基于GFM和GAMM模型分析對蝦白斑綜合征(WSSV)對黃海和東海北部水域蝦類生物量的影響*
2022-01-14 07:48:12徐婷婷滕廣亮李英瑕吳強單秀娟2張慶利金顯仕
漁業科學進展 2022年1期
關鍵詞:模型
徐婷婷滕廣亮李英瑕吳 強單秀娟,2張慶利,2① 金顯仕,2
(1.中國水產科學研究院黃海水產研究所 農業農村部海水養殖病害防治重點實驗室青島市海水養殖流行病學與生物安保重點實驗室 山東 青島 266071;2.青島海洋科學與技術試點國家實驗室海洋漁業科學與食物產出過程功能實驗室 山東 青島 266071)
中國近海漁業資源種類豐富,魚類和蝦類資源在漁業生態系統中占據著重要地位(唐啟升, 2006;吳強等, 2012)。自20世紀80年代以來,受過度捕撈、氣候變化、環境污染等多重壓力的影響,黃東海生態系統發生明顯的變化,傳統經濟魚類資源出現一定程度的衰退,包括蝦類在內的甲殼類資源所占比例明顯升高(Jin et al, 2013; Tang et al, 2016; Wu et al, 2018)。近期的相關研究表明,受人類海洋活動日趨頻繁和全球氣候變化加劇的影響,漁業資源種群演變更加復雜(Ramos et al, 2018; Baltar et al, 2019; Ward et al, 2019;Walworth et al, 2020)。除捕撈因素外,近海甲殼類動物的生物量還受水深、底層溫度、鹽度、經緯度、底質類型和離岸距離等因子的影響(Gribble et al, 2007;黃慶洋等, 2009; 逄志偉等, 2015; 趙靜等, 2017;張孝民等, 2018)。蝦類作為海洋生態系統中重要的次級生產者,是構建食物網結構、解析能量流動和物質循環過程中必不可缺的關鍵環節(陳小慶等,2010;梁金玲,2015)。因此,探討海洋中蝦類生物量變化及其影響因素對近海生物種群動態和海洋生態學研究有重要意義。
20世紀80年代以來,對蝦養殖業在世界多國蓬勃興起,為經濟發展和優質動物蛋白供應做出了突出貢獻(余云軍,2006)。然而,與全球對蝦養殖業快速發展相伴的是多種病毒性疫病不斷出現,并對養殖產業造成沖擊(李建軍等,1998)。其中,對蝦白斑綜合征病毒(white spot syndrome virus, WSSV)是已報道的對蝦病毒性疫病中危害嚴重且流行區域最廣的一類病原(Lightner,1996;何建國等,1999;雷質文等,2002、2003;閆冬春, 2004;李鈁, 2009)。池塘養殖對蝦感染WSSV后2~7d內的累計死亡率可達100%(雷質文等,2003)。除了感染養殖對蝦外,近年來陸續有WSSV感染近海甲殼類動物的報道。