干擾素-ε研究進展
2021-11-24 10:27:24張海玲廉士珍張東亮章沙沙劉方圓盧士英
畜牧獸醫學報 2021年11期
張海玲,廉士珍,張東亮,白 雪,章沙沙,張 蕾,李 偉,劉方圓,王 菡,胡 博,盧士英*
(1.吉林大學人獸共患病研究所 人獸共患病教育部重點實驗室,長春 130062; 2.中國農業科學院特產研究所 農業部特種經濟動物疫病重點實驗室,長春 130112;3.吉林特研生物技術有限責任公司,長春 130122)
干擾素(interferons,IFNs)是由脊椎動物細胞產生的一類分泌型糖蛋白[1-2],是由Issacs和Lindenmann 1957年在雞胚絨毛尿囊膜培養物中檢測流感病毒多重復制的干擾現象時發現[3]。干擾素應答是抵御病毒入侵的第一道防線[1,4-5],機體通過天然免疫識別病毒感染激活IFNs應答,IFNs與以自分泌和旁分泌方式表達而呈廣泛分布的干擾素受體(interferon receptor,IFNAR)結合,激活數百個干擾素刺激基因(interferon-stimulated genes,ISGs)參與的抗病毒防御程序,這些基因能夠干擾病毒復制的每一步[6-8],使機體快速進入一種抗病毒狀態,同時激活對病毒的適應性免疫應答[9-10]。干擾素的研究非常廣泛,在人及猴、小鼠、牛、羊、犬、豬、馬、大熊貓、兔、鼬等哺乳動物,以及野生動物禽類、龜類、魚類等都發現干擾素物質的存在[11-13]。根據干擾素的染色體定位、受體特異性和基因特征將其分為3種類型,Ⅰ型、Ⅱ型和Ⅲ型。Ⅰ型干擾素具有抗病毒、抗腫瘤作用[7, 14];Ⅱ型由單基編碼的IFN-γ構成,具有免疫調節作用[15-17]。Ⅲ型為IFN-λ,與Ⅰ型干擾素(IFN-α和IFN-β)的功能相似[18-20],此外,IFN-λ在黏膜屏障的先天抗病毒、抗真菌和抗原蟲感染中具有重要防御作用[20-22]。
Ⅰ型干擾素家族具有相同的受體類型和基因位點,迄今為止已確認有9類,分別是IFN-α、IFN-β、IFN-ω、IFN-κ、IFN-ε、IFN-δ、IFN-τ、IFN-ν和IFN-ζ[4, 23]。由于IFN-ε發現較晚,相關研究和報道較少,其研究還停留在初步階段,其很多性質和生物學作用尚不明確。IFN-ε作為Ⅰ型干擾素具有與IFN-α、IFN-β等其他Ⅰ型干擾素不同的、獨特的內源性表達模式,不需要病毒刺激或者干擾素誘導因子誘導,并在各個器官和組織黏膜上皮細胞中組成型表達,表達水平受雌激素調節[24-26]。其具有重要的抗病毒和黏膜免疫調節功能[27],對神經系統的有利調節也優于IFN-α/β[26]。盡管IFN-ε與IFN-α、IFN-β等其他Ⅰ型干擾素存在不同,但與其他類型一樣,可與IFNAR結合,激活JAK/STAT信號,產生抗病毒、抗增殖和免疫調節反應[28-29]。研究發現,IFN-ε在人和其他動物體內器官和組織的分布規律不同,不同動物基因序列差異顯著[30]。目前,研究僅證實人、小鼠、牛、羊和犬IFN-ε具有明顯的抗病毒作用[31-32],駱駝IFN-ε具有抗腫瘤作用[26],其他生物學功能需要開展進一步研究。
1 IFN-ε的起源
