小麥果糖激酶基因TaFRK與TaPFT的互作分析
2022-01-06 03:07:18張露凡宋普文毋柳柳胡海燕李成偉
麥類作物學報 2021年11期
張露凡,宋普文,毋柳柳,胡海燕,李成偉
(河南科技學院生命科技學院/河南省植物遺傳改良與土壤修復國際聯合實驗室,河南新鄉 453000)
小麥(TriticumaestivumL.)是世界最重要的糧食之一,也是我國重要的糧食作物,目前我國耕地面積逐漸減少,保證糧食作物高產穩產和高質量發展就顯得尤為重要。同時我國也是小麥的生產大國和消費大國,小麥赤霉病的流行已經嚴重威脅到我國的糧食和食品生產安全[1]。
小麥赤霉病是由鐮刀菌屬真菌引起的一種世界性病害,流行年份會導致小麥產量下降,甚至導致絕收[2]。同時鐮刀菌還會在小麥體內產生多種真菌毒素,如脫氧雪腐鐮刀菌烯醇(DON)、玉米赤霉烯酮(ZEN)等,人畜誤食會造成嚴重危害[3]。
減輕小麥赤霉病危害最經濟、環保的手段就是培育抗病品種[4]。但是國內外抗源較少,農藝性狀較差和抗性機理不清楚阻礙了抗病品種的培育[5]。因此,挖掘小麥赤霉病抗性基因資源、創制抗赤霉病新種質以及研究小麥抗赤霉病抗性機理仍是目前小麥赤霉病的研究方向。Fhb1是在蘇麥3號中發現的抗赤霉病效應和穩定性最好的位點,隨后在望水白、黃方柱和日本品種Nyubai中均發現了該位點[6]。Rawat等[7]通過對蘇麥3號3BS染色體上主效位點Fhb1的精細定位和圖位克隆,獲得了抗赤霉病擴展的基因PFT(pore-forming toxin-like)。2019年Su等[8]在Fhb1位點鑒定到一個編碼組氨酸富集的鈣離子結合蛋白基因TaHRC,被認為是Fhb1位點的主要抗性基因,并發現TaHRC基因沉默后小麥抗性增強,而Li等[9]將TaHRC基因過表達后發現小麥抗性增強,所以,其作用機制目前也存在分歧,同時也說明了小麥赤霉病抗性機制可能不是由一個基因控制的,可能存在其他基因的干預。He等[10]通過以PFT為誘餌篩選小麥酵母雙雜文庫,得到23種與PFT互作的蛋白,并通過實時定量PCR分析這些互作蛋白基因的表達模式,發現5種蛋白在植物生長和發育的各個過程中以及在抵抗生物和非生物脅迫方面發揮關鍵作用。
本研究先以PFT為誘餌蛋白篩選小麥酵母雙雜文庫,得到一個新的基因,然后通過酵母雙雜交試驗、雙分子熒光互補試驗和實時熒光定量分析,對其表達模式以及與小麥TaPFT的相互作用進行分析,以期為解析小麥赤霉病抗性機制奠定基礎。
1 材料與方法
1.1 材 料
抗赤霉病小麥蘇麥3號和本氏煙草均由河南科技學院河南省糧食作物基因組編輯工程技術研究中心提供。大腸桿菌DH5α感受態細胞、根癌農桿菌GV3101感受態細胞、酵母Y2HGold菌株均由本實驗室保存。
1.2 方法
1.2.1 小麥RNA的提取與cDNA的合成
在小麥生長至兩葉一心時,取一整株小麥用ddH2O沖洗干凈后迅速放入液氮冷凍,-80 ℃保存。采用Trizol法提取小麥RNA,使用RNase-Free DNase I試劑盒去除混雜的基因組DNA,用超微量分光光度計測量RNA樣品的純度和濃度。cDNA的合成按照PrimeScriptTMII 1st Strand cDNA Synthesis Kit試劑盒(寶生物,大連)說明書進行,反轉錄產物于-20 ℃保存。
1.2.2 PFT互作蛋白基因的酵母篩庫
