亞氣生球狀綠藻一新種
——重慶單星藻
2021-12-24 05:19:02王清華宋會銀劉旭東胡征宇劉國祥
水生生物學報 2021年6期
關鍵詞:重慶
王清華 宋會銀 劉旭東 胡征宇 劉國祥
(1. 太原師范學院生物系, 晉中 030619; 2. 中國科學院海洋研究所海洋生態(tài)與環(huán)境科學重點實驗室, 青島 266071; 3. 山西大學生命科學學院, 太原 030006; 4. 中國科學院水生生物研究所中國科學院藻類生物學重點實驗室, 武漢 430072)
單星藻屬CoelastrellaChodat隸屬于綠藻綱Chlorophyceae環(huán)藻目Sphaeropleales柵藻科Scenedesmaceae空星藻亞科Coelastroideae, 屬內植物多為單細胞或聚合群體, 常附生于干燥或潮濕的石塊、墻壁或樹皮表面等氣生和亞氣生環(huán)境, 有時也可見于淡水水體。單星藻屬主要的形態(tài)特征是細胞呈球形、橢球型或呈紡錘形, 細胞壁具一端或兩端頂點增厚, 多數細胞壁表面具規(guī)則的縱向分布的肋紋, 但部分種類肋紋僅在掃描電鏡下可見, 而在光鏡下并不明顯。
單星藻屬植物具有共性特征, 即在自然生長后期或是在高鹽、高光等脅迫條件下, 藻細胞極易積累類胡蘿卜素和油脂而變?yōu)槌赛S色至磚紅色外觀,主要積累的類胡蘿卜素有角黃素、蝦青素、葉黃素、紫黃素和金盞花黃質等[1,2]。近年來, 大量新發(fā)現的單星藻藻株被應用于生理學研究。有研究表明, 一株分離于中國臺灣南部熱帶地區(qū)能耐受高溫環(huán)境的單星藻藻株Coelastrellasp. F50, 其積累類胡蘿素的含量可占細胞干重的4%以上[3]。藻株Coelastrellasp. M60在3%的NaCl鹽脅迫條件下, 細胞積累油脂含量可占細胞干重的37%以上[4]。除了在天然抗氧化劑、食品色素和生物油脂等生物質生產方面的優(yōu)勢, 該類群在生物水環(huán)境監(jiān)測、水體營養(yǎng)鹽去除、金納米粒子生物合成和工業(yè)廢氣再利用等方面研究均有良好表現[5—8]。然而, 目前關于該類群的基礎分類學研究卻相對薄弱, 致使許多新分離的有經濟價值的藻株無法得到準確的物種鑒定和命名。藻株命名的混淆與雜亂, 也使得類似的生理學研究間缺乏可比性, 阻礙了這些優(yōu)質藻種資源的進一步開發(fā)利用。
單星藻屬最初由Chodat建于1922年, 以肋紋單星藻Coelastrella striolata為模式種[9]。其后, 經過多次分類學變更, 目前被置于柵藻科空星藻亞科[10]。屬內成員主要包括原單星藻屬OriginalCoelastrella和原擬螺翼藻屬OriginalScotiellopsis成員。擬螺翼藻屬Scotiellopsis最初由Vinatzer建于1975年,以紅色擬螺翼藻Scotiellopsis rubescens為模式種[11]。