如Chapman等(2004)通過PCR、組織病理學和組織原位雜交實驗證實捕獲自南大西洋灣的凡納濱對蝦(Litopenaeus vannamei)樣品中存在 WSSV感染。Mijangos-Alquisires等(2006)從墨西哥加利福尼亞灣采集的近海蝦類樣品中檢測到WSSV較低陽性率(0.8%)的感染。Orosco等(2017)通過PCR檢測表明,采集自菲律賓近岸水域的蝦類中有WSSV陽性檢出。Gholamhosseini等(2020)通過PCR、組織病理學實驗和透射電子顯微鏡技術確定了伊朗近海印度明對蝦(Penaeusindicus)中存在WSSV。鑒于WSSV對甲殼類的廣泛致病性,近海甲殼類樣品中存在WSSV流行不免引發其可能危害野生蝦類種群動態的擔憂。
基于此,本研究于2016—2018年每年8月對WSSV在黃海和東海北部蝦類中的流行情況進行系統調查,通過構建梯度隨機森林模型(gradient random forest model,GFM)和廣義加性混合模型(generalized additive mixed models, GAMM),評估WSSV流行情況及相關環境因子與調查海域蝦類生物量的相關性,以期為揭示水產養殖疫病病原對海洋甲殼類資源的影響提供新的視角。
1 材料與方法
1.1 樣品采集
2016、2017和 2018年 8月(休漁期),搭乘中國水產科學研究院黃海水產研究所“北斗號”漁業調查船赴黃海和東海北部海域進行3個航次的調查,并利用底拖網采集蝦類樣品。根據臨近站位間的距離和實時海況,在預設的120個站點中選擇站點進行取樣(圖1)。根據底拖網所獲蝦的種類和生物量,每個采樣站點隨機取樣4~6尾。取樣時,將樣本個體的頭胸甲剖開涂抹于FTA?卡片上,自然晾干后保存于–20℃冰箱,以備后續檢測。調查網具參數見單秀娟等(2011),拖速為3.0~3.5 nmile/h,拖網時間0.5~1 h。記錄各站點捕獲蝦的種類和數量,將不同種蝦分別標準化為每小時的數量(個/h)作為其種群生物量(陳云龍等, 2013)。
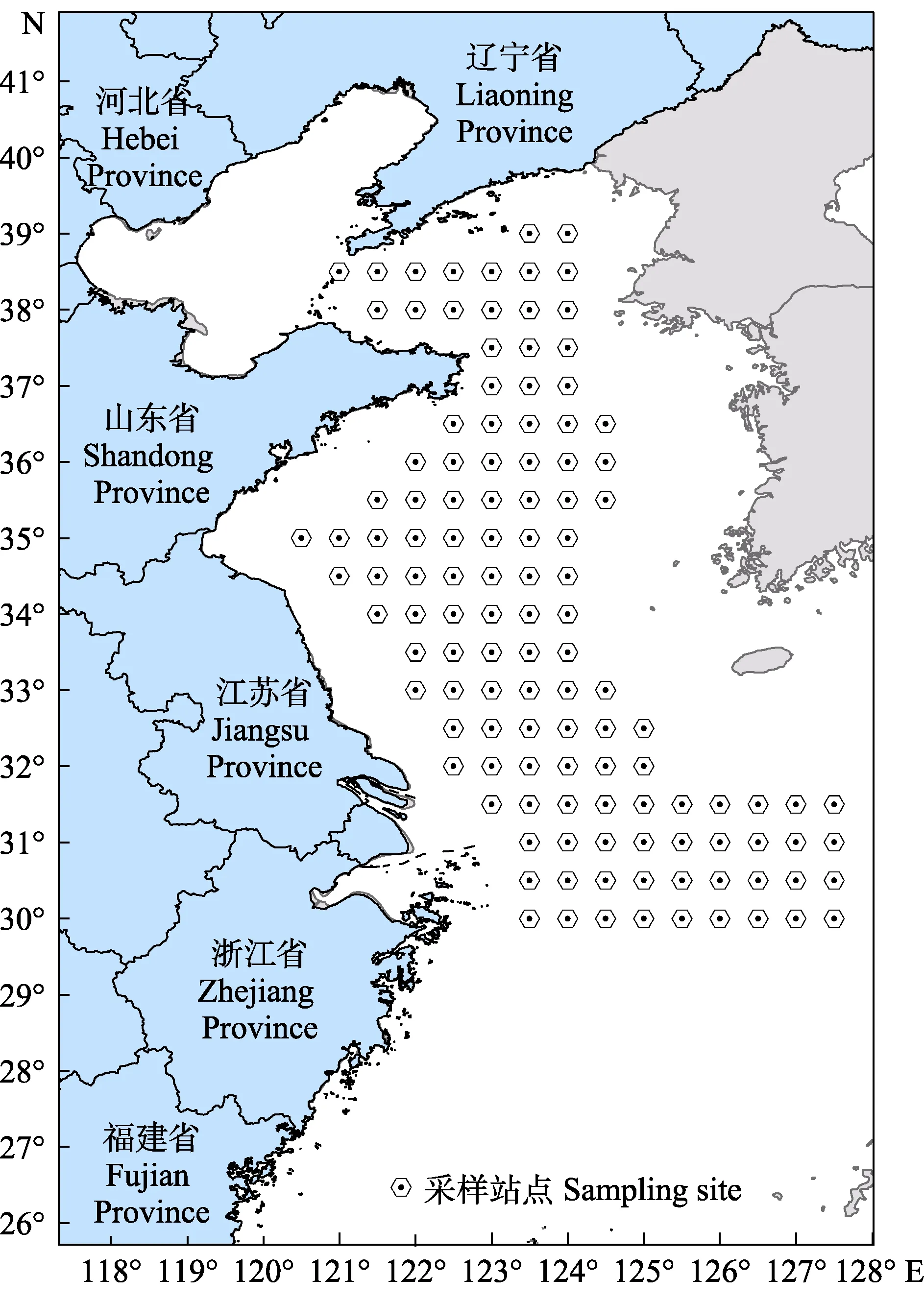
圖1 2016—2018年黃海和東海北部設置的采樣站點Fig.1Sampling sites in the Yellow Sea and the northern East China Sea from 2016 to 2018
1.2 蝦類樣品中WSSV 的環介導等溫擴增(LAMP)檢測
按照馬芳等(2016)報道的方法,使用WSSV現場快速高靈敏檢測試劑盒對涂抹于 FTA?卡片上的樣品進行LAMP檢測。為避免檢測過程中出現假陽性和假陰性,采用 2套不同引物制作的WSSV現場快速高靈敏檢測試劑盒對同一樣品進行重復檢測和復核。