將TaPFT基因連接至酵母表達載體pGBKT7上,通過酵母轉化的方法導入酵母菌株Y2HGold中,涂布于SD-Trp平板培養基上, 30 ℃培養長出單克隆后,挑取單克隆接種至50 mL SD-Trp液體培養基中培養12~16 h,作為原代培菌液。利用酵母轉化試劑盒(TaKaRa,大連)制備酵母感受態細胞,將前期制備好的小麥酵母雙雜cDNA文庫轉化至酵母感受態中,并涂布于SD/-His-Leu-Trp-Ade平板培養基上,于 30 ℃培養3~7 d后,挑取單克隆進行擴繁培養,使用酵母質粒提取試劑盒(天根,北京)提取酵母質粒,并以酵母質粒為模板進行PCR擴增,擴增產物用膠回收試劑盒(天根,北京)進行回收后,送至武漢金開瑞生物工程有限公司進行測序。用ORF Finder分析表明,其中一個文庫插入序列具有771 bp的完整編碼框,進一步BlAST分析表明,其序列與已知小麥cDNA文庫克隆Triticum aestivum clone wyr1c.pk002.o12(BT008921.2)序列具有99.48%的相似性,其編碼蛋白與野生二粒小麥(Triticumdicoccoides)果糖激酶Fructokinase-2蛋白序列具有99.48%的相似性,因此推測該基因編碼蛋白為Fructokinase,命名為TaFRK。
1.2.3 基因克隆
使用Primer 6.0軟件設計引物TaFRK-F/R和 TaPFT-F/R(表1),擴增TaFRK和TaPFT基因的全長序列。以TaFRK和TaPFT的cDNA為模板擴增TaFRK(771 bp)和TaPFT(1 434 bp)的編碼區序列。PCR反應體系為25 μL,包括模板cDNA 1 μL,上下游引物(10 μmol·L-1)各1 μL,Taq酶12.5 μL,ddH2O 9.5 μL。PCR反應程序:預變性94 ℃ 5 min,變性94 ℃ 30 s,退火55 ℃(TaFRK:45 s;TaPFT:90 s),延伸72 ℃ 30 s,35個循環;延伸72 ℃ 7 mim,4 ℃保存。用1%瓊脂糖凝膠電泳檢測PCR產物。
PCR產物膠回收后分別與pMD19-T載體于16 ℃連接過夜,連接產物轉化至大腸桿菌DH5α感受態細胞后,涂抹于含有Amp+抗性的培養基上,隨機選取5個單克隆菌落,菌落PCR鑒定為陽性后,送至武漢金開瑞生物工程有限公司進行測序。
1.2.4 酵母雙雜交分析
使用Primer 6.0軟件設計的引物pGADT7-TaFRK-F/R和pGBKT7-TaPFT-F/R(表1)分別擴增TaFRK和TaPFT基因,對擴增產物進行電泳并進行回收后,將回收產物TaFRK連接至經過EcoR I和XhoI雙酶切的pGADT7載體上,構建pGADT7-TaFRK重組載體;將回收產物TaPFT連接至經BamH I和EcoR I酶切的pGBKT7載體上,構建pGBKT7-TaPFT重組載體,然后將這兩個重組載體分別轉化至大腸桿菌DH5α感受態細胞。對單克隆進行PCR驗證,將陽性克隆送至武漢金開瑞生物工程有限公司進行測序。用通用引物T7和3AD對pGADT7-TaFRK進行測序,用通用引物T7和3BD對pGBKT7-TaPFT進行測序,用DNAMAN軟件對測序結果進行對比分析。

表1 本試驗使用的引物Table 1 Primers used in the study
使用PEG/LiAc法轉化酵母菌株Y2HGold,以pGBKT7-TaPFT和pGADT7-TaFRK共轉化作為試驗組,以pGBKT7和pGADT7空載體共轉化作為對照組,pGBKT7-TaPFT和pGADT7、pGADT7-TaFRK和pGBKT7共轉化作為陰性對照。轉化后的酵母涂抹在含有X-α-Gal和AbA的 SD/-Leu/-Trp/-His/-Ade平板培養基上,于30 ℃靜置培養3~ 5 d,拍照并記錄。
1.2.5 雙分子熒光互補分析