1981年Pun?ochá?ová和Kalina對該屬進行修訂, 將許多螺翼藻屬(ScotiellaFritsch)中以似親孢子繁殖的個體轉移至此屬。他們的研究最先闡明單星藻屬與擬螺翼藻屬可能具有較近的親緣關系, 兩個屬間最主要的形態(tài)區(qū)別僅僅是細胞形狀不同(單星藻屬呈球形或寬橢球型, 擬螺翼藻屬多呈紡錘形)及在單星藻屬中不存在頂點增厚[12]。1998年, Hanagata首次基于18S rDNA分子數據對擬螺翼藻亞科Scotiellocystoideae進行系統(tǒng)發(fā)育分析, 發(fā)現單星藻屬與擬螺翼藻屬在系統(tǒng)發(fā)生上與柵藻類群較近, 至此二者由小球藻科轉移至柵藻科[13]。關于二者間的關系一直以來都有爭論, 依據傳統(tǒng)的形態(tài)學分類方法,普遍認為二者是不同的屬[14]。但是2013年, 捷克藻類學家Kaufnerová和Eliá?對該類群18S rDNA及ITS2二級結構特征進行分析, 證明了擬螺翼藻屬的模式種紅色擬螺翼藻與單星藻類群在系統(tǒng)發(fā)生上無法區(qū)分, 二者合并后, 目前認為擬螺翼藻屬是單星藻屬的異名[15]。
我國對單星藻屬的關注較晚, 宋會銀[16]最先在2015年報道在我國分離到單星藻藻株, 發(fā)現2個中國新記錄種(塞班單星藻Coelastrella saipanensis和紅色單星藻Coelastrella rubescens)。其后作者對我國的單星藻類群做過初步調查, 共發(fā)現3個新種(耐熱單星藻Coelastrella thermophila、英山單星藻Coelastrella yingshanensis及薄壁單星藻Coelastrella tenuitheca), 2個新變種(紅色單星藻卵形變種Coelastrella rubescensvar.oocystiformis和耐熱單星藻球形變種Coelastrella thermophilavar.globulina),另有1個中國新記錄種(卵形單星藻Coelastrella oocystiformis)[17]。此外, 作者也發(fā)表了塞班單星藻的葉綠體基因組數據[18]。本研究于我國重慶縉云山和河南周口地區(qū)采集到2株單星藻藻株, 經形態(tài)學和系統(tǒng)發(fā)育學鑒定為一新種, 命名為重慶單星藻Coelastrella chongqingensissp. nov.。
1 材料與方法
1.1 樣本的采集與培養(yǎng)
本研究所用藻株分別采集于河南周口及重慶縉云山。河南周口藻株FACHB-3314以無菌BG11平板置于空氣下12h后, 帶回實驗室培養(yǎng)分離獲得。重慶縉云山藻株FACHB-3315分離于陰暗區(qū)潮濕石頭表面, 以刀片直接刮取樣品, 放于離心管后帶回實驗室經平板涂布法分離獲得。分純后的藻株置于25℃恒溫培養(yǎng)箱中培養(yǎng), 光照強度為 30 μmol/(m2·s)左右。
1.2 藻株形態(tài)特征觀察
