甜角大小孢子發生和雌雄配子體發育及受精過程
2021-12-04 07:43:04夏富嫻李夢雪李體初楊時宇王兵益
云南大學學報(自然科學版) 2021年6期
夏富嫻,李夢雪,李體初,楊時宇,王兵益**
(1.中國林業科學研究院 資源昆蟲研究所,云南 昆明 650224;2.南京林業大學,江蘇 南京 210037)
甜角(Tamarindus indicaL.)又名羅望子、酸豆、酸梅,豆科(Leguminosae)酸豆屬(TamarindusL.)的熱帶亞熱帶常綠大喬木,原產地為非洲,后傳至其它大洲[1].甜角是單種屬植物,包括兩種果實類型,甜型和酸型.酸型甜角很早以前就被引種到中國,主要分布于云南、海南、廣州、廣西、福建、臺灣等地;甜型是20世紀末才從泰國引種到國內,主要栽培區域在云南南部及干熱河谷地區[2].甜角果實是特色熱帶水果,含多種營養物質,包括多種有機酸、糖類、維生素、蛋白質、游離氨基酸和礦質元素等,鮮食生津止渴,清熱解暑,消食化積,可加工成果脯、蜜餞、飲品等[3].種子中富含羅望子多糖,可制成羅望子膠,是一種優質的食品添加劑[4].所以,甜角具有良好的綜合經濟利用價值和前景.
甜角的經濟價值主要體現在其果實上,然而,作為新引進物種,甜角在種植過程中一直存在開花量大,但結實率和果實產量低的問題.基于課題組之前調查統計,甜角自然結實率僅為7%[5].成功的有性生殖過程保證了植物的結實,很多學者認為,胚胎發育進程受到阻礙是植物生殖失敗的重要原因之一[6].目前,國內外甜角的研究主要集中在果實營養、化學成分分析、藥理等方面[7-9].在胚胎學方面的研究還鮮有報道. 為此,本研究采用石蠟切片法和熒光顯微觀察法對甜角大小孢子發生和雌雄配子體發育及受精過程進行觀察,旨在闡明甜角大小孢子發生和雌雄配子體發育過程,為甜角生殖生物學積累資料,并為了解其結實率低的原因和解決該問題提供細胞學證據,以期為后續的豐產措施提供理論基礎.
1 試驗地概況
實驗所用材料為引種自泰國,種植于云南省昆明市中國林業科學研究院資源昆蟲研究所元江試驗站內(102°22′E,23°55′N,海拔390~420 m)22年生的甜角植株.試驗站區屬于北熱帶干熱河谷氣候類型,年平均溫度12~24℃,最冷月平均氣溫7~17℃,最熱月平均氣溫16~29℃,年平均降水量770~2 400 mm,土壤為砂紅壤,呈微酸性至中性.
2 材料與方法
2.1 組織切片選取植株中不同發育階段的花蕾和花朵,保存于FAA固定液中(V(甲醛)∶V(冰乙酸)∶V(70%乙醇)=1∶1∶18)固定24 h,4℃冰箱中保存備用.采用常規石蠟切片法制片[10]. 利用Leica RM輪式切片機切片,切片厚度為8~10μm,經番紅固綠(Safranin-Fast Green)染色法染色,中性樹膠封片后用光學顯微鏡觀察并拍照.
2.2 花粉粒形態觀察采集甜角花朵,用解剖針輕輕撥動花藥壁,使花粉粒散落到導電膠上,離子濺射儀(JS-1600)噴金鍍膜后,用TM3000掃描電鏡進行觀察并拍照.
2.3 甜角受精作用花粉管生長動態觀察在開花前1 d對即將開放的200個花蕾去雄套袋并進行掛牌標記,于開花當天10:30—11:00進行授粉.于授粉后1、2、3、4、5、6、7、8、9、20、24 h分別采集雌蕊各10個樣本放于卡諾固定液(V(無水乙醇)∶V(冰醋酸)=3∶1)中固定并帶回實驗室,用2 mol·L-1的NaOH溶液進行透明和軟化3 h,再用苯胺藍溶液(質量分數0.1%)染色1 h后壓片,用OLYMPUS LX71型熒光顯微鏡觀察花粉的萌發和花粉管的生長并拍照.
