射陽湖水環境現狀與沉積物氮循環功能基因研究
2021-11-25 10:11:50樊祥科高海優張曉倩李博文張朝暉張永江
生態與農村環境學報 2021年11期
鄭 浩,樊祥科,鐘 非,高海優,張曉倩,李博文,張朝暉,張永江
(1.江蘇省漁業技術推廣中心,江蘇 南京 210036;2. 南通大學生命科學學院,江蘇 南通 226019)
為加強水產種質資源及其生存環境的保護,近年來農業農村部積極推進水產種質資源保護區建設工作,已審定公布11批535處國家級水產種質資源保護區。在保護區內,除了采取禁捕措施以外,還需要為水生生物的生存繁衍提供適宜的生存環境[1],包括豐富的天然餌料和優良的水環境等。目前,有關水產種質資源保護區的研究主要集中在浮游動物群落調查和水環境質量評價兩個方面。梁正其等[2]基于錦江河國家級水產種質資源保護區浮游動物群落特征數據,評估了生物餌料資源現狀。羅建波等[3]對梁子湖、鄱陽湖和千島湖國家級水產種質資源保護區的富營養化狀況進行評價,發現3個保護區整體處于中營養狀態。劉凱等[4]調查了嬉子湖國家級水產種質資源保護區浮游動物群落特征和水環境質量狀況,探討了影響浮游動物群落組成的關鍵環境因子,并指出總氮是決定該水域所屬環境質量類別的關鍵指標。
射陽湖水域于2011年被列為國家級水產種質資源保護區(第4批)。然而,保護區內相關水環境基礎數據缺乏,僅有少量歷史數據資料,難以支撐保護區水產種質資源養護工作的開展。為避免出現重建輕管、只建不管的狀況,有必要系統開展射陽湖保護區水環境質量現狀調查。此外,底泥基本理化性質和氮循環微生物特征調查也亟待開展。底泥在水生態系統中充當著“源”或“匯”的雙重角色:外源輸入的氮素除了被水生生物吸收利用,還會通過物理、化學及生物作用沉降至水體底部;底泥中富集的氮素在一定條件下又會向上覆水釋放,對水環境質量造成影響[5-6]。同時,底泥還是微生物的天然載體,由底泥中微生物驅動的氮循環在維持湖泊生態平衡中發揮重要作用[7]。氮循環微生物參與了氨化、硝化和反硝化等生態過程,上述過程影響了水生態系統中氮的轉化和去除[8]。通過調查氮循環功能基因豐度,有利于掌握射陽湖保護區氮素的生物地球化學循環過程。另外,董志穎等[9]報道外源性氮素輸入可直接影響寡營養水體中氮循環功能基因豐度,因此,氮循環功能基因豐度可能對外源性氮素輸入具有一定指示作用。筆者研究在射陽湖保護區共布設11個站位,分析主要水體理化指標的時空變化規律,同時在核心區中心及外圍區域(共5個站位)開展底質及微生物群落現狀調查。采用實時熒光定量PCR技術,調查各站位底泥中氮循環細菌功能基因豐度的季節變化規律,并結合水質和底質調查結果,分析氮循環功能基因間及其與射陽湖水域主要環境指標間的相關關系。筆者研究將有助于全面了解射陽湖國家級水產種質資源保護區水質和底質狀況,以及氮循環微生物特征,并為保護區后續水環境管理和整治工作提供重要基礎數據和理論依據。
1 材料與方法
1.1 研究水域與樣品采集
射陽湖國家級水產種質資源保護區位于江蘇省揚州市寶應縣東郊,保護區是由4個拐點(33°16′35″ N,119°39′22″ E;33°16′15″ N,119°36′28″ E;33°19′53″ N,119°31′45″ E;33°20′14″ N, 119°37′41″ E)順次連線圍成的水域,總面積為666.7 hm2。其中,核心區面積為100 hm2,為由5個拐點(33°18′42″ N,119°37′20″ E;33°18′31″ N, 119°37′31″ E;33°18′34″ N,119°37′12″ E;33°18′45″ N,119°37′09″ E;33°18′57″ N, 119°37′12″ E)順次連線圍成的水域(圖1)。
