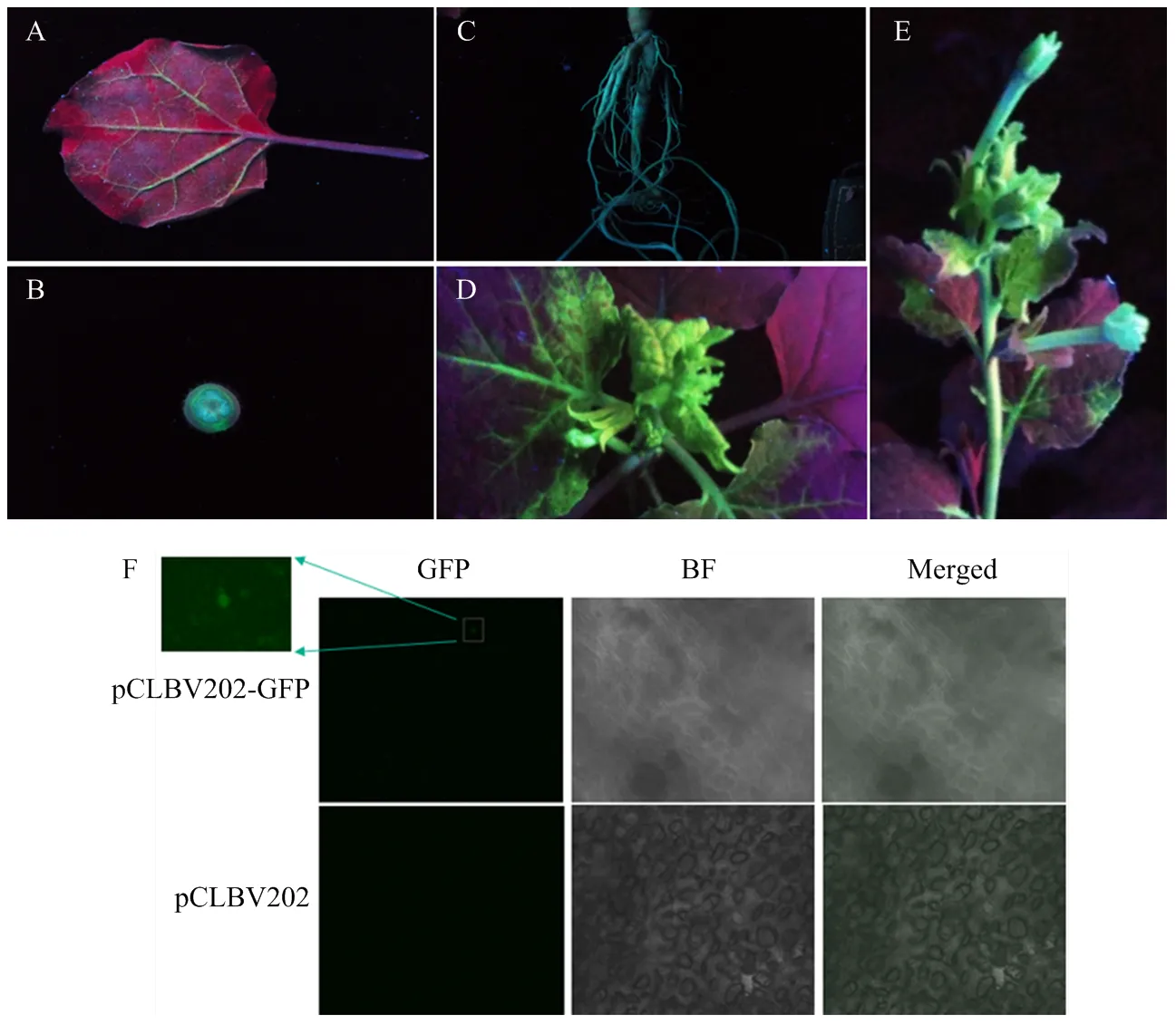
本氏煙N. benthamianaA:注射葉背面The reverse side of injection leaf (7 dpi);B:莖稈橫切面Cross-section of stalk (11 dpi);C:根莖Rhizome (11dpi);D:系統葉systemic leaves (15 dpi);E:花蕾Buds (18 dpi)。尤力克檸檬C. limon F:系統葉Systemic leaves (30 dpi)

A:煙草青枯病癥狀的整體圖overall picture of tobacco bacterial wilt symptoms;B:煙草青枯病分級classification of tobacco bacterial wilt;C:pCLBV202-CB組煙草青枯病癥狀tobacco bacterial wilt symptoms in pCLBV202-CB group;D:pCLBV202組煙草青枯病癥狀tobacco bacterial wilt symptoms in pCLBV202 group;E:pCLBV202-CB組注射葉injected leaf of pCLBV202-CB group;F:pCLBV202組注射葉injected leaf of pCLBV202 group
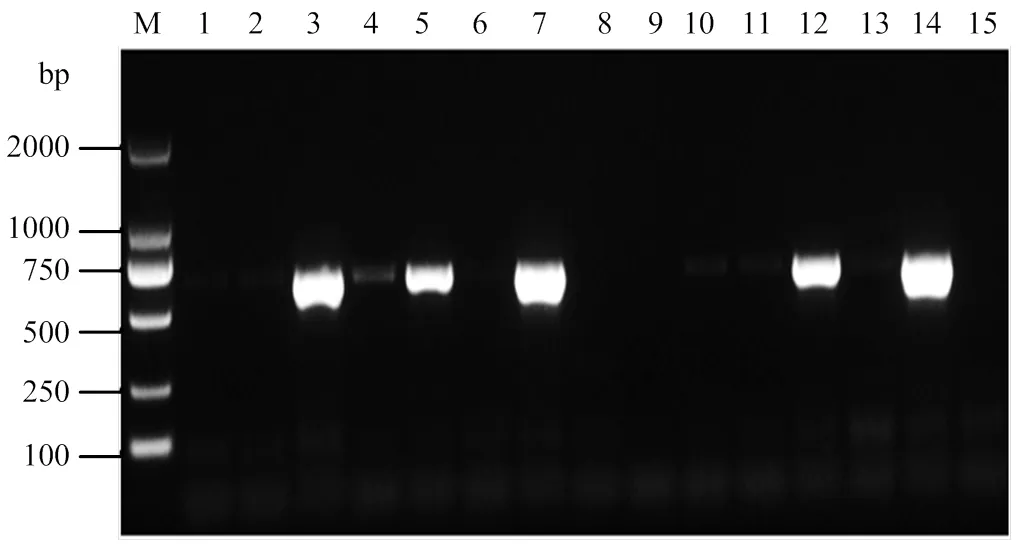
M:2000 bp標準分子量2000 bp molecular marker;1—12:pCLBV202-CB接種樣品pCLBV202-CB inoculated samples;13:陰性對照Negative control;14:陽性對照Positive control;15:水對照Water control
觀察記錄接種柑橘潰瘍病菌的尤力克檸檬葉片,在7 dpi時處理組和對照組均發病(圖6)。處理組病斑數為47個,發病率為43.5%;對照組病斑個數為73個,發病率為67.6%。

圖6 pCLBV202-CB對柑橘潰瘍病的防治效果
潰瘍菌斑研磨稀釋后進行細菌的回補培養,結果如圖7所示,處理組潰瘍病菌在稀釋51倍便停止生長;對照組的潰瘍病菌稀釋54倍停止生長。因此,處理組中潰瘍病菌菌群數少于對照組潰瘍病菌菌群數。
上述結果表明,利用pCLBV202-CB處理后的尤力克檸檬較對照的柑橘潰瘍病抗性增強。

1、2、3、4、5、6:潰瘍病菌的稀釋倍數分別為51、52、53、54、55、56倍Dilution gradients of Xcc were 51, 52, 53, 54, 55, 56 times, respectively
3 討論
3.1 pCLBV202-CB可提高植物對細菌性病害的抗性