3 結果與分析
3.1 花藥壁的發育甜角花兩性,每朵花有3枚雄蕊,雌蕊1枚(圖1(a)~(c)).切片結果顯示花藥有4個花粉囊,呈左右不對稱型并排排列,相鄰兩個花粉囊之間被藥隔分離(圖2(b)).花藥發育初期的結構比較簡單,由外側的表皮和內側的分生組織兩部分構成,隨著花藥的發育,在花藥的4個角隅處有一群體積較大、細胞質較濃、細胞核明顯的細胞,這就是孢原細胞(圖2(a)).孢原細胞經過1次平周分裂后形成初生壁細胞和初生造孢細胞.初生壁細胞經1次平周分裂和多次垂周分裂后形成內外2層細胞,隨著進一步發育內層細胞形成絨氈層細胞,外層細胞形成次生壁細胞.次生壁細胞隨后發育形成花藥的中層和藥室內壁.5層花藥壁形成,由外向內依次是表皮、藥室內壁、中層(2層)和絨氈層(圖2(j)).觀察結果顯示,甜角花藥壁的發育類型屬于基本型.

圖1 甜角花部結構Fig.1 The floral structure of T.indica
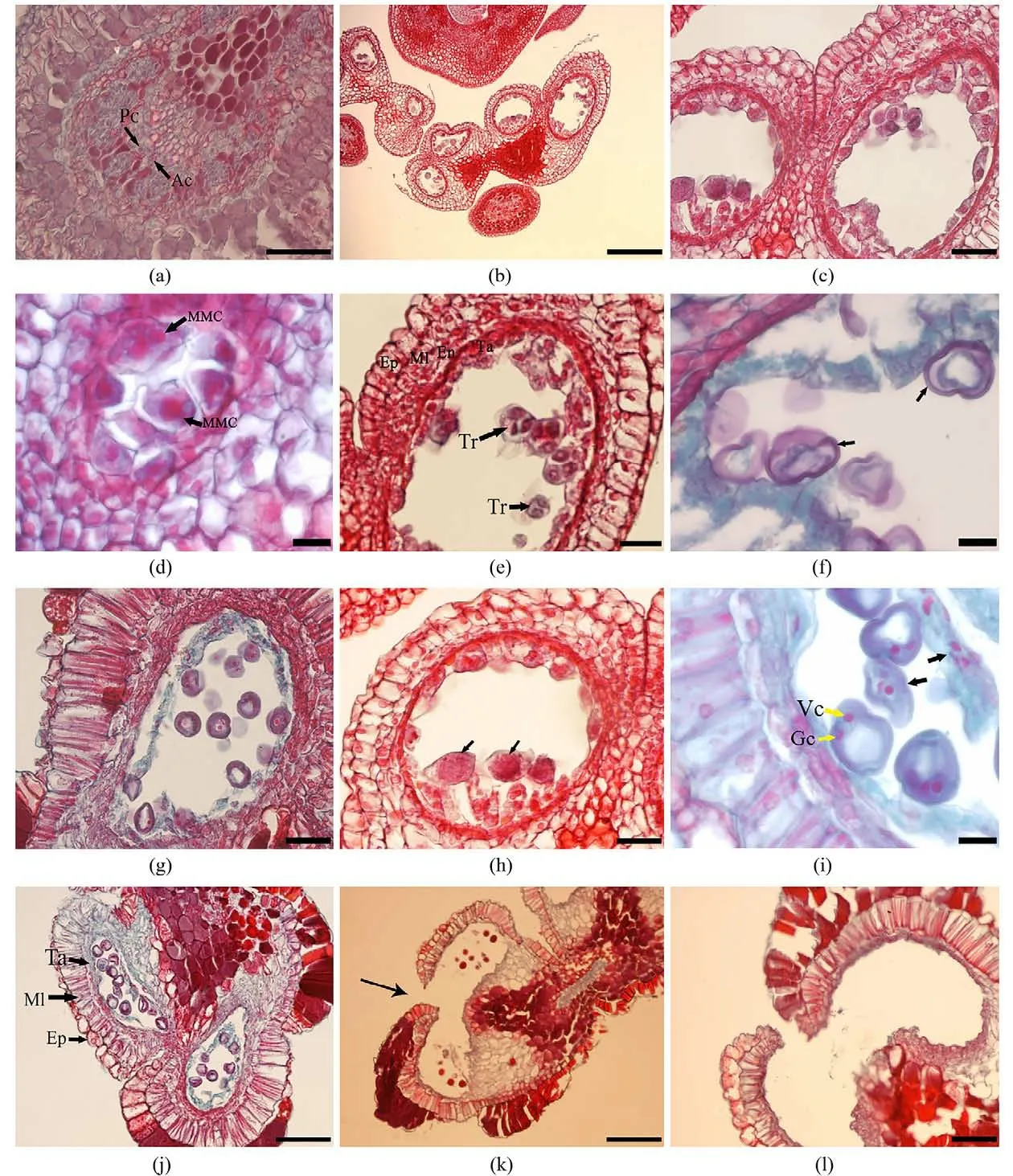
圖2 甜角小孢子發生和雄配子體發育Fig.2 The microsporogensis and development of male gametophyte of T.indica
在小孢子母細胞形成的時期,表皮細胞進行分裂并液泡化,這一時期細胞特征明顯,其徑向壁延長,垂周壁加厚呈長方形狀,并且細胞排列緊密,核大(圖2(h)).至后期小孢子細胞成熟后,部分表皮細胞逐漸解體,僅留下波浪狀的角質層細胞(圖2(j)).觀察發現,甜角的絨氈層屬于變形絨氈層,當小孢子母細胞處于減數分裂的過程中時絨氈層的細胞從花藥壁脫離,成團移至花藥腔內(圖2(g)).當小孢子成熟時,同側的2個花粉囊連通,在2個花粉囊相連處的藥室壁漸漸變薄裂開,花粉散出(圖2(k))