本研究光學顯微圖片主要采集于Olympus BX53(Olympus Cop, 日本)光學顯微鏡, 分別用明場(Bringt field, BF)、微分干涉(Differentical interference contrast, DIC)模式和熒光顯微技術(Epifluorescence microscopy, EFM), 觀察藻株的細胞形態(tài),用印度墨水負染, 確認藻株是否具膠被。
掃描電鏡制樣選取對數生長期的細胞, 先用2.5%戊二醛固定, 再經乙醇梯度脫水后用臨界點干燥儀干燥樣品, 后經細胞噴金, 用Hitachi S-4800掃描電鏡(Hitachi High-Technologies, 日本)觀察[18]。
透射電子顯微鏡制樣選取對數生長期細胞, 分別經2.5%戊二醛及1%鋨酸固定后, 再用1%的醋酸鈾過夜處理樣品。經過一系列濃度的乙醇梯度脫水, 在包埋劑內包埋, 固化后用Leica uc7超薄切片機(Wetzlar, 德國)切片, 厚度為70 nm。用2% 醋酸鈾-枸櫞酸鉛雙染色, 最后用透射電鏡(Hitachi High-Technologies, 日本)觀察[19]。
1.3 基因組DNA獲取及單分子標記擴增
取培養(yǎng)至對數生長期的藻株于2 mL凍存管中,用磷酸鹽緩沖液(PBS pH=7.0)洗滌3次, 加入0.4 mL的磷酸鹽緩沖液和適量的玻璃珠(d=0.5 mm), 用MiniBeadBeater(Model607EUR, 美國)震蕩3min(分3次, 每次1min)以破碎細胞。之后, 使用植物基因組小量制備試劑盒(AxyPrep, 蘇州, 中國)提取基因組DNA。
18S rDNA擴增采用引物對18A: 5′-AACCTG GTTGATCCTGCCAGT-3′; 18B: 5′-TGATCCTTCT GCAGGTTCACCTAC-3′[20]。ITS序列擴增采用引物對NS7: 5′-GGCAATAACAGGTCTGT-3′[21];LR1850: 5′-CCTCACGGTACTTGTTC-3′。tufA基因擴增采用引物對tufAF: 5′-TGAAACAGAAMAW CGTCATTATGC-3′;tufAR: 5′-CCTTCNCGAAT MGCRAAWCGC-3′[22]。PCR產物經瓊脂糖凝膠電泳檢測后送測序公司測序(擎科生物, 武漢)。測序結果使用ContigExpress Project 進行拼接[23], 并手動校正, 經校正后的序列提交至GenBank數據庫。
1.4 系統(tǒng)發(fā)育學分析