使用Primer 6.0軟件設計的引物BiFC-TaFRK-F/R和BiFC-TaPFT-F/R(表1)分別擴增TaFRK和TaPFT基因的開放閱讀框。對擴增產物進行電泳并回收。使用Gateway體系構建雙分子熒光互補載體,經BP反應將回收產物TaFRK和TaPFT分別連接至pDNOR221中間載體,構建pDNOR221-TaFRK和pDNOR221-TaPFT重組載體,然后轉化至大腸桿菌DH5α感受態細胞。對PCR鑒定為陽性的單克隆送至武漢金開瑞生物工程有限公司進行測序。將測序正確且含有pDNOR221-TaFRK和pDNOR221-TaPFT的單克隆分別進行搖菌提質粒后,經過MluI快切酶酶切,用回收試劑盒(天根,北京)進行回收,將目的基因經LR反應連接至YC和YN載體中,構建YC-TaFRK和YN-TaPFT載體,然后轉化至大腸桿菌DH5α感受態細胞,對PCR鑒定為陽性的單克隆送至武漢金開瑞生物工程有限公司進行測序。用DNAMAN軟件對測序結果進行對比分析。
將YC-TaFRK和YN-TaPFT載體分別轉入農桿菌GV3101中進行搖培,待OD600約為0.6~0.8時,5 000 g離心1 min富集菌體,用無菌水洗滌菌體,再用煙草侵染緩沖液(D-glucose 250 mg,0.5 mol·L-1的MES 5 mL, 1 mol·L-1的乙酰丁香酮5 μL,0.02 mol·L-1的Na3PO4·12H2O 5 mL,加無菌水至50 mL)重懸菌體后將兩種菌液等比例混合,黑暗靜置2 h,然后使用注射器將菌液注射至6~8周齡的煙草葉片;同時設置對照將空載體YC和YN注射至煙草,用記號筆標記注射區域,25 ℃培養48 h后,在激光共聚焦顯微鏡下觀察結果,并拍照。
1.2.6 接種禾谷鐮刀菌后TaFRK基因的表達模式分析
按照Bai等[11]的方法用綠豆湯培養基培養禾谷鐮刀菌,產生孢子后于7 000 g 離心15 min收集孢子,用無菌水沖洗2~3遍。于小麥開花期,使用移液器將15 μL禾谷鐮刀菌孢子懸浮液(200個孢子·μL-1)注入穗狀花序的中央小花中,接種過的穗包裹在夾層袋中,夾層袋內部用水霧化,分別在小麥接種禾谷鐮刀菌0、 3、6、12、24、36、48和72 h時取樣,對照組用無菌水接種小麥小穗。每個時間點收集2個小穗,取下后置于液氮中冷凍, -80 ℃保存。

為了標準化每個反應中的cDNA總量,并消除樣品之間的差異,以小麥微管蛋白基因Tubulin作為內部參照。每個樣品重復3次,使用2-ΔΔCT方法計算TaFRK基因的相對表達量,進行3次生物學重復。
2 結果與分析
2.1 酵母雙雜交載體和雙分子熒光互補載體的構建
用引物TaPFT-F/R和TaFRK-F/R擴增TaPFT和TaFRK基因的全長片段,擴增結果如圖1A和圖1B所示。對擴增片段進行回收,與pMD19-T載體16 ℃連接過夜后轉化大腸桿菌DH5α感受態細胞,挑5個單克隆搖菌,菌液PCR驗證為陽性后進行測序。將測序正確的菌液提取質粒后分別連接在pGBKT7(BamH I和EcoR I雙酶切)和pGADT7(EcoR I和XhoI雙酶切)載體上。經PCR檢測后送測序,分別得到約1 434和771 bp的片段(圖1C和圖1D),與TaPFT和TaFRK的片段大小一致,表明表達載體構建 成功。