3.2 小孢子發生和雄配子體形成在花藥壁發育的時候,小孢子同步發生,初生造孢細胞經過數次的有絲分裂,形成次生造孢細胞.這一時期的次生造孢細胞體積大,排列緊密,細胞質濃厚,形狀多呈多邊形.其進一步發育后形成小孢子母細胞,初期的小孢子母細胞排列緊密,發育到一定程度后因為胼胝質的沉積會使它們彼此分離開形成橢圓形、細胞核大、核仁明顯、細胞質濃厚的單個游離小孢子母細胞(圖2(c),(d)).隨后,游離的小孢子母細胞進入減數分裂期,第1次減數分裂后形成2個子核,但這個時期并沒有細胞壁的形成,細胞直接進入第2次減數分裂形成4核,這一時期因為彼此間有胼胝質的相互隔離,形成四分體. 甜角小孢子四分體的排列方式以四面體型為主,偶見左右對稱型(圖2(e)),胞質分裂為同時型.四分體存在的時間較短,發育至后期,彼此間的胼胝質逐漸解體,形成4個體積小,形狀不規則的游離小孢子,在此期間,絨氈層細胞逐漸解體(圖2(f)).隨后,小孢子細胞進一步發育形成一個體積較大圓形有中央大液泡的細胞.在此期間,中央大液泡會將細胞核擠向細胞壁的一側,此時進入單核靠邊期;單核靠邊期的小孢子細胞的細胞核進行一次不均等的有絲分裂,形成一個圓的較大的營養核和一個小的生殖核.之后,細胞內營養物質不斷積累,中央大液泡逐漸消失,形成2-細胞型的成熟花粉粒,此時觀察到絨氈層細胞并未完全降解(圖2(i)).
通過對甜角小孢子母細胞發育過程的觀察,發現同一花藥不同藥室間以及同一藥室內的小孢子母細胞發育進程具有不同步現象(圖2(h)).在甜角雄配子體的發育過程中,觀察到小孢子在發育過程中出現細胞核降解、細胞質濃縮及細胞皺縮的敗育現象(圖2(i)).在觀察中還發現很多花粉囊中花粉粒數目很少,這部分花粉囊中單個花藥腔內約只有8~10粒花粉(圖2(k)),此外,還觀察到部分花粉囊為空室(圖2(l)).