射陽湖保護區河網發達,周圍分布有安豐河、走馬河、大三王河、楊家河和鴿子河等河流。保護區核心區靠近村鎮,且周邊水產養殖業和種植業發達。根據保護區現狀,在核心區外圍(S1~S6)和核心區內(S7~S11)共布設11個站位。在核心區外圍,S1、S5和S6站位靠近村鎮,而S2、S3和S4站位靠近農業區;在核心區與各條支流交界處分別設置S7~S10站位,S11站位位于核心區中心。分別于2018年5月(春季)、8月(夏季)、11月(秋季)和2019年2月(冬季),開展射陽湖水域水體與沉積物環境質量現狀調查。調查期間,分別從11個站位采集水樣,同時選擇S1、S2、S3、S4和S11共5個站位(由于底泥疏浚,S5站位未采集到底泥樣品),使用抓斗式底泥采樣器對 0~10 cm表層沉積物進行采集,每個站位采集 3個混合樣,混合后分為兩份封裝,并分別經冷藏和冷凍帶回實驗室用于后續理化和微生物分析。
1.2 理化指標測定
采用EXO多參數水質監測儀(YSI EXO2)現場測定水體溫度(T)、pH值、溶解氧(DO)和葉綠素(Chl a)濃度。采用自動化學分析儀(CleverChem anna)測定水樣中氨氮(NH4+-N)、硝酸鹽氮(NO3--N)、亞硝酸鹽氮(NO2--N)和正磷酸鹽(PO43--P)濃度。使用ArcGIS 10.2軟件,采用插值法繪制NH4+-N、NOx--N和PO43--P濃度時空分布圖。水體總氮(TN)、總磷(TP)和高錳酸鹽指數(CODMn)檢測參照《水和廢水監測分析方法》[10]。另外,取新鮮底泥10.0 g放入100 mL三角瓶,加入2 mol·L-1KCl溶液50 mL,用橡皮塞塞緊,振蕩30 min,過濾后使用自動化學分析儀測定浸出液中NH4+-N濃度。風干后的底泥樣品中全碳、全氮和全硫含量使用元素分析儀(Elemantar vario)檢測。
1.3 樣品基因組總DNA提取和PCR擴增
使用E.Z.N.A.?soil DNA Kit(Omega Bio-tek)試劑盒提取底泥樣品基因組DNA,用質量濃度為10 g·L-1的瓊脂糖凝膠電泳檢測抽提DNA的完整性,并用NanoDrop 2000 分光光度計(Thermo Scientific)檢測DNA濃度和純度。
用引物分別擴增細菌16S rDNA和氮循環細菌功能基因(AOB-amoA、nxrA、narG、nirK、nirS和nosZ),qPCR擴增引物見表1。

表1 該研究所用擴增引物
qPCR反應采用20 μL體系,包括10 μL 2×SYBR Premix Ex TaqTM(TaKaRa)、20 μmol·L-1上下游引物、2 μL模板DNA(6~10 ng)以及7.6 μL滅菌超純水。將PCR產物回收,連接至pMD18-T載體(TaKaRa),轉化至大腸桿菌DH5α感受態中,篩選陽性克隆,提取各基因重組質粒,用NanoDrop 2000 分光光度計測定質粒濃度,計算各基因拷貝數,按10倍梯度稀釋備用,制作標準曲線。若標準曲線斜率在-3.5~-3.0范圍內,則繼續開展后續樣品測試。采用SYBR Green定量PCR法測定各基因拷貝數,熒光定量PCR反應在ABI7500熒光定量PCR儀(ABI)上進行,每個樣品設置3個平行樣,驗證檢測重復性及可靠性。
1.4 統計分析