柑橘產業是我國南方許多地區鄉村振興的支柱產業,但柑橘病害尤其是黃龍病、潰瘍病嚴重影響了該產業的健康發展[28]。通過轉基因技術將抗菌肽基因轉入柑橘中可以增強柑橘對細菌病害的抗病性,但柑橘轉基因周期長、操作繁瑣并且存在較高的技術難度和一定的品種限制。本研究基于前期構建的侵染性克隆pCY-CLBV201,通過在其外殼蛋白基因終止子后插入92 bp亞基因組啟動子序列及克隆位點,構建了病毒表達載體pCLBV202。抗菌肽cecropin B通過靜電作用抑制細菌細胞呼吸,細菌最后死亡或者停止生長,從而有效降低細菌病害的危害與傳播。李乃堅等[29]將天蠶抗菌肽B基因導入煙草品種‘Nc82’‘Col76’和‘K326’并獲得了抗性后代,其抗青枯病能力與受體品種相比有較大幅度的提高。本研究利用該載體在本氏煙中表達抗菌肽CB,顯著降低了煙草青枯病的發病率以及癥狀嚴重程度,提高了植物對煙草青枯病的抗性,這與李乃堅等[29]的結果一致;在尤力克檸檬植株體內表達CB也可顯著降低柑橘潰瘍病菌的病斑數和菌群數量,提高檸檬的柑橘潰瘍病抗性,這與之前關于轉基因抗菌肽的研究結果一致[30-32],這些結果均證明病毒表達載體在植物中系統的表達CB與轉基因植物具有同樣的抗細菌病害能力。
3.2 基于CLBV表達載體的優勢
由于果樹是多年生植物,因此構建能在果樹植株內長期、穩定表達外源基因的病毒表達載體具有積極意義。目前已獲得的果樹病毒載體中,CTV載體的穩定性最好,這可能與長線型病毒屬中所特有的HSP70同源蛋白和較低的重組率有關[33]。但CTV可以通過蚜蟲傳播[34],存在一定的病毒擴散風險。CLBV能夠侵染大多數柑橘栽培品種,不引起明顯的癥狀,且無昆蟲傳播媒介,從而適宜用于載體改造。已有研究表明基于CLBV的表達載體可以在柑橘中穩定表達GFP達7年之久,穩定性良好。插入的外源基因最終會丟失,不存在生物安全問題。因而,利用本研究構建的表達載體表達CB或其他更加高效的抗菌肽可使柑橘獲得較持久的細菌性病害抗性,減輕病害防控壓力。CLBV寄主廣泛,可以侵染獼猴桃、桑樹、牡丹等多種植物,因此,基于CLBV的表達載體也可能適用于更多的寄主,應用前景良好。另外,本研究所獲載體也可用于反向遺傳學研究,通過pCLBV202過表達外源基因,可快速對其功能進行研究,對于快速篩選抗性基因尋找更好的病害防控途徑具有積極意義。目前,利用pCLBV202在柑橘中表達外源基因的表達量仍不夠高,后續可通過篩選高滴度病毒、優選外源基因插入位點等研究繼續優化,從而提高其外源抗性基因的表達量,這對提升柑橘抗性以及病害防控效果極其重要。
4 結論
構建了基于CLBV的病毒表達載體pCLBV202。利用pCLBV202在本氏煙、柑橘中系統表達抗菌肽可顯著提高植株的細菌性病害抗性,為柑橘病害防控提供了新思路。