3.3 花粉形態觀察通過掃描電鏡觀察,甜角花粉為單粒花粉(圖3(a)). 赤道面觀為長球形(圖3(b)),極面觀呈3裂鈍三角形(圖3(c)、(d)).于甜角的開花期隨機選取若干花粉粒進行測量,并參照G.Erdtman對花粉形狀類型的劃分標準(P*E值表示花粉粒的大小,P代表極軸長,E代表赤道軸長;P/E值在1.33~2.0之間屬于長球形,P/E值大于2.0屬于超長球形)[11].結果顯示甜角P為(49.77±1.24) μm,E為(33.32±0.98) μm,P*E值為1 658.34 μm2,P/E值為1.49,所以甜角花粉粒的形狀是長球形或近長球形.萌發溝3裂延伸至兩極、溝痕整體呈長條形,寬度1.8~2.4μm.花粉粒外壁瘤狀紋飾緊密交錯排列分布.

圖3 甜角花粉形態掃描電鏡觀察Fig.3 Observation of pollen morphology by scanning electron microscope
甜角為單屬種,甜角花粉形態的研究對酸豆屬花粉形態特征具有代表性.所以將甜角花粉粒的P和E與豆科中其他5個屬花粉特征資料進行了比較分析[12-14].發現,酸豆屬花粉的P和E與其他5屬相比較存在顯著差異,且酸豆屬P值明顯大于其他5屬(圖4,不同屬之間相同字母表示差異不顯著p>0.05,不同字母表示差異顯著p<0.05),說明同科不同屬之間花粉形態差異較大.
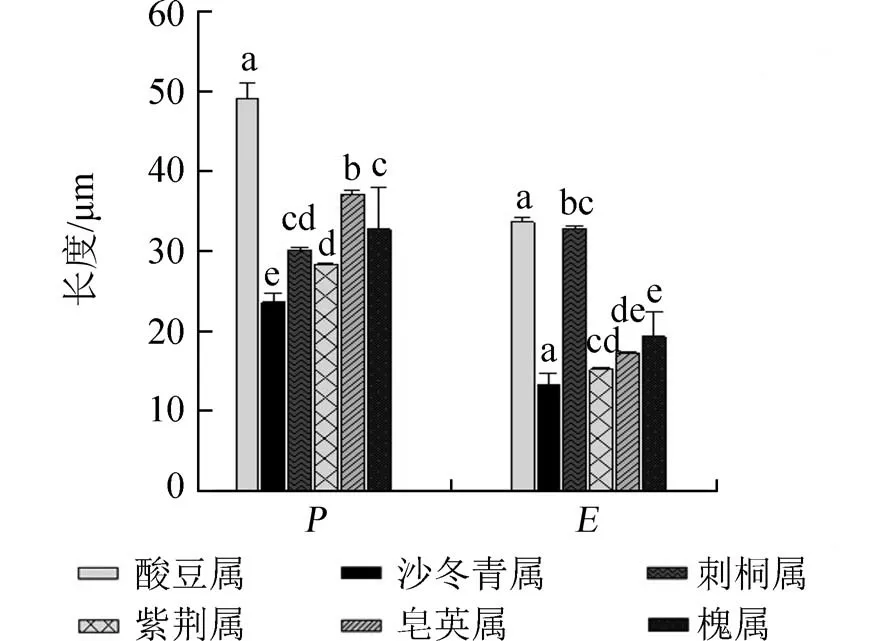
圖4 酸豆屬與豆科其他屬花粉極軸和赤道軸長比較Fig.4 Comparison of pollen polar and equatorial axis lengths between Tamarindus and other Leguminous genera
3.4 大孢子發生和雌配子體的形成甜角為單心皮雌蕊,子房一室,邊緣胎座,厚珠心,胚珠彎生,著生在腹縫線的邊緣上排成縱列.胚珠原基開始發育時,由薄壁組織細胞組成的珠心組織形成,近珠心基部形成內、外珠被原基,進而形成內、外雙珠被(圖5(a)).隨后,伴隨細胞的分裂生長在珠心表皮下形成一個體積大、原生質濃、有大細胞核與周圍細胞差異明顯的孢原細胞(圖5(b)).孢原細胞進一步快速分裂發育形成一個體積更大、原生質更濃厚的大孢子母細胞(圖5(c)).隨著胚囊進一步發育大孢子母細胞分裂后形成4個大孢子.4個大孢子中只有靠近合點端的1個大孢子繼續發育形成功能大孢子,其余3個大孢子退化消失.之后,功能大孢子生長膨大經有絲分裂后形成二核胚囊(圖5(d),(e)).在此時期,二核胚囊的2個核因中央大液泡的作用逐漸移向胚囊的兩端(圖5(f)).二核胚囊再經過有絲分裂形成四核胚囊(圖5(g)),四核胚囊繼續進行分裂形成八核胚囊(圖5(h)).