1.3 海洋環境數據
各航次站點的底層溫度、底層鹽度、深度數據通過SBE25plus自容式CTD (SeaBird公司,美國)進行現場采集。底質類型(BST)參考石學法(2014)報道的方法進行分類,并考慮到各底質類型的站點數量,所有調查站點被概括為 4種類型:砂質(包括砂、砂質粉砂和中砂)、粉砂質砂、粉砂(包括粉砂和粉砂質黏土)和黏土質砂(包括黏土質粉砂和黏土質砂)。
1.4 模型的構建
1.4.1 模型的假設 以 WSSV 陽性蝦類總生物量作為模型的響應因子;考慮到蝦類總生物量數據呈正偏態分布(官文江,2015),以蝦類總生物量的對數(ln_Abu)作為響應變量。選取 WSSV陽性檢出率(P_rate)作為模型的解釋變量,并假設 WSSV陽性蝦類被WSSV感染的概率一致,即蝦類總陽性樣品數量=陽性檢出率×蝦類總生物量,并將陽性樣品量進行對數處理,得到的陽性樣品量的對數(ln_posi)作為模型的解釋變量。
1.4.2 模型影響因子的選取 除了P_rate和ln_posi以外,根據已發表的環境因子對蝦類生物量影響的研究報道(Gribble et al, 2007;黃慶洋等, 2009;逄志偉等, 2015;趙靜等, 2017),選取水深、底層溫度、鹽度、經緯度、底質類型和離岸距離等相關因子作為模型的解釋變量。
1.4.3 蝦類生物量影響因子的初步評價 采用梯度隨機森林模型篩選因子,并初步評價各環境因子對蝦類資源密度分布的影響。GFM 是一種機器學習方法,建立在隨機森林的基礎上,通過整合各響應變量上的單個隨機森林分析來捕獲潛在相關預測因子和多個響應變量之間的復雜關系(Ellis et al, 2012)。從本質上講,隨機森林是一種回歸樹,它將響應變量按每個預測因子p的特定分割值劃分為2組,以最大化同質性。與其他措施一起,梯度森林提供了每個響應變量f的擬合優度以及加權的重要性。對梯度森林進行1000次運行,獲得的變異度,并使用總體性能最高的模型(即擬合優度R2最大)進行進一步分析。
1.4.4 利用GAMM模型評價環境因子對蝦類分布的影響 考慮到解釋能力和預測能力的潛在差異,采用廣義加性混合模型對可能影響蝦類生物量的時空分布因子、環境因子和WSSV陽性檢出率等進行分析(Derville et al, 2018)。在建模之前,首先對數據進行初步分析、檢驗以選擇合適的模型,主要進行異常值的剔除以及檢查解釋變量之間的共線性。采用方差膨脹因子(VIF)度量多重共線性的程度,并對選取的初始因子進行多重共線性檢驗,篩選可納入模型的因子(Shono, 2008)。VIF越高,多重共線性的影響越嚴重,以VIF>5作為判斷是否存在多重共線性的標準。解釋變量中底質類型(BST)作為隨機效應。模型的連接函數為“identity”(Kabacoff, 2015)。利用F檢驗評估解釋變量的顯著性。將不同環境因子進行組合,生成多個候選GAMM模型。