[1] 陳祝安, 曹光照, 許益偉, 黃基榮, 葛嵐屏. 柑桔害蟲病原真菌資源的考察和生測. 微生物學通報, 1985(5): 194-198.
CHEN Z A, CAO G Z, XU Y W, HUANG J R, GE L P. Investigation and bioassay of citrus pest pathogenic fungi resources. Microbiology China, 1985(5): 194-198. (in Chinese)
[2] BROWN K. Florida fights to stop citrus canker. Science, 2001, 292(5525): 2275-2276.
[3] 宋二玲. 三個病原物誘導啟動子在轉基因柑橘中受潰瘍病菌和創傷誘導的表達分析[D]. 重慶: 西南大學, 2013.
SONG E L. Canker bacterium- and wound-response characteristics of three pathogen-induced promoters in transgenic citrus[D]. Chongqing: Southwest University, 2013. (in Chinese)
[4] NARDO A D, VITIELLO A, GALLO R L. Cutting edge: Mast cell antimicrobial activity is mediated by expression of cathelicidin antimicrobial peptide. Journal of Immunology, 2003, 170(5): 2274-2278.
[5] ZASLOFF M. Antimicrobial peptides of multicellular organisms. Nature, 2002, 415(6870): 389-395.
[6] 鄭啟發, 陳大成, 黃自然, 胡桂兵. 人工合成柞蠶抗菌肽D基因轉化沙田柚. 華南農業大學學報, 1999, 20(1): 103-106. ZHENG Q F, CHEN D C, HUANG Z R, HU G B. Transformation of the Shatian pummelo () with the synthetic oak silkworm antibacterial peptide D gene. Journal of South China Agricultural University, 1999, 20(1): 103-106. (in Chinese)
[7] 陳善春, 張進仁, 黃自然, 高峰, 陳鳳珍, 隆有慶. 根癌農桿菌介導柞蠶抗菌肽D基因轉化柑桔的研究. 中國農業科學, 1997, 30(3): 7-13.