從GenBank數據庫中獲得柵藻類群近緣序列,18S rDNA和ITS序列登錄號及藻株信息示于表 1。序列比對使用軟件Mafft7.0[24], 比對后的數據集使用MEGA 6.0[25]手工裁剪校正。序列拼接串聯(lián)使用軟件SequenceMatrix[26], 使用MEGA 6.0和Bioedit 7.0[27]進行堿基組成分析、堿基轉換與顛換的比率分析等。序列飽和度檢驗利用軟件DAMBE[28]。系統(tǒng)發(fā)育分析主要采用最大似然法(Maximum likelihood, ML)和貝葉斯算法(Bayesian Inference, BI)。ML法使用IQ-TREE web server[29]完成。BI法則選擇使用MrBayes 3.2.1完成[30]。貝葉斯分析運行3000000代, 每1000代取樣一次, 分析達到平穩(wěn)(標準誤差<0.01)則運行結束。

表1 下載自GenBank數據庫中的近緣藻株序列(18S rDNA和 ITS)信息Tab. 1 Strains whose sequences (18S rDNA & ITS) were downloaded from the GenBank

續(xù)表1
2 結果
2.1 形態(tài)學描述及新種確認
營養(yǎng)細胞多為單細胞, 幼期細胞多呈橢球型,少數卵形或不規(guī)則球形, 細胞大小約(6—9) μm×(5—7) μm, 一個周生片狀色素體(圖 1A和1B)。成熟細胞多呈球形或橢球型, 直徑約8—14—(18) μm(圖 1C)。色素體周生, 具一個明顯蛋白核, 并隨細胞成熟逐漸碎片化。細胞通常具一至多個液泡。多通過形成2—4—8—16個似親孢子進行無性生殖(圖 1D—F), 有性生殖未觀察到。老化細胞的細胞壁增厚, 細胞內逐漸積累類胡蘿卜素而變?yōu)榇u紅色(圖 1G、1H), 印度墨水染色后顯示細胞外無膠被(圖 2I)。掃描電鏡照片顯示該種細胞壁近似光滑或僅具細微皺紋(圖 2A和2B), 不具有單星藻屬典型的縱向分布的肋紋。透射電鏡顯示周生色素體具一個明顯蛋白核(圖 3A、3C—E)及若干淀粉粒(圖 3B),蛋白核外裹有2個厚淀粉鞘(圖 3A、3C—E)。老化細胞的細胞壁增厚, 并富集大量溶有類胡蘿卜素的油滴(圖 3F)。
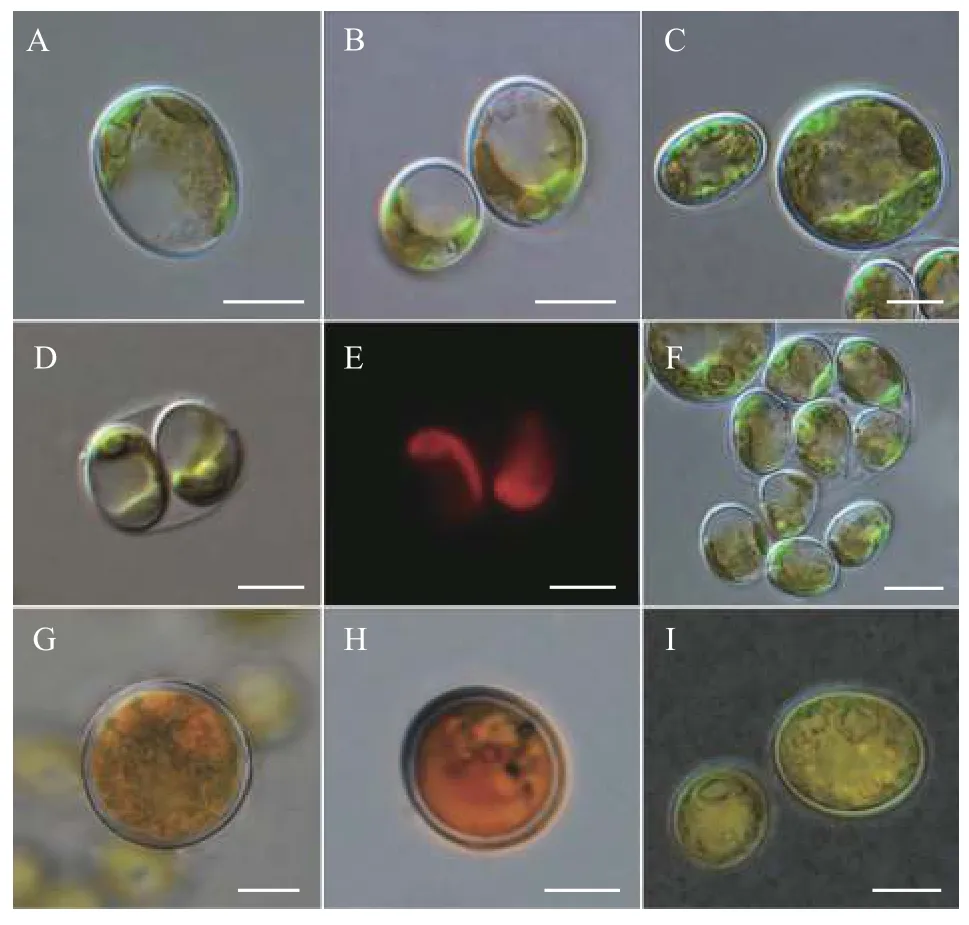
圖1 重慶單星藻普通光學顯微照片(標尺=5 μm)Fig. 1 Light micrographs of Coelastrella chongqingensis sp. nov.(Scale bar=5 μm)
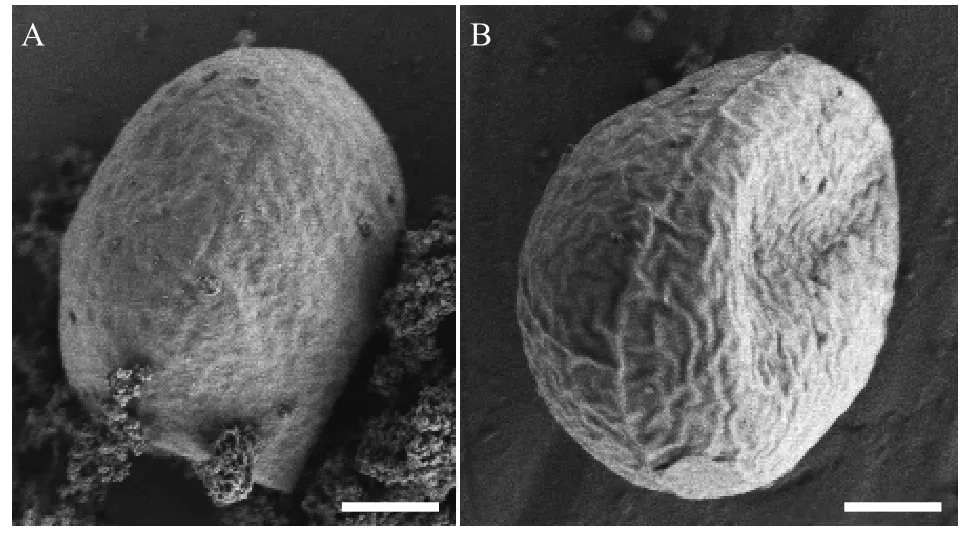
圖2 重慶單星藻掃描電鏡照片(標尺=2 μm)Fig. 2 The SEM images of Coelastrella chongqingensis sp. nov.(Scale bar=2 μm)

圖3 重慶單星藻透射電鏡照片(標尺=2 μm)Fig. 3 The TEM images of Coelastrella chongqingensis sp. nov.(Scale bar=2 μm)
Coelastrella chongqingensis Wang Q H, Song H Y& Liu G X sp. nov. Fig. 1—3.