采用SPSS 19.0軟件,對水體理化指標監測值及氮循環功能基因拷貝數在不同季節間的差異顯著性進行分析,并對不同功能基因間及功能基因與主要環境因子間進行Pearson相關性分析。
2 結果與討論
2.1 水環境質量現狀
2.1.1水質季節變化
參照GB 38382—2002《地表水環境質量標準》,由水體理化指標監測結果(表2)可知,射陽湖保護區冬、春季水體TN濃度符合Ⅴ類標準要求,秋季水體TN和TP濃度均符合Ⅳ類標準要求;夏季水體TP濃度達到(0.21±0.13) mg·L-1,若參照湖、庫標準,則水質為劣Ⅴ類,如果將射陽湖水域視為河網,則水質符合Ⅳ類標準要求。氮、磷濃度是決定該水域水質所屬類別的關鍵指標。冬季水體TN濃度最高,且顯著高于秋季和夏季(P<0.05);而夏季水體TP濃度最高,且顯著高于秋季和春季(P<0.05)。參考《湖泊(水庫)富營養化評價方法及分級技術規定》,射陽湖保護區4個季節綜合營養狀態指數(TLI)分別為53.5(秋)、54.9(冬)、59.8(春)和57.6(夏),存在輕度富營養化問題。
春季射陽湖保護區水體pH值和Chl a濃度顯著高于其他季節(P<0.05),說明浮游植物生物量較高。朱英等[11]指出夏季水溫較高,光照充足,而且暴雨徑流進入上海大蓮湖后,導致水體N、P濃度升高,從而誘發浮游植物生長,使得夏季部分湖區藻密度高于其他季節。筆者研究中夏季水體Chl a濃度低于春季,可能與射陽湖保護區夏季水體N/P比值較低[12]和水面浮葉植物生物量較高[13]有關。此外,春季小型硅藻大量繁殖[14]也可能是射陽湖保護區水體Chl a濃度在春季偏高的原因之一。
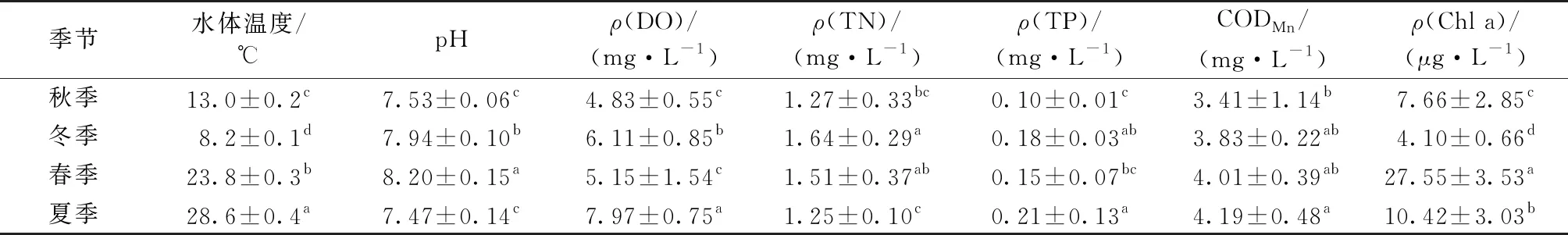
表2 調查水域不同季節水體理化指標均值
2.1.2水質空間變化
為探明射陽湖保護區氮、磷污染源頭,對水體NH4+-N、NOx--N(NO2--N和NO3--N濃度之和)和PO43--P濃度進行空間插值分析。由圖2可知,不同季節水體氮、磷濃度空間分布存在一定差異。秋季S3和S8站位水體ρ(NH4+-N)分別為0.89和0.68 mg·L-1,高于其他站位;由于兩個站位所處河流(走馬河)兩岸水產養殖池塘較多,養殖收獲季節清塘廢水排放[15]可能是局部水體NH4+-N濃度較高的主要原因。
冬季S5、S7和S11站位水體ρ(NH4+-N)分別為0.96、1.74和0.97 mg·L-1,高于其他站位;3個站位所處河流(大三王河)為射陽湖水域的主要河道,除了河道緊鄰的射陽湖鎮污水排放輸入外,枯水期上游NH4+-N污染輸入[16]可能也是導致保護區水域NH4+-N濃度升高的原因之一。春季S5和S6站位水體NH4+-N濃度較高,村鎮污水排放輸入[17]可能是主要原因。夏季S1、S6和S9站位(安豐河)水體NH4+-N濃度較高,同時S1和S6站位水體ρ(PO43--P)異常偏高,分別為0.27和0.35 mg·L-1,村鎮地表徑流輸入與上游河流輸入可能是該河段氮、磷濃度偏高的主要原因。在秋、冬季,NOx--N濃度均處于較低水平;而春季S2、S9和S11站位水體ρ(NOx--N)超過0.30 mg·L-1,夏季S8和S10站位水體ρ(NOx--N)分別為0.49和0.78 mg·L-1。上述點位大多位于射陽湖保護區核心區,春、夏季NOx--N濃度偏高可能與東側農業面源污染有關[18]。射陽湖保護區河網復雜,為從源頭上防治污染,需要進一步調查水體流向,并做好村鎮污染輸入截留與水產養殖轉型升級等工作,確保核心區水質逐年提升。
2.2 沉積物環境質量現狀