采用赤池信息準則(Akaike information criterion,AIC)和方差解釋率(devianceexplained)來比較候選模型的性能(周慧敏等, 2014)。在分析中將不顯著變量(P≥0.1)從初始模型中逐一剔除,最終得到最優模型。主要步驟:1)剔除解釋偏差最小的不顯著變量;2)擬合簡化的 GAMM到數據;3)重復步驟 1和步驟 2,直到AIC值最小化(deLeeuw,1992;Burnham et al,2003)。數據分析和模型構建的過程中分別使用 R軟件中的“car”,“gradient Forest”和“mgcv”進行(R Core Team, 2017)。
2 結果
2.1 WSSV 在黃海與東海北部蝦類中的流行情況
在2016、2017和2018年8月對黃海和東海北部的調查中,從73個采樣點共采集517份蝦類樣品(圖 2,表 1)。LAMP檢測結果顯示,2016、2017和2018年所涉及的25、31和46個采樣站點中,WSSV陽性采樣站點的比例分別達48.40%、38.75%和21.74% (圖2、圖3),且3個年度樣品中WSSV陽性檢出率分別為16.86%、9.60%和4.80%(圖3);上述3個年度所采集樣品包括 26種蝦類,其中,脊腹褐蝦(Crangon affinis)、葛氏長臂蝦(Palaemon gravieri),哈氏仿對蝦(Parapenaeopsis harbwickii)、日本囊對蝦(Penaeus japonicus)、戴氏赤蝦(Metapenaeopsis dale)、中國毛蝦(Aceteschinensis)和海蜇蝦(Latreutes anoplonyx)等11種蝦類樣品中均有WSSV陽性檢出,除樣品量較少(少于10尾)的滑脊等腕蝦(Heterocarpoides laevicarina)、細螯蝦(Leptochela gracilis stimpson)和刀額仿對蝦(P.acultrirostris)外,中華安樂蝦(Eualus siensis)、海蜇蝦、葛氏長臂蝦和脊腹褐蝦中WSSV陽性檢出率皆在10%以上(表1)。
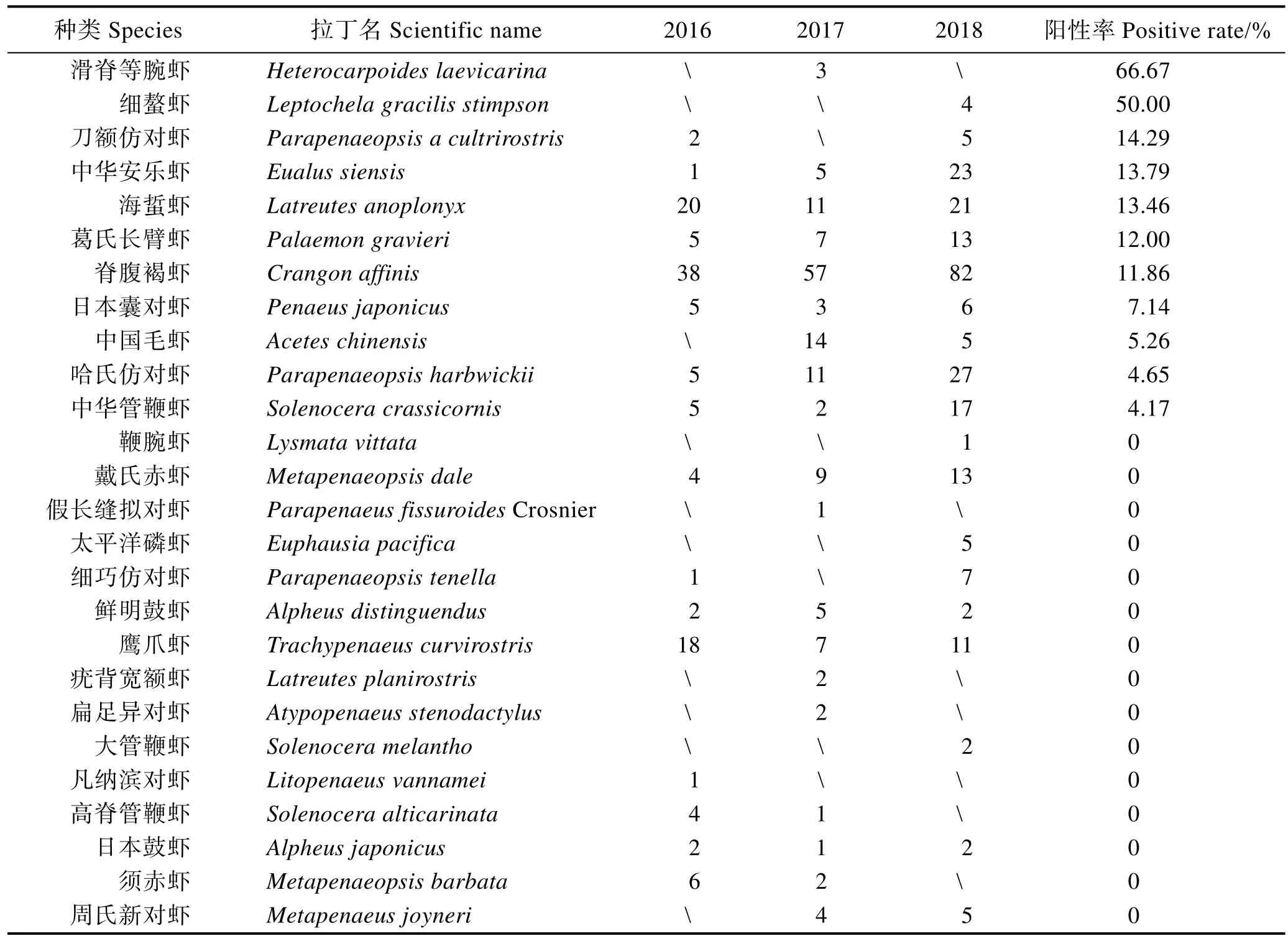
表1 2016—2018年在黃海和東海北部采集的不同蝦類樣品數量及樣品中WSSV的陽性檢出率Tab.1Number and WSSV positive rates in different shrimps collected from Yellow Sea and the northern East China Sea (2016—2018)