Description.Vegetative cells are solitary, with approximately smooth cell walls or only slightly wrinkled. Young cells are mostly ellipsoidal or oval, a few near-spherical, blunt end and a parietal chloroplast,measure (6—9) μm×(5—7) μm. Mature cells are mostly spherical or ellipsoidal, 8—14—(18) μm. The chloroplast is parietal with a distinct pyrenoid (coated with 2 thick starch envelope), fragmenting as cells age. Cells usually have one or more vacuoles. Aged cells accumulate carotenoids and turn brick red.Asexual reproduction occurs by 2—4—8—16 autospores. Sexual reproduction was not observed.The SEM images explained the approximately smooth cell wall surface.
This species differs fromC. tenuithecaby the ellipsoidal and ovoid cell shape in young age, and differs fromC. vacuolataby the independent phylogenetic position based on 18S+ITS andtufAgene phylogeny.
Holotype.HBI SHY432, March 2014, growing on the surface of moist stone in the dark, leg. Huiyin Song.
Authentic culture.Culture strain FACHB-3315 is deposited at FACHB (http://algae.ihb.ac.cn/).
Type locality.Jinyun mountain, Chongqing Municipality (29°50′22″N—106°23′18″E).
Habitat.Aerial or sub-aerial habitat such as wet stone surfaces.
Distribution.Chongqing & Zhoukou, China.
Etymology.The species epithet ‘chongqingensis’(fem. adj.) refers to the type locality Chongqing Municipality
營養(yǎng)細胞多為單細胞, 細胞壁近似光滑, 不具有單星藻屬典型的縱向分布的肋紋。幼期細胞多呈橢球型或卵形, 少數近似球形, 末端鈍圓, 一個周生片狀色素體, 細胞大小約(6—9) μm×(5—7) μm。成熟細胞多呈球形或橢球型, 直徑約8—14—(18) μm。色素體周生具一個明顯蛋白核, 蛋白核外裹有厚淀粉鞘, 色素體隨細胞成熟逐漸碎片化。細胞通常具一至多個液泡。多通過形成2—4—8—16個似親孢子進行無性生殖, 有性生殖未觀察到。
該種與近緣種薄壁單星藻相比, 幼期細胞多呈現橢球型或卵形, 薄壁單星藻幼期細胞多呈圓球狀。與液泡單星藻相比, 該種具有獨立的系統(tǒng)發(fā)育學位置(基于18S+ITS及tufA序列)。
正模標本: 甲醛固定標本HBI SHY432保藏于中國科學院水生生物研究所, 標本采集于陰暗處潮濕石頭表面, 采集人宋會銀。
培養(yǎng)標本: 編號FACHB-3315, 保藏于中國科學院水生生物研究所淡水藻種庫。
模式標本產地: 重慶縉云山(29°50′22″N—106°23′18″E)。