早在1981年,美國的Allan和Jarvis[33]在專利中描述其發現了一種新的具有抗病毒活性的物質——干擾素-ε,是將原代二倍體人上皮細胞接種病毒并分泌到培養液中產生,其分子量約20 ku,這種新物質與IFN-α、β和γ具有抗原特異性和種屬特異性,在人上皮細胞中具有明顯的抗病毒特性,但在其他類型的人細胞以及牛和小鼠細胞均未顯示抗病毒活性[32-33]。由此,發展了利用培養上皮細胞產生IFN-ε治療人上皮細胞病毒感染的方法。隨后,1984年由加拿大學者Roy和Pang[34]公開了一種定量檢測IFN-ε的方法,該方法將IFN-ε與人角質細胞和人成纖維細胞一起孵育,然后進行病毒攻擊,同時設立標準品,以此來確定抗病毒活性單位。直到1999年,由美國基因泰克公司首次公開了人IFN-ε基因序列[35]。研究發現,人IFN-ε在不同器官組織表達水平不同,人IFN-ε基因在胎盤組織、支氣管上皮細胞、微血管內皮細胞、動脈血管平滑肌細胞、卵巢、子宮、陰道、腦、小腸等組織中高表達[26, 36],并受雌激素調控。
2 IFN-ε在動物體內的分布規律
IFN-ε具有種屬特異性,在不同物種中,IFN-ε基因在各個器官、組織中表達水平也不盡相同。IFN-ε可在大部分動物生殖器官組織中高表達。包括獼猴、小鼠和雌性綿羊卵巢、子宮以及雄性睪丸組織中高水平表達,且隨著雌激素水平的升高,IFN-ε轉錄水平升高[28]。其次,在獼猴、豬和牛的胸腺、小腸、腸系膜淋巴結、骨髓和皮膚中也檢測到IFN-ε表達[31,37-38]。在獼猴和牛的腎,豬、牛和綿羊的肝也能檢測到表達[33]。由于IFN-ε處于研究的初步階段,研究人員選取檢測器官組織類別不一致會導致IFN-ε在不同組織中的分布規律差異,特別是脾、淋巴結、骨髓、肌肉、膀胱和氣管檢測數據較少,想要明確IFN-ε的分布規律還需要開展完善的試驗檢測。
3 IFN-ε表達與黏膜免疫的關系
IFN-ε是一種獨特的Ⅰ型IFN,不受模式識別反應元件的誘導。在女性體內,IFN-ε基因在子宮、宮頸、陰道和卵巢組織高表達。在正常生理狀態下,IFN-ε可增強女性生殖道(female reproductive tract,FRT)黏膜免疫,具有保護女性生殖系統不受病毒和細菌感染的重要作用[24, 28-39]。推測IFN-ε從上皮細胞分泌到FRT管腔和間質中,可作用于應答細胞,但其分布的確切機制和模式尚待闡明。IFN-ε的表達受雌激素和孕激素調節,分別促進或抑制IFN-ε的表達[24]。IFN-ε通過免疫細胞和上皮細胞表面的Ⅰ型干擾素受體(IFNAR)信號上調干擾素刺激基因(ISGs)(圖1),激活細胞對IFN-ε的特異性應答,增加免疫細胞的數量和活性,上調上皮細胞的抗病毒和抗菌免疫應答。
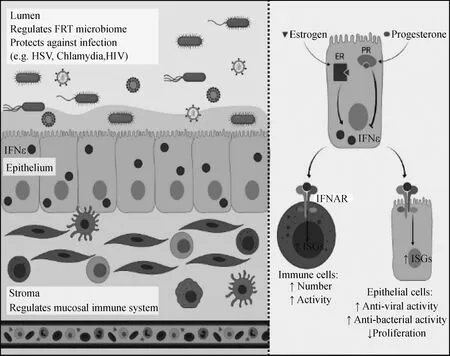
左:IFN-ε由FRT上皮細胞組成型表達,在此部位有助于局部維持穩態和黏膜免疫。右:在FRT上部,IFN-ε的表達受雌激素和孕激素的激素調節,分別促進或抑制IFN-ε的表達
有研究顯示,IFN-ε也可能是女性新冠病毒患者死亡率低的原因之一。通過對COVID-19感染死亡人數數據分析發現,男性死亡率高于女性[40-43]。土耳其巴爾基西爾大學醫學研究人員在COVID-19感染治療研究中發現,IFN-ε抗病毒效果降低了COVID-19女性感染者的致死率[44]。分析認為,IFN-ε與其他類型的Ⅰ型IFNs一樣,在抵抗病毒感染時具備基本自然免疫防御反應,同時女性受雌激素水平影響又促進了IFN-ε在體內表達,這可能是降低女性新冠病毒患者死亡率的原因。另一份報告認為,男性死亡率較高與冠狀病毒功能性受體血管緊張素轉化酶2(angiotensin-converting enzyme 2,ACE2)水平較高有關,ACE2受體在睪丸細胞中大量表達,且其能夠幫助SARS-CoV-2進入宿主細胞,故較高的ACE2濃度可能導致患者對SARS-CoV-2的易感性增加[45-46]。其他IFNs,如IFN-λ可減輕病毒載量,改善患者臨床癥狀,是治療COVID-19的理想藥物,但對死亡率無影響[47]。此外,IFN-ε還保護女性生殖道免受細菌感染,在月經周期、生殖和更年期受激素調節,并受外源性激素調節[24, 28]。