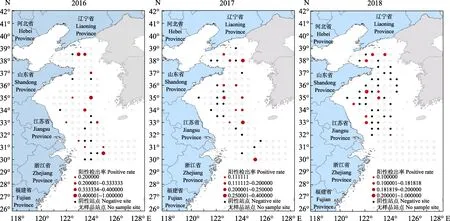
圖2 2016—2018年黃海和東海北部蝦類中WSSV的流行范圍和流行率Fig.2The prevalence scopes and prevalence rates of WSSV in the shrimps of the Yellow Sea and the northern East China Sea (2016—2018)
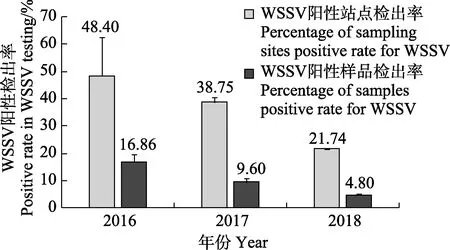
圖3 2016—2018年黃海和東海北部WSSV陽性采樣站點檢出率和陽性樣品檢出率Fig.3WSSV positive rate in the sampling sites and the collected samples of the Yellow Sea and the northern East China Sea (2016—2018)
2.2 GFM模型的結果分析
利用GFM模型對蝦類生物量相關影響因子的初步評價結果顯示,ln_posi(陽性樣品數的對數)重要性最高,其次是溫度(Temp)、離岸距離(Distance)、緯度(Lat)、經度(Long)、陽性率(P_rate)、鹽度(Sal)和底質類型(BST)(圖 4)。
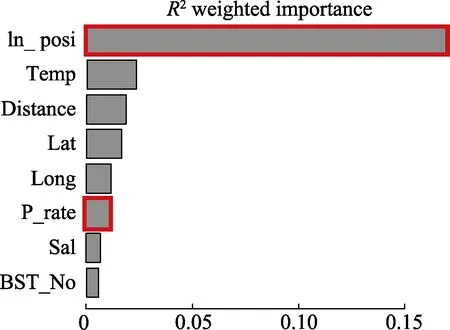
圖4 不同解釋變量在GFM模型中的重要性分析Fig.4Importance analysis of different predictor variables from the gradient forest
2.3 GAMM模型的結果分析
通過 VIF對初始影響因子進行多重共線性檢驗,發現初始影響因子之間存在共線性現象。將P_number和深度2個因子排除后,各因子的VIF值均小于5。因此,選取 ln_posi、Temp、Distance、Lat、Long、P_rate、Sal、BST_No作為構建GAMM 模型的備選影響因子(表2)。根據AIC最小原則,通過逐步剔除非重要因子,得到最優模型為Model 6:ln_Abu~P_rate+ln_posi+Long。P_rate和ln_posi是Model 6中的極顯著相關因子(P<0.001)。ln_Abu隨著P_rate的升高而降低,而ln_posi也會隨著ln_Abu的升高而升高(圖5)。
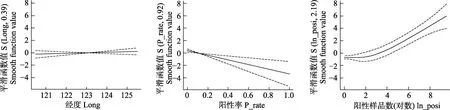