CHEN S C, ZHANG J R, HUANG Z R, GAO F, CHEN F Z, LONG Y Q. Studies on-mediated antibacterial peptide D gene transfer in citrus. Scientia Agricultura Sinica, 1997, 30(3): 7-13. (in Chinese)
[8] 劉琦琦. 轉cecropin B基因柑橘對潰瘍病抗性的研究[D]. 重慶: 西南大學, 2014.
LIU Q Q.resistance of transgenic plants with cecropin B gene to citrus canker caused bypv.[D]. Chongqing: Southwest University, 2014. (in Chinese)
[9] SATYANARAYANA T, GOWDA S, BOYKO V P, ALBIACH- MARTI M R, MAWASSI M, NAVAS-CASTILLO J, KARASEV A V, DOLJA V, HILF M E, LEWANDOWSKI D J, MORENO P, BAR-JOSEPH M, GARNSEY S M, DAWSON W O. An engineered closterovirus RNA replicon and analysis of heterologous terminal sequences for replication. Proceedings of the National Academy of Sciences of the United States of America, 1999, 96(13): 7433-7438.
[10] VIVES M C, MARTíN S, AMBRóS S, RENOVELL á, NAVARRO L, PINA J, MORENO P, GUERRI J. Development of a full-genome cDNA clone of citrus leaf blotch virus and infection of citrus plants. Molecular Plant Pathology, 2008, 9(6): 787-797.
[11] OHIRA K, NAMBA S, ROZANOV M, KUSUMI T, TSUCHIZAKI T. Complete sequence of an infectious full-length cDNA clone of citrus tatter leaf capillovirus: comparative sequence analysis of capillovirus genomes. Journal of General Virology, 1995, 76(9): 2305-2309.
[12] GOWDA S, SATYANARAYANA T, ROBERTSON C J, GARNSEY S M, DAWSON W O. Infection of citrus plants with virions generated inplants agroinfiltrated with a binary vector based citrus tristeza virus//Proceedings of the Sixteenth Conference of the International Organization of Citrus Virologist. Riverside: IOCV, 2005: 23-33.
[13] SáNCHEZ F, MARTINEZ-HERRERA D, AGUILAR I, PONZ F. Infectivity of turnip mosaic potyvirus cDNA clones and transcripts on the systemic hostand local lesion hosts. Virus Research, 1998, 55(2): 207-219.
[14] 周彥. 果樹病毒載體研究進展. 中國農業科學, 2014, 47(6): 1119-1127.
ZHOU Y. Progresses in study of virus-based vectors of fruit trees. Scientia Agricultura Sinica, 2014, 47(6): 1119-1127. (in Chinese)
[15] GALIAKPAROV N, TANNE E, SELA I, GAFNY R. Infectious RNA transcripts from grapevine virus A cDNA clone. Virus Genes, 1999, 19(3): 235-242.
[16] VELA L B, SKARIA M. Seven isolates of citrus tatter leaf virus induce varying levels of xylem tissue abnormalities in two citrange rootstocks//Proceedings of the Fifteenth Conference of the International Organization of Citrus Virologists. Riverside: IOCV, 2002: 371-372.
[17] MIYAKAWA T, MATSUI C. A bud-union abnormality of Satsuma mandarin onrootstock in Japan//Proceedings of the Seventh Conference of the International Organization of Citrus Virologists. Riverside: IOCV, 1976: 125-131.
[18] CAO M J, YU Y Q, TIAN X, YANG F Y, LI R H, ZHOU C Y. First report of citrus leaf blotch virus in lemon in China. Plant Disease, 2017, 101(8): 1561.
[19] CHAVAN R R, BLOUIN A G, COHEN D, PEARSON M N. Characterization of the complete genome of a novel citrivirus infecting. Archives of Virology, 2013, 158(8): 1679-1686.
[20] XUAN Z Y, XIE J X, YU H D, ZHANG S, LI R H, CAO M J. Mulberry () is a new natural host of citrus leaf blotch virus in China. Plant Disease, 2021, 105(3): 716.