除了外源污染輸入外,底泥中內源污染釋放也可能造成上覆水體富營養化[19]。射陽湖水域4個季節底泥中全碳、全氮和全硫含量最高值大部分出現在S1和S3站位(圖3)。S1站位所處河段位于村鎮一側,底泥淤積嚴重,春、夏季底泥中全碳、全氮和全硫含量均高于其他站位。S3站位所處河段兩側分布有大量水產養殖池塘,冬季底泥中全碳、全氮含量和秋季底泥中全硫含量高于其他站位。村鎮污水排放和農業污染輸入可能分別是S1和S3站位底泥污染物積累的主要原因。除S1站位外,其他站位春、秋季底泥全氮含量均低于冬、夏季。此外,春季各站位底泥浸出液中NH4+-N濃度均值〔(7.49±1.89) mg·L-1〕顯著低于其他3個季節〔秋季,(16.56±6.09) mg·L-1;冬季,(14.70±2.28) mg·L-1;夏季,(12.91±3.57) mg·L-1〕。由于冬季水體溶解氧濃度較高,同時S2、S3和S4站位底泥中全氮含量處于4個季節最高水平,底泥中有機氮氧化引起的內源釋放可能是水體NH4+-N濃度升高的主要原因之一。在S3和S4站位(分布在農業種養殖區一側)4個季節底泥全硫含量變化不大,均處于較高水平;在S1、S2和S11站位(分布在射陽湖鎮一側),秋季底泥全硫含量明顯低于其他季節。特別是在S1站位,春、夏季底泥全硫含量明顯升高,且底泥全碳和全氮含量也處于較高水平,容易引發水體黑臭,需要進行河道清淤。
2.3 微生物群落氮循環功能基因
在不同形態氮素的相互轉化過程中,氨單加氧酶(AMO)、羥氨氧化還原酶(HAO)、亞硝酸鹽氧化還原酶(NXR)在硝化過程中發揮著重要作用,而在反硝化過程中硝酸還原酶(NAR)、亞硝酸還原酶(NIR)、一氧化氮還原酶(NOR)和氧化亞氮還原酶(NOS)可催化相關反應[20]。射陽湖保護區各站位底泥氮循環細菌功能基因(nosZ、nirS、nirK、narG、nxrA和AOB-amoA)拷貝數及其占細菌16S rDNA拷貝數比例的季節變化見圖4。S1站位春、秋季氮循環功能基因拷貝數及其占細菌16S rDNA拷貝數比例均高于冬、夏季,其中,春季S1站位各氮循環功能基因拷貝數總和遠高于其他季節和站位。冬季S3站位氮循環功能基因拷貝數雖然低于S2和S4站位,但氮循環功能基因占細菌16S rDNA比例在5個站位中最高。在春、冬兩季調查中,底泥全氮含量最高的站位分別是S1和S3,說明氮循環細菌功能基因拷貝數及其占細菌16S rDNA比例對底泥氮污染狀況具有一定指示作用。
春季各站位nosZ、narG和nirK基因豐度均較高,其中,nosZ和nirK基因豐度顯著高于其他季節(P<0.05)。同時,春季各站位nosZ基因相對拷貝數(nosZ/16S rDNA),以及nosZ基因與nirK和nirS基因之和(nir)的比值(nosZ/nir)均高于其他3個季節。春季除S1、S2站位外底泥全氮含量均低于其他季節,而浸出液NH4+-N濃度均值在4個季節中最低。較高的nosZ/nir比值說明在春季營養鹽水平相對較低的沉積物環境中,可能發生了完全反硝化過程[21]。相比于nirK基因,nirS基因豐度在4個季節均較高,其中,夏季各站位nirS基因豐度顯著高于春、秋季(P<0.05),僅在春季nirK/nirS基因豐度比值出現升高,說明nirS型反硝化菌在射陽湖保護區底泥反硝化過程中扮演重要角色。研究表明,與nirS型反硝化菌相比,含nirK基因的菌群適合在溶解氧濃度稍高、有機物濃度較低的環境生長[22]。在富營養化較為嚴重的太湖沉積物中nirS型反硝化菌是主要的生態類型[23],與之類似,射陽湖水域沉積物中的厭氧環境更適宜nirS型反硝化菌生長。