生境: 附生于潮濕石頭表面等氣生、亞氣生環(huán)境。
分布: 中國重慶、河南周口等地。
詞源學: 新種主要基于樣品采集地命名。
2.2 系統(tǒng)發(fā)育學分析
基于18S+ITS序列系統(tǒng)發(fā)育分析結果示于圖 4,序列比對產生包含53條序列的數據集, 經手動刪除空位后得到長度1878 bp的矩陣, 其中包含保守位點1545 bp(82.27%), 可變位點332 bp(17.68%), 簡約信息位點265 bp(14.11%)。堿基組成分析結果顯示平均的堿基組成為A=25.44%, G=27.04%, C=21.33%,T=26.19%, AT含量(51.65%)。系統(tǒng)樹重建選取4個水網藻科成員作為外類群。

圖4 基于18S rDNA和ITS序列聯(lián)合構建的柵藻科系統(tǒng)樹Fig. 4 Phylogenetic tree of Scenedesmaceae based on 18S+ITS sequences
基于tufA基因的系統(tǒng)發(fā)育分析結果示于圖 5,序列比對產生包含40條序列的數據集, 經手動刪除空位后得到長度736 bp的矩陣, 其中包含保守位點488 bp(66.3%), 可變位點248 bp(33.7%), 簡約信息位點206 bp(27.99%)。堿基組成分析結果顯示平均的堿基組成為A=36.68%, G=19.84%, C=13.86%,T=29.62%, AT含量(66.98%)高于GC含量(33.70%)。系統(tǒng)樹重建選取2個水網藻科成員作為外類群。

圖5 基于葉綠體tufA基因構建的柵藻科系統(tǒng)樹Fig. 5 Phylogenetic tree of Scenedesmaceae based on tufA gene
18S+ITS序列系統(tǒng)發(fā)育分析結果表明, 單星藻屬與星網藻屬AsterarcysComas (即廣義單星藻分支Coelastrella sensu lato)在柵藻科內能夠形成獨立單系分支, 且分支獲得較高的支持值(100/1)。原單星藻屬OriginalCoelastrella與原擬螺翼藻屬OriginalScotiellopsis聚為一支共同構成核心單星藻分支, 并獲得較高的支持值(99/1)。本研究所提供2株重慶單星藻藻株(FACHB-3315和FACHB-3314)與液泡單星藻具有較近的親緣關系, 且二者在形態(tài)學上也較為相似, 即細胞壁不具有核心單星藻類群典型的縱向肋紋。此外, 薄壁單星藻也具有類似的形態(tài)型, 但三者在系統(tǒng)樹中并未形成單系分支, 而形成并系類群。基于葉綠體tufA基因的系統(tǒng)發(fā)育分析結果與18S+ITS序列分析結果基本一致, 同樣表明單星藻屬與星網藻屬成姐妹類群, 星網藻屬位于廣義單星藻屬分支基部。此外, 重慶單星藻與液泡單星藻具有最近的親緣關系, 二者形成姐妹分支, 進一步與薄壁單星藻聚為一支且具有較高的分支支持值(95/0.96), 三者在單星藻屬內形成單系分支并代表了單星藻屬的第三種形態(tài)型, 因此, 基于tufA基因的系統(tǒng)發(fā)育研究結果與形態(tài)學特征更為吻合。
3 討論
廣義單星藻分支Coelastrella sensu lato概念最初是由捷克藻類學家Kaufnerová 和Eliá?基于18S rDNA序列的系統(tǒng)發(fā)育研究結果提出的[15]。其中主要包含核心單星藻分支“Core”Coelastrella及部分邊界類群, 及星網藻屬模式種四細胞星網藻Asterarcys quadricellulare。其中核心單星藻分支則主要包括原單星藻屬OriginalCoelastrella和原擬螺翼藻屬OriginalScotiellopsis。作者前期對廣義單星藻分支的研究, 確定了該分支實際包含了原柵藻科內5個屬的成員, 并對應了至少4種形態(tài)型。第一種形態(tài)型(Morphotype 1)主要指原單星藻屬的形態(tài)型,細胞球形或寬橢球型, 沒有或很少出現細胞頂點增厚, 細胞壁縱向肋紋多達16—40個; 第二種形態(tài)型(Morphotype 2)指原擬螺翼藻屬和伊那藻屬EnallaxPascher形態(tài), 細胞紡錘形或檸檬狀, 具頂點增厚, 肋紋數一般較少約4—12(20)個。第三種形態(tài)型(Morphotype 3)則主要指液泡單星藻、薄壁單星藻及重慶單星藻的形態(tài), 細胞壁近似光滑或僅具細微網紋, 無單星藻屬典型的縱向肋紋。第四種形態(tài)型指星網藻屬形態(tài)型, 即四細胞群體, 群體外具膠被[17]。