八核胚囊在較短的時間里繼續生長發育形成細胞壁,此時胚囊發育成熟.觀察發現,甜角胚囊發育為單孢子-蓼型胚囊,為比較原始的胚囊發育類型,是由1個功能大孢子發育形成7胞8核胚囊,包括珠孔端的2個助細胞和1個卵細胞、1個二倍體中央細胞和合點端的3個反足細胞.2個助細胞呈對稱排列,具大液泡.卵細胞比助細胞小,易于辨認.二倍體的中央細胞液泡化明顯(圖5(i)).3個反足細胞體積較大,具2核.實驗觀察到甜角八核胚囊存在時間較短,胚囊成熟后不久3個反足細胞退化消失(圖5(j));2個助細胞在受精前有開始退化解體的趨勢,最后消失只留下卵細胞和中央細胞(圖5(k)).觀察中發現胚囊有敗育現象(圖5(l)).
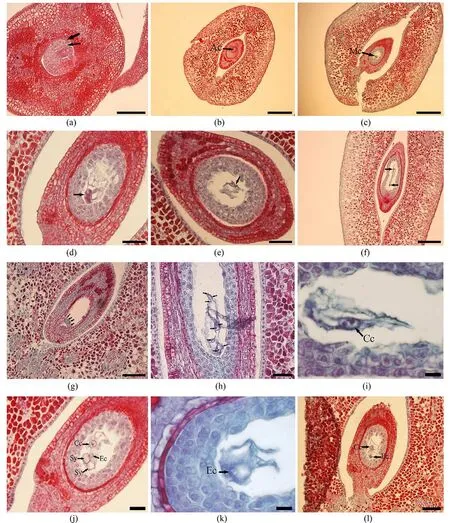
圖5 甜角大孢子發生和雌配子體發育Fig.5 The megasporogenesis and development of female gametophyte of T.indica
3.5 受精作用甜角花粉粒萌發時,在熒光染色下顯示黃綠色熒光,隨著花粉管的伸長,花柱內壁以及子房中也會呈現黃綠色熒光.授粉后30 min,柱頭表面粘附大量花粉粒,但花粉粒未見明顯變化(圖6(a));授粉后1 h,部分花粉粒在柱頭表面水合開始萌發,可觀察到零星未穿過柱頭表面的萌發的花粉管(圖6(b));授粉后2 h,萌發的花粉粒數量明顯增多,并有少量的花粉管穿過柱頭表面的乳突細胞開始進入花柱,但熒光現象不強且主要集中在柱頭處(圖6(c));授粉后3 h,大量花粉粒已經萌發,熒光現象明顯,許多花粉管穿過柱頭表面沿引導組織開始生長(圖6(d)),在花柱中可明顯看到帶有生殖核的花粉管(圖6(e));授粉后4 h,柱頭表面花粉粒基本已全部萌發,花粉管以束狀形式向花柱基部延伸,少量花粉管已生長至花柱長度的3/4(圖6(f));授粉后5 h,可觀察到柱頭表面幾乎已無花粉粒萌發,花柱中花粉管的熒光寬度約為上一時間段的2倍,整個花柱內壁十分明亮(圖6(g)),表明該時間段為花粉管伸長的最旺盛時期,此時,胚囊中中央細胞緊緊挨著卵細胞,卵細胞挨近中央細胞;授粉后6 h柱頭和花柱中熒光現象開始減弱,花粉管離開引導組織,穿出隔膜進入子房腔內,沿著珠柄生長,伸入子房,從珠孔進入胚囊內部,釋放出2個精細胞;然后,一個精細胞會向中央細胞靠近,另一個精細胞會向卵細胞靠近,精核緊貼卵細胞,其精核內容物會逐漸進入卵細胞內,此時精子與極核也完成融合,形成一個較大的細胞完成雙受精過程,甜角的受精模式為珠孔受精(圖6(h)).花柱中束狀的花粉管緩慢呈現扭曲、斷裂、不連續狀態,柱頭表面附著的花粉粒逐漸失去熒光效果,此狀態為受精結束后花粉管解體的初期,甜角授粉12 h后花柱中僅可見極少量的、扭曲的花粉管,且花粉管解體、撤退的態勢越發明顯,柱頭表面的花粉粒數量明顯減少,此狀態為受精結束后花粉管解體的末期(圖6(i));授粉后24 h,柱頭及花柱中的熒光消失,花粉管完全解體.