雙分子熒光互補載體的構建使用Gateway體系,分別使用引物BiFC-TaPFT-F/R和BiFC-TaFRK-F/R分別擴增TaPFT和TaFRK基因的開放閱讀框,經BP反應將TaPFT和TaFRK基因連接至中間載體pDNOR221,菌落PCR驗證為陽性后送測序。將測序正確的載體用MluI酶切后回收,然后經LR反應將TaPFT和TaFRK分別與YN和YC載體連接,并進行測序,分別得到大小分別為1 434和771 bp的片段(圖1E和1F),與TaPFT和TaFRK的片段大小一致,表明表達載體構建成功。
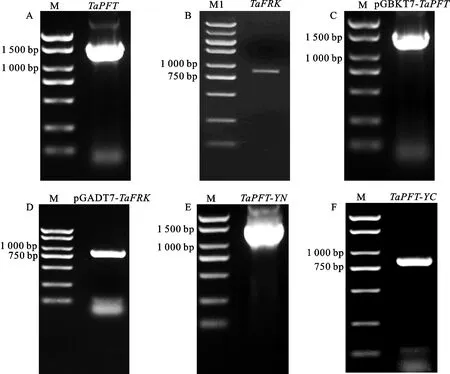
M:Marker DL2000;M1:Marker DL5000.圖1 TaPFT和 TaFRK基因的克隆及酵母雙雜交載體和雙分子熒光互補載體的構建Fig.1 Cloning of TaPFT and TaFRK genes and construction of yeast two hybrid vector and bimolecular fluorescence complementary vector of TaPFT and TaFRK
2.2 酵母雙雜交驗證 TaFRK和 TaPFT的相互 作用
酵母雙雜交是鑒定蛋白相互作用最常用的手段。將共轉pGBKT7-TaPFT和pGADT7-TaFRK、pGBKT7-TaPFT和pGADT7、pGBKT7和pGADT7-TaFRK、pGBKT7和pGADT7的酵母菌菌株Y2HCold在含有X-α-Gal和AbA的SD/-His-Leu-Trp-Ade的平板培養基上進行劃線培養,發現只有共轉pGBKT7-TaPFT和pGADT7-TaFRK的酵母變藍(圖2)。表明TaPFT和TaFRK基因可以在酵母中發生相互作用。
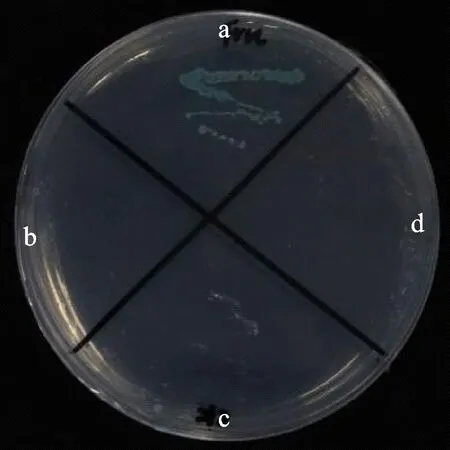
a:pGBKT7-TaPFT載體與pGADT7-TaFRK載體共轉酵母涂板;b:pGBKT7-TaPFT載體與pGADT7載體共轉酵母涂板;c:pGBKT7載體與pGADT7-TaFRK載體共轉酵母涂板;d:pGBKT7載體與pGADT7載體共轉酵母涂板。a: pGBKT7-TaPFT vector and pGADT7-TaFRK vector were co-transferred into yeast; b: pGBKT7-TaPFT vector and pGADT7 vector were co-transferred into yeast; c: pGBKT7 vector and pGADT7-TaFRK vector were co-transferred into yeast; d: pGBKT7 vector and pGADT7 vector were co-transferred into yeast.圖2 酵母雙雜交檢測 TaFRK和 TaPFT的相互作用Fig.2 Interaction between TaPFT and TaFRK detected by yeast two-hybrid system
2.3 雙分子熒光互補試驗驗證 TaFRK和 TaPFT的相互作用
為了進一步驗證TaFRK和TaPFT是否可以在植物體內發生相互作用,利用雙分子熒光互補試驗探討TaFRK和TaPFT在煙草細胞中的相互作用情況。結果(圖3)發現,攜帶YC-TaFRK和YN-TaPFT農桿菌組合的煙草表皮細胞中可以觀察到黃色熒光,且熒光主要分布在細胞質中,表明TaFRK和TaPFT在細胞質中發生了相互作用,而只攜帶YC和YN農桿菌組合的煙草表皮細胞中未觀察到黃色熒光。
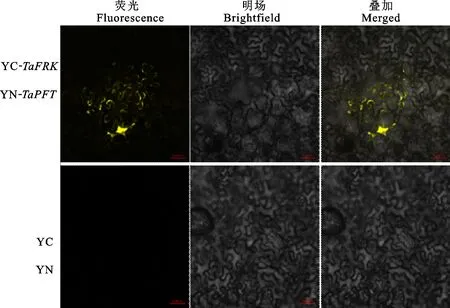