[21] VIVES M C, GALIPIENSO L, NAVARRO L, MORENO P, GUERRIL J. The nucleotide sequence and genomic organization of citrus leaf blotch virus: candidate type species for a new virus genus. Virology, 2001, 287(1): 225-233.
[22] AGüERO J, VIVES M C, VELáZQUEZ K, RUIZ-RUIZ S, JUáREZ J, NAVARRO L, MORENO P, GUERRI J. Citrus leaf blotch virus invades meristematic regions inand citrus. Molecular plant pathology, 2013, 14(6): 610-616.
[23] RENOVELLá, GAGO S, RUIZ-RUIZ S, VELáZQUEZ K, NAVARRO L, MORENO P, VIVES M C, GUERRI J. Mapping the subgenomic RNA promoter of the citrus leaf blotch virus coat protein gene by-mediated inoculation. Virology, 2010, 406(2): 360-369.
[24] AGüERO J, RUIZ-RUIZ S, VIVES M C, VELáZQUEZ K, NAVARRO L, Pe?a L, MORENO P, GUERRI J. Development of viral vectors based on citrus leaf blotch virus to express foreign proteins or analyze gene function in citrus plants. Molecular plant-microbe interactions, 2012, 25(10): 1326-1337.
[25] 崔甜甜. 柑橘黃化脈明病毒和柑橘葉斑駁病毒的侵染性克隆構建[D]. 重慶: 西南大學, 2018.
CUI T T. Construction of infectious cDNA clones of citrus yellow vein clearing virus and citrus leaf blotch virus[D]. Chongqing: Southwest University, 2018. (in Chinese)
[26] 段玉, 許建建, 馬志敏, 賓羽, 周常勇, 宋震. 柑橘葉斑駁病毒的逆轉錄重組酶聚合酶擴增檢測. 中國農業科學, 2021, 54(9): 1904-1912.
DUAN Y, XU J J, MA Z M, BIN Y, ZHOU C Y, SONG Z. Detection of citrus leaf blotch virus by reverse transcription-recombinase polymerase amplification (RT-RPA). Scientia Agricultura Sinica, 2021, 54(9): 1904-1912. (in Chinese)
[27] 任廣偉, 孔凡玉, 王鳳龍. 煙草病蟲害分級及調查方法: GB/T 23222—2008[S]. (2008-12-31) [2022-07-04].
REN G W, KONG F Y, WANG F L. Grade and investigation method of tobacco diseases and insect pests: GB/T 23222—2008[S]. (2008- 12-31) [2022-07-04]. (in Chinese)
[28] DUAN S, JIA H G, PANG Z Q, TEPER D, WHITE F, JONES J, ZHOU C Y, WANG N. Functional characterization of the citrus canker susceptibility gene. Molecular plant pathology, 2018, 19(8): 1908-1916.
[29] 李乃堅, 袁四清, 蒲漢麗, 周會光, 戴冕, 陳俊標, 羅戰勇, 崔植琳, 黃自然, 李傳瑛, 陳鳳珍. 抗菌肽B基因轉化煙草及轉基因植株抗青枯病的鑒定. 農業生物技術學報, 1998, 6(2): 178-184.
LI N J, YUAN S Q, PU H L, ZHOU H G, DAI M, CHEN J B, LUO Z Y, CUI Z L, HUANG Z L, LI C Y, CHEN F Z. Transfer of cecropin B gene into tobacco plant and confer the bacterial wilt resistance. journal of agricultural biotechnology, 1998, 6(2): 178-184. (in Chinese)
[30] 黎觀紅, 洪智敏, 賈永杰, 瞿明仁. 抗菌肽的抗菌作用及其機制. 動物營養學報, 2011, 23(4): 546-555.
LI G H, HONG Z M, JIA Y J, QU M R. Activities and mechanisms of action of antimicrobial peptides. Chinese Journal of Animal Nutrition, 2011, 23(4): 546-555. (in Chinese)
[31] 王志興, 賈士榮. 抗菌肽分泌型載體的構建及轉基因馬鈴薯中蛋白的胞外分泌. 農業生物技術學報, 1996, 4(3): 277-286.
WANG Z X, JIA S R. Construction of vectors harboring a barley-amylase signal sequence and a cecropin B or shiva A gene that resulted in the extracellular protein secretion in potato transgenic plants. journal of agricultural biotechnology, 1996, 4(3): 277-286. (in Chinese)
[32] 賈士榮, 屈賢銘, 馮蘭香, 唐惕, 唐益熊, 劉坤, 趙艷麗, 白永延, 蔡敏鶯. 轉抗菌肽基因提高馬鈴薯對青枯病的抗性. 中國農業科學, 1998, 31(3): 5-12.
JIA S R, QU X M, FENG L X, TANG T, TANG Y X, LIU K, ZHAO Y L, BAI Y Y, CAI M Y. Development of potato clones with enhanced resistance to bacterial wilt by introducing antibacterial peptide gene. Scientia Agricultura Sinica, 1998, 31(3): 5-12. (in Chinese)
[33] Kurth E G, Peremyslov V V, Prokhnevsky A I, Kasschau K D, Miller M, Carrington J C, Dolja V V. Virus-derived gene expression and RNA interference vector for grapevine. Journal of Virology, 2012, 86(11): 6002-6009.
[34] Zhou Y, Zhou C Y, Song Z, Liu K H, Yang F Y. Characterization of citrus tristeza virus isolates by indicators and molecular biology methods. Agricultural Sciences in China, 2007, 6(5): 573-579.
Construction and application of expression vector based on Citrus leaf blotch virus
ZHANG Qi, DUAN Yu, SU Yue, JIANG QiQi, WANG ChunQing, BIN Yu, SONG Zhen
Citrus Research Institute, Southwest University/National Citrus Engineering Research Center, Chongqing 400712
【Objective】The objective of this study is to construct an expression vector based on citrus leaf blotch virus (CLBV) and systematically express antimicrobial peptides to improve plant disease resistance, which will provide a new technical means for the prevention and control of citrus canker, citrus Huanglongbing (HLB) and other diseases.【Method】The subgenomic promoter sequence and multiple cloning sites were inserted after the terminator of the coat protein gene of CLBV to construct the viral expression vector pCLBV202 based on the previously constructed infectious clone pCY-CLBV201. Then green fluorescent proteingene () was inserted into the multiple cloning sites, and the expression of GFP was verified by-mediated inoculation and fluorescence observation. the cecropin B (CB) gene ofwas cloned and the recombinant vector pCLBV202-CB was constructed.and citrus seedlings were inoculated by pCLBV202-CB using-mediated injection and vacuum infiltration, respectively. Positive plants were screened and subjected to inoculation ofby injection and root irrigation orsubsp.() by pricking the detached leaves. At the same time, the plants inoculated with the empty vector were set as the control. The resistance to tobacco bacterial wilt of the inoculated plants was evaluated by symptom observation, incidence rate and disease index. The citrus resistance to canker was evaluated by the detriment, incidence rate and colony concentration on leaves. 【Result】After inoculated by pCLBV202-GFP, green fluorescence could be observed on the systemic leaves of theand, which was brighter in, indicating that the expression vector based on CLBV was successfully constructed. After inoculation of, the onset time of treatment group (pCLBV202-CB) was delayed by 4 days compared with the control group (pCLBV202). On the 24th day past inoculation (24 dpi), the incidence rate of the treatment group was 14.3%, and that of the control group was 100%. Compared with the control group, the resistance index of the treatment group was -2.66, and the resistance evaluation was high, indicating that the expression of CB by pCLBV202-CB enhanced the resistance to tobacco bacterial wilt. Whenleaves was inoculated by, the number of detriment in the treatment group at 7 dpi was 47, and the incidence rate was 43.5%, while that of the control group was 73, and 67.6%. The detection of colony concentration changes ofshowed that the number ofin the treatment group was less than that in the control group, indicating that the expression of CB by pCLBV202-CB enhanced the resistance to citrus canker.【Conclusion】The CLBV-based viral expression vector pCLBV202 was constructed. Using pCLBV202 to systematically express CB inand citrus can improve the resistance of plants to bacterial diseases, which provided a new technology for the prevention and control of citrus bacterial diseases.
citrus leaf blotch virus (CLBV);viral expression vector;antimicrobial peptide;disease resistance

10.3864/j.issn.0578-1752.2022.22.006
2022-07-04;
2022-07-16
國家重點研發計劃(2021YFD1400800)、重慶市自然科學基金(CSTB2022NSCQ-MSX0752)
張琦,E-mail:18839773525@163.com。段玉,E-mail:982432080@qq.com。張琦和段玉為同等貢獻作者。通信作者宋震,E-mail:songzhen@cric.cn
(責任編輯 岳梅)
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