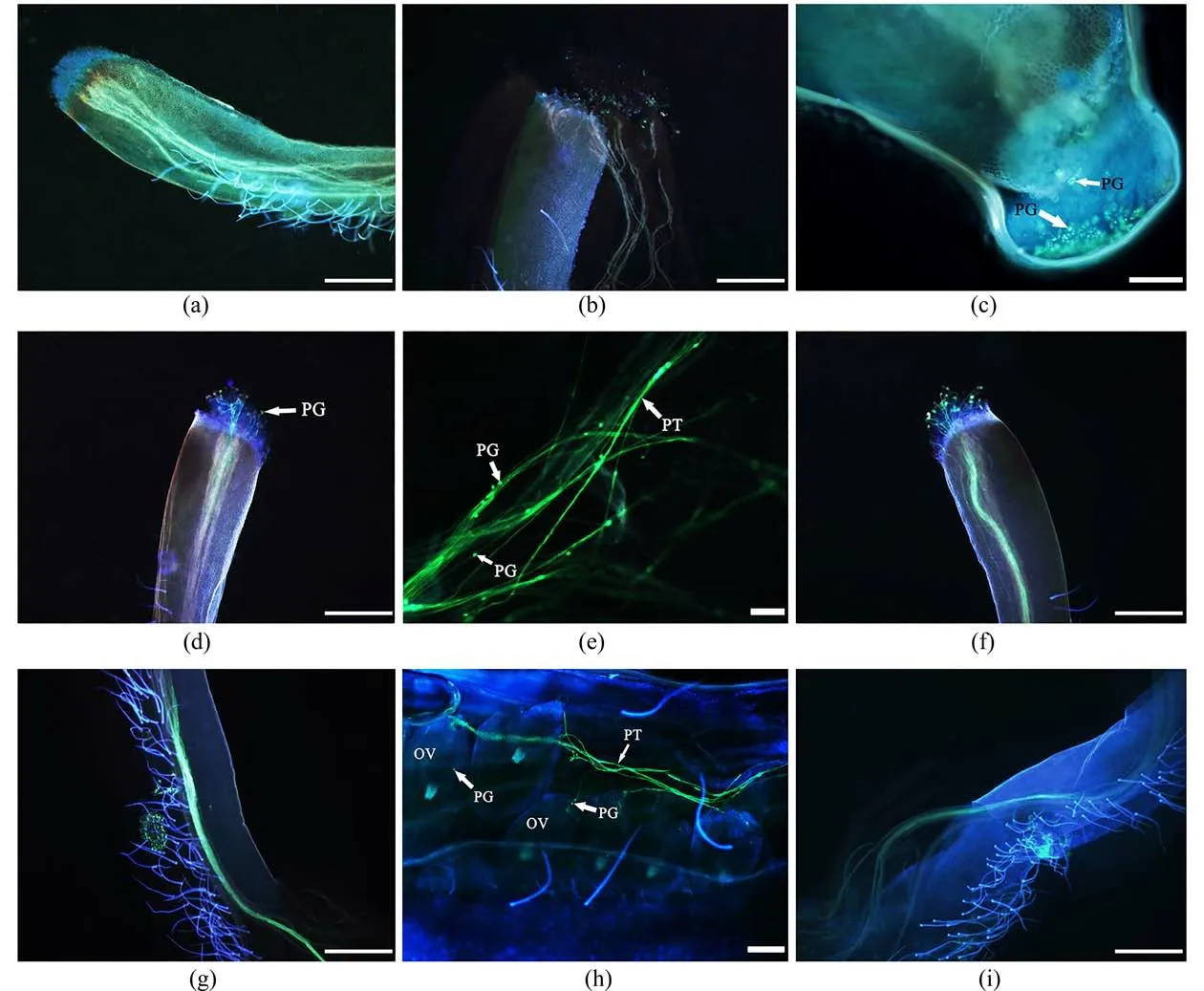
圖6 甜角花粉萌發、和花粉管生長及受精過程Fig.6 Pollen germination, pollen tube growth and fertilization process of T.indica
4 討論
植物胚胎學特征是植物系統進化過程中比較穩定的性狀,是判定植物親緣關系、系統進化進程的重要依據[15].比較酸豆屬甜角與豆科中的豇豆屬(VignaSavi)植物綠豆(V.radiata(L.)Wilczek)[16-17]和紅小豆(V.angularis(Willd)Ohwi et Ohashi)[18],兩型豆屬(AmphicarpaeaElliot)的兩型豆(A.edgeworthiiBenth.)[19],沙冬青屬(AmmopiptanthusS.H.Cheng)的矮沙東青(A.mongolicus(Kom.)S.H.Cheng)[20],黃芪屬(AstragalusL.)的黃芪(A.membranaceus(Fish.)Bunge)[21]的胚胎學特征,發現它們具備很多一致的特征,如花藥具有4個花粉囊,花藥壁由5層細胞構成,從外至內依次為1層表皮、1層藥室內壁、2層中層和1層絨氈層,成熟花粉粒為2-細胞花粉,子房一室,邊緣胎座,雙珠被,厚珠心,胚珠彎生,胚珠多數,單孢原直接發育為大孢子母細胞,胚囊發育為8個時期,即孢原細胞形成期、大孢子母細胞形成期、大孢子母細胞減數分裂期、功能大孢子形成期、單核胚囊形成期、胚囊有絲分裂期、8核胚囊發育期和成熟胚囊期,胚囊發育類型為蓼型等.