重慶單星藻最明顯的形態(tài)特征是細胞壁不具典型的縱向肋紋, 接近單星藻屬的第三種形態(tài)型,掃描電鏡中細胞壁出現的細密皺紋推測是由于制樣過程的細胞壁失水皺縮形成的, 不作為典型特征予以討論。重慶單星藻與近緣種薄壁單星藻的形態(tài)區(qū)別, 主要為薄壁單星藻幼期或成熟細胞均呈圓球狀[17], 而重慶單星藻幼期細胞多呈橢球型或卵形,成熟細胞呈橢球形或圓球形。重慶單星藻與液泡單星藻的形態(tài)區(qū)別主要表現為, 液泡單星藻細胞壁具細微網紋[31,32], 而重慶單星藻未見規(guī)則網狀細紋。
根據前人的研究, 星網藻屬位于廣義單星藻屬分支內部[15], 因此星網藻屬的成立一直以來都有所爭議。本研究基于18S+ITS序列的系統(tǒng)發(fā)育研究結果表明, 單星藻屬與星網藻屬確實具有較近的親緣關系, 與之前研究結果中單星藻屬與星網藻屬呈多系分布[15]略有不同的是, 在本研究中星網藻屬與單星藻屬互相分離形成獨立分支, 該結果與tufA基因的系統(tǒng)發(fā)育研究結果更為一致, 與形態(tài)學特征也較為吻合(因星網藻為四細胞群體, 群體外具膠被, 而目前所發(fā)現的所有單星藻均不具膠被)。因此, 上述結果進一步支持四細胞星網藻從廣義單星藻分支中獨立成屬。在18S+ITS系統(tǒng)樹中重慶單星藻與另外兩種細胞壁相對光滑的物種液泡單星藻、薄壁單星藻未形成單系分支, 而形成并系類群。但是在tufA基因的系統(tǒng)樹中三者形成單系分支, 代表了單星藻屬的第三種形態(tài)型。鑒于核基因18S rDNA和ITS序列在該類群系統(tǒng)發(fā)育研究中的局限性[17,33,34],推測近緣分支拓撲結構可能較易隨序列增減而出現變動。因為, 對于柵藻類群, 18S序列往往過于保守, 所含有效系統(tǒng)發(fā)育信息位點較少, 而ITS序列變異度過高, 序列較易達到突變飽和, 也不適于科級別的系統(tǒng)發(fā)育分析, 但ITS序列在屬及屬以下水平具有較高的分辨率, 可作為區(qū)分種和亞種的有力證據。而葉綠體tufA基因因有效系統(tǒng)發(fā)生信號較多,系統(tǒng)樹拓撲結構更為穩(wěn)定, 可作為該類群更有效的系統(tǒng)發(fā)生重建手段。
基于以上形態(tài)學和系統(tǒng)發(fā)育研究結果, 本研究建議將采集于重慶縉云山的單星藻作為1個新種提出, 命名為重慶單星藻Coelastrella chongqingensissp. nov., 該新種的發(fā)現擴充了單星藻屬第三種形態(tài)型的物種組成。鑒于利用形態(tài)特征鑒定此類單細胞球狀綠藻時, 可供區(qū)分的特征較少且在不同培養(yǎng)狀態(tài)下還存在一定的形態(tài)可塑性, 對于該類群的物種水平鑒定, 建議將ITS和tufA分子數據作為更可靠的鑒定依據。
猜你喜歡
故事作文·低年級(2023年12期)2023-12-25 02:50:32
環(huán)球人文地理(2022年8期)2022-09-21 03:49:42
公民導刊(2022年4期)2022-04-15 21:03:14
當代黨員(2022年6期)2022-04-02 03:14:56
意林·全彩Color(2019年11期)2019-12-30 06:08:38
當代黨員(2019年19期)2019-11-13 01:43:29
重慶行政(公共人物)(2018年5期)2018-11-06 07:42:18
城市地理(2016年6期)2017-10-31 03:42:32
重慶文理學院學報(社會科學版)(2017年5期)2017-10-23 01:30:02
今日重慶(2017年5期)2017-07-05 12:52:25