圖5 最優模型中顯著因子與蝦類生物量對數(ln_Abu)的關系Fig.5The relationship between significant variables and abundance of shrimps for the final generalized additive mixed models

表2 備選GAMM模型的擬合效果Tab.2Fitting effect of candidate GAMMs
3 討論
WSSV可引起養殖對蝦白斑綜合征,該病毒在1992年首次發現于我國臺灣(Chouet al, 1995),此后迅速蔓延到日本、韓國、東南亞各國以及南北美洲、歐洲等地,給全球對蝦養殖業造成了巨大的損失(Nakanoet al,1994; Inouyeet al,1994; Momoyamaet al,1994; Takahashiet al,1994; Wanget al,1995;Lightner,1999)。研究發現,在墨西哥、菲律賓和伊朗等國近岸海域捕獲的野生蝦類中存在WSSV感染(Chapmanet al, 2004; Mijangos-Alquisireset al, 2006;Oroscoet al, 2017; Gholamhosseiniet al, 2020),但WSSV是否也在遠離陸地海域的蝦類中流行尚未見報道。本研究對黃海和東海北部海域蝦類中WSSV流行情況的調查結果顯示,WSSV在我國上述海域的流行范圍比較廣、流行率也較高;特別是2016年,WSSV陽性采樣站點和陽性樣品的比率分別高達48.395%和16.86%,其中,樣品陽性率高于同年我國養殖對蝦中WSSV的陽性檢出率(12.9%)(農業部漁業漁政管理局等,2017)。在采集到的26種蝦類中,除已證實可被WSSV感染的日本囊對蝦和中國毛蝦(雷質文等,2003)外,還有9種蝦類樣品中有WSSV陽性檢出,這提示WSSV經由上述蝦類傳播擴散的風險不容忽視。2016、2017和2018年黃海和東海北部WSSV的陽性樣品檢出率和陽性站點檢出率均呈不斷下降的趨勢,推測可能與漁業主管部門和對蝦養殖從業者加大了苗種檢疫力度,海洋環境監測部門加強了養殖廢水排放監管,進而降低了WSSV由近岸養殖池塘向近海傳播有關。
本研究引入梯度隨機森林模型(GFM)和廣義加性混合模型(GAMM)評估了WSSV對調查海域蝦類生物量的潛在影響。對GFM模型中各解釋變量的分析顯示,ln_posi (WSSV陽性樣品數的對數)對響應變量蝦類生物量的對數(ln_Abu)的重要性最高。根據AIC最小原則篩選出的GAMM 最優模型顯示,ln_posi和 P_rate是蝦類生物量的極顯著影響因子,并且蝦類生物量隨WSSV陽性率的增高而下降。