與上述反硝化功能基因相比,調查發現射陽湖水域沉積物中硝化功能基因(如nxrA和AOB-amoA)豐度相對較低。冬季nxrA和AOB-amoA基因豐度均值高于其他季節,其中,nxrA基因豐度顯著高于其他季節(P<0.05),可能與上覆水較高的NH4+-N濃度有關。研究表明,NH4+-N濃度較低的環境適宜AOA的生長,而在富營養化水域,AOB是驅動硝化作用的主要微生物類群[24]。筆者研究未調查AOA-amoA基因豐度,但在上覆水或底泥中NH4+-N濃度較低的季節,AOA可能也在硝化過程中發揮作用。
2.4 氮循環功能基因與主要環境因子的相關分析
通過將氮循環功能基因與主要環境因子進行相關分析(圖5),發現AOB-amoA基因豐度分別與nxrA和nirS基因豐度呈極顯著正相關,且nxrA基因豐度與nirS基因豐度呈顯著正相關。由3種功能基因編碼的AMO、NXR和NIR分別調控氨氧化、硝化和反硝化作用,依次將NH4+-N轉化為NO2--N,將NO2--N轉化為NO3--N,以及將NO2--N轉化為NO或NH3[20,25]。上述結果表明,傳統的硝化、反硝化路徑可能在射陽湖水域沉積物氮循環中占據重要地位。此外,narG基因豐度分別與nirK基因和nosZ基因豐度呈顯著正相關,且nirK基因豐度與nosZ基因豐度呈極顯著正相關,3種反硝化功能基因豐度均在春季出現明顯升高。李秋芬等[26]研究發現環境中同時存在含nirK和nosZ基因的反硝化菌,春季此類反硝化菌可能在射陽湖水域大量出現。
底泥浸出液中NH4+-N含量與nosZ基因豐度及上覆水pH值呈顯著負相關,與nosZ/nir和nirK/nirS呈極顯著負相關。春季多數站位較低的底泥全氮含量以及底泥浸出液NH4+-N濃度可能與nosZ與nirK基因豐度升高有關。此外,上覆水pH值與nirK和nosZ基因豐度,以及nosZ/nir和nirK/nirS比值呈極顯著正相關;同時,pH與Chl a濃度呈極顯著正相關。春季藻類大量繁殖引起上覆水pH升高,進而對底泥微生物群落氮循環菌具有一定影響。
底泥中全碳與全氮含量呈極顯著正相關,與上覆水CODMn濃度呈顯著正相關,表明上覆水中污染物的沉降是底泥有機污染的主要源頭,并引起內源性營養物質負荷增加[27]。上覆水NH4+-N濃度與水溫和Chl a濃度呈顯著負相關,而上覆水NOx--N濃度與水溫呈極顯著正相關。藻類一般優先利用污水中 NH4+-N 和其他還原態氮,它們對硝酸鹽的吸收利用僅僅發生在污水中 NH4+-N 濃度很低或耗盡的情況[28]。春、夏季水溫升高導致射陽湖水域Chl a濃度升高,藻類對不同形態氮素的選擇性吸收利用是引起上覆水中NH4+-N濃度降低的原因之一。
3 結論
射陽湖國家級水產種質資源保護區存在水體輕度富營養化問題,TN和TP是決定該水域所屬環境質量類別的關鍵指標。不同季節保護區水體氮、磷濃度的空間分布存在一定差異,射陽湖鎮污水排放和農業污染輸入可能是造成氮、磷濃度時空差異的主要原因。
射陽湖保護區4個季節底泥中全碳、全氮和全硫含量在S1(靠近村鎮)和S3(靠近農業區)站位較高。其中,S1站位春、夏季底泥全硫含量明顯升高,且底泥全碳和全氮含量也處于較高水平。
氮循環細菌功能基因拷貝數及其占細菌16S rDNA比例對底泥氮污染狀況具有一定指示作用。在射陽湖保護區底泥反硝化過程中nirS型反硝化菌占主導地位,另外,同時含nirK和nosZ基因的反硝化菌豐度可能在春季較高。