比較研究反映了豆科各屬種之間系統發育上的一致性,說明他們有較近的親緣關系.本研究認為酸豆屬和上述幾屬的胚胎學特征雖然很多相一致,但在有些特征方面也有所不同.例如:甜角小孢子四分體多為四面體型,偶爾可見左右對稱型,而上述其它植物四分體都為四面體型;經觀察發現甜角花藥壁兩中層分別自內、外側次生周緣細胞平周分裂形成的兩層細胞,其發育類型為基本型,但黃芪藥壁發育類型為單子葉型,紅小豆為雙子葉型. 其次,甜角小孢子胞質分裂是同時型,兩型豆兼具同時型和連續型兩種類型. 研究認為2-細胞花粉,雙珠被,厚珠心,蓼型胚囊發育是比較原始的胚胎學發育特征,彎生胚珠和直生胚珠要比倒生胚珠進化[20].說明甜角在胚囊的形態發育中具有較明顯的原始特征,但胚珠的發育特征比較進化.甜角既具有相對原始的特征,又具有相對進化的特征,鑲嵌進化或異步演化明顯,我們認為酸豆屬以及豆科在被子植物中屬于相對較原始的類群.
花粉是遺傳信息的載體,具有較強的遺傳保守性,其固有的形態特征在科與屬之間對植物的系統分類地位具有一定實際意義[22].甜角為單屬種,對其進行花粉形態特征的研究填補了酸豆屬花粉形態學資料的空白和可為其準確分類提供重要依據.本研究中,甜角與豆科其他屬的花粉形態特征有明顯的差異性.甜角花粉外壁有瘤狀紋飾緊密排列,沙冬青屬、刺桐屬(ErythrinaL.)、紫荊屬(CercisL.)、皂莢屬(GleditsiaL.)、槐屬(SophoraL.)外壁都是網狀紋飾,且沙冬青屬花粉外壁具有多角形或圓形網眼.所以從孢粉學的觀點看將甜角單獨歸為酸豆屬是合理的.