對穿山甲免疫系分析發現,IFN-ε具有相似的黏膜免疫保護作用。穿山甲被認為是SARS-CoV-2樣冠狀病毒的天然蓄水池[45],其免疫系統的進化與其他哺乳動物不同,非洲、亞洲穿山甲的皮膚和黏膜上皮細胞(肺、腸和生殖組織)高水平表達IFN-ε,對皮膚和黏膜免疫具有重要防御作用[28, 46]。這可能是穿山甲在冠狀病毒感中呈現輕微癥狀的原因之一。
4 IFN-ε對神經系統的調節作用
IFN-α是一種多效性細胞因子,應用于臨床治療已有30多年,許多干擾素治療的副作用也相繼被報道[48-49],特別是對神經系統的不良影響。IFN受體存在于中樞神經系統中,臨床上使用的IFN與這些受體結合,可引起癡呆、抑郁、厭食、運動神經系統抑制等癥狀[49-53]。此外,IFN-α干擾神經營養信號,阻礙神經突起生長、突觸可塑性、內源性神經發生和神經元存活[54]。與其他干擾素不同,IFN-ε的一個特點是在腦內呈組成型高表達,提示IFN-ε可能對大腦和中樞神經系統有特殊影響。對與大腦和神經系統相關的IFN-ε調節基因進行分析發現,這些基因參與許多生理功能,如維持大腦結構、神經細胞的分化、興奮調節等。例如,rhIFN-ε可促進中樞神經系統鈣調蛋白結合蛋白striatin與細胞內Ca2 +結合,調節神經興奮性突觸的功能,而IFN-α沒有此功能。rhIFN-ε也可調控與神經細胞結構相關的軸周蛋白(Periaxin,PRX)基因的的表達,影響神經鞘磷脂的形成和神經細胞正常結構的維持[26]。IFN-ε調控的透明質酸介導的運動受體被腦微血管內皮細胞高表達,可能參與腦微血管的形成[55]。最后,IFN-ε激活了胰島淀粉樣多肽的表達,在非胰島素依賴型糖尿病和阿爾茨海默病神經元損傷的病理過程中發揮作用[26]。IFN-ε由于其持續表達于血管細胞表面,抑制細胞增殖,有望用于動脈粥樣硬化癥的治療。此外,經IFN-ε作用后,細胞產生有利于神經細胞的調節因子,可用于治療中樞神經系統的異常,包括焦慮、抑郁、神經分裂癥、帕金森氏癥等疾病。由于目前臨床應用治療的干擾素(IFN-α、IFN-β)普遍會產生神經抑郁的副反應,因此,IFN-ε有望成為對干擾素治療產生不良反應嚴重患者的替代藥品。
5 IFN-ε的抗病毒作用
近期的研究發現,利用基因工程技術體外表達的動物IFN-ε具有明顯的抗病毒活性。在同源和異源動物體外細胞上,IFN-ε顯示交叉保護抑制病毒復制的跨物種特性[56]。例如,應用大腸桿菌表達重組牛IFN-ε能夠抑制水泡性口炎病毒(vesicular stomatitis virus,VSV)在牛腎細胞(Madin-Darby bovine kidney,MDBK)、牛鼻甲細胞(bovine turbinate cells,BT)、胎牛腎細胞(embryonic bovine kidney cells,EBK)和豬腎細胞(porcine kidney 15 cell,PK-15)上的復制[56]。重組綿羊IFN-ε在體外可誘導ISG15、黏病毒抗性蛋白1(myxovirus resistanceproteinA,Mx1)和2’-5′寡聚腺苷酸合成酶(2’-5′ oligoadenylate synthetase,OAS)的轉錄,產生抗病毒蛋白抑制外周血淋巴細胞和MDBK、RK-13、F81、PK-15、BT細胞上的病毒復制,而在MDCK、BHK-21細胞無抗病毒活性[57]。
IFN-ε也可作用于HIV感染的早期階段,包括病毒進入、反轉錄和核導入。在人巨噬細胞中誘導一種阻斷HIV-1復制的抗病毒狀態。IFN-ε刺激的免疫介