上述 GFM和 GAMM模型的分析結果均表明,WSSV流行對黃海和東海蝦類生物量存在潛在影響,野生蝦類感染WSSV后,可能會導致其生物量減少。黃東海是我國重要的漁業水域,甲殼類資源豐富,許多蝦類除了是重要的捕撈對象外,更是諸多頭足類、魚類和哺乳類等動物的主要捕食對象(韋晟等,1992;程濟生等, 1997)。如WSSV陽性檢出率較高的脊腹褐蝦,為黃東海的優勢種類,是眾多底層和近底層魚類的主要餌料生物(唐啟升,2006),其資源變動將會進一步影響主要經濟魚類的生物量和近海生態系統的穩定。
限于目前尚無WSSV對野生蝦類致病性的相關數據,本研究為便于模型分析,假設WSSV對呈現WSSV陽性蝦類的感染和危害風險相同,這一假設可能與實際情況存在偏差:有些蝦類對WSSV較為敏感,染病個體會出現病理變化甚至會死亡,而有些陽性檢出的種類或僅是WSSV的載體,并不一定會發病或死亡,僅在WSSV的水平傳播中起重要作用(Rajendran et al,1999)。因此,將來可通過進一步研究WSSV對近海蝦類致病性及其在群體中的傳播方式,為模型分析提供更多參數,以便提出更加合理的假設,進而提高模型的評估準確性和預測能力。
本研究的流行病學調查表明,黃海和東海北部蝦類優勢種——脊腹褐蝦、葛氏長臂蝦和日本囊對蝦等基本都攜帶了WSSV或被WSSV感染。為探討病毒傳播對近海蝦類生物量的影響,本研究把WSSV LAMP檢測呈陽性的蝦類總生物量作為模型響應因子,這有別于傳統的物種分布模型。傳統的物種分布模型一般評估海域溫度、鹽度和深度等環境因子對單一種群時空分布的影響,個別環境因子對種群分布的影響往往較大。而本研究模型的被解釋變量是蝦類生物量,包含蝦的種類較多,其環境適應性更廣,對單一蝦類存在影響的環境因子對蝦類生物量的解釋率不會被高估,這在客觀上更有利于分析WSSV傳播流行對蝦類生物量的影響。
從總體上看,觀測數據積累和基礎研究成果可為模型模擬或預測提供更多或更適合的因子與參數,并有利于提高響應變量對解釋變量的響應能力和預測能力。此外,構建模型時關注諸如殘差異質性、空間自相關、因子間交互作用等因素可有助于提高模型的預測能力。本研究僅是運用統計模型探索水產養殖動物病毒影響近海野生群體資源動態的初步嘗試,待完成更長時間、更大空間尺度上的調查數據積累,獲得養殖蝦類病毒對野生蝦類致病特性等信息,并進一步優化模型因子后,將會更準確地解釋和預測養殖動物病原傳播對近海生物種群、生態系統影響的規律和趨勢。
致謝:中國水產科學研究院黃海水產研究所“北斗號”漁業科考船上科研人員與工作人員在本研究的調查和樣品采集過程中給予了大力幫助,謹致謝忱!
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