甜角在減數分裂過程中,同一花藥不同藥室間以及同一藥室內的小孢子母細胞發育具有不同步現象.這種現象在很多植物中都有發現,例如柿品種‘禪 寺 丸’(Diospyros kaki‘Zenjimaru’)[23]、楸 樹(Catalpa bungeiC.A.Meyer)[24]、車桑子(Dodonaea viscosa(L.)Jacq.)[25]、‘早 籽 銀 桂’(Osmanthus fragrans(Thunb.)Lour.‘Zao Zi Yin’)[26]、‘晚銀桂’(Osmanthus fragrans(Thunb.)Lour.‘Wan Yin’)[27]等中也有出現.這種不同步現象使花粉散播的總時間延長,從而使傳粉和受精的時間也有所延長的一種適應機制,可能是植物長期適應自然環境而形成的一種生殖策略.
成功的有性生殖過程保證了植物群體的結實,從而保證了植物群體的擴大和經濟效益的提高.目前研究表明,胚胎發育進程受到阻礙是植物生殖失敗的重要原因之一[28].本研究觀察到甜角有空花粉囊現象,空花粉囊的出現可能是由于雄配子體在其生長環節出現異常狀況,無法發育成成熟花粉所致.絨氈層在花粉發育的過程中起著重要的作用,它會隨著花粉的發育通過溶解自身為花粉發育提供所需要的營養物質.絨氈層細胞無法正常生長、過早退化或后期退化不完全,都會使小孢子母細胞的發育出現異常,導致花粉敗育[29].研究表明,毛白楊(Populus tomentosaCarrière)絨氈層細胞Ca2+-ATPase的分布異常會影響絨氈層細胞的正常PCD(programmed cell death)進程,使絨氈層細胞無法正常降解,導致花粉的敗育;擬南芥(Arabidopsis thaliana(L.)Heynh.)在高溫環境下,絨氈層PCD進程會出現滯后現象,因絨氈層后期無法徹底降解,導致不能產生正常的花粉粒[6,30].我們的研究發現,甜角花粉發育過程中小孢子在發育過程中出現細胞核降解、細胞質濃縮及細胞皺縮的敗育現象.還觀察到2-細胞花粉形成時期絨氈層出現降解不完全現象.因此推斷,甜角胚胎發育過程中可能絨氈層生長發育異常,降解滯后使花粉發育過程中不能吸收足夠的營養物質形成成熟的花粉粒,造成了花粉敗育現象,從而降低了花粉數量,影響其后期的傳粉生殖過程,推斷這可能是甜角多花少實的原因之一.此外,本研究中甜角生長于干濕季分明的干熱河谷地區,氣候特殊,夏季高溫干旱且漫長.因此,我們推測甜角絨氈層的生長發育異常,可能與其生長環境因素有關,這還有待于進一步研究.這將為干熱河谷地區甜角的進一步開發利用提供一定的理論依據.
受精是植物生殖生長的重要過程,它不僅是植物界有性生殖最進化的形式,也是保證植物生殖成功的關鍵.植物的受精作用是一個動態的變化過程,每種植物均有不同的特性.用熒光顯微鏡是觀察植物從花粉落到柱頭開始萌發到花粉管生長伸長到達子房進入胚囊完成受精情況最直觀和最有效的方式.本研究中花粉粒黏附在柱頭上1 h后能正常開始萌發,授粉6 h后,花粉管生長延伸到達子房從珠孔進入胚囊完成受精.之前研究認為,胚囊中的助細胞對花粉管生長延伸至子房具有主要的定向指引作用,因為助細胞分泌的多肽LURES對花粉管具有吸引作用[31].本研究中觀察到在受精之前2個助細胞退化消失.何孟元等[32]對豆科植物大豆(Glycine max(L.) Merr.)的胚胎學研究也發現受精前成熟胚囊中2個助細胞已經退化消失,有時只有1個退化,極少數出現2個助細胞都未退化的現象.楊維才等[33]對擬南芥的研究發現中央細胞中CCG(central cell guidance)基因的特異性表達是將花粉管從柱頭導向進入胚囊所必需的,研究還認為中央細胞可通過助細胞并協同助細胞一起完成對花粉管的導向作用.甜角雌配子體發育后期兩個助細胞退化,只留下中央細胞和卵細胞,推測其中央細胞在甜角受精過程中對花粉管的導向性具有重要作用.關于甜角具體的受精機制還有待進一步研究.