圖3 雙分子熒光互補實驗檢測 TaPFT和 TaFRK相互作用Fig.3 Interaction between TaPFT and TaFRK detected by Bi-Fluorescent Complementation
2.4 接種禾谷鐮刀菌后 TaFRK的表達模式
由表2可以看出,隨著時間的延長,接種禾谷鐮刀菌孢子處理組TaFRK基因的相對表達量總體上呈上升趨勢(除接種12 h時有所下降);而對照組TaFRK基因的相對表達量呈先上升后下降的趨勢。在接種6~72 h(除接種12 h),處理組TaFRK基因的相對表達量均顯著高于對照組。表明禾谷鐮刀菌孢子侵染后誘導了TaFRK基因的上調表達。說明TaFRK基因可能參與小麥赤霉病抗性反應。

表2 接種禾谷鐮刀菌后小麥小穗中TaFRK的相對表達量Table 2 Relative expression of TaFRK gene in wheat spikelets after anoculation with Fusarium graminearum spores
3 討 論
果糖激酶(Frlctokinase,FRK)是己糖激酶的一種,不僅參與糖的代謝,還可作為糖感受器和信號分子來調控植物的生長發育[12-14],激活或阻遏植物體內相關基因的表達來影響細胞周期、光合作用、碳氫代謝、逆境響應、萌發、營養生長、生殖發育、衰老等過程[15]。Granot等[16]通過嫁接試驗表明,反義抑制番茄LeFRK2可引起葉片枯萎以及莖組織中活性木質部面積明顯變小。說明LeFRK2參與木質部的發育過程,且木質部發育可能與果糖有關。果糖是蔗糖的水解產物,蔗糖合成酶(SUSY)可將蔗糖裂解成UDP葡萄糖和果糖,其中UDP葡萄糖已經證明參與番茄的維管系統表達[17-18],這說明SUSY也可能與木質素發育有關。
Roach等[19]研究表明,在楊木發育過程中RNA干擾FRK2基因的表達,可增加可溶性糖的含量以及降低己糖磷酸和UDP-葡萄糖的含量,表明FRK2基因與胞壁多糖前體的碳流有關。
FRK2蛋白的活性降低也導致纖維細胞壁變薄,纖維素比例降低[19]。這一結果確立了FRK2在木材纖維素碳通量中的中心作用。
Tomassini等[20]研究表明,禾谷鐮刀菌在入侵小麥時在體外產生纖維素酶、木聚糖酶等降解宿主細胞壁的酶來降解細胞壁,從而穿透和定殖宿主組織。而木質素在禾本科植物細胞壁占比20%,是一種重要的結構成分,可防御病原體的入侵,使細胞壁對細胞壁降解酶更具抵抗力,同時阻止病原體產生的毒素擴散。Lionetti等[21]研究表明,較高的S型木質素含量可能是一種與抗病性有關的細胞壁生化特性,同時也表明有利于S型木質素積累的基因可能參與了對病原菌的抗性。
Yang等[22]研究發現,蘋果MdFRK2的轉錄受到褪黑素(N-乙酰基-5-甲氧基色胺)的抑制,較高的褪黑素濃度會抑制MdFRK2的表達和降低FRK蛋白的活性。褪黑素是植物中廣泛存在的具有高度保守性的多功能小分子,當由于生物脅迫導致細胞滲漏時,可通過激活防御相關基因的表達參與植物的光周期反應、清除活性氧以及保護植物免受細菌病原體的侵害[23-24]。說明MdFRK2受褪黑素的調控,進而參與植物的抗病 過程。
本研究使用酵母雙雜交和雙分子熒光互補試驗證明,TaFRK和抗病基因PFT能夠發生互作反應,實時熒光定量研究表明,TaFRK基因表達受赤霉病侵染誘導。而FRK基因在植物細胞壁和木質素合成中起重要作用[20-21],我們推測TaPFT可能通過與TaFRK互作影響細胞壁或木質素的積累來提高植物細胞壁的抗病特性。本研究為解讀小麥抗赤霉病抗性機理具有一定的參考意義,為解析TaPFT抗性機制提供新途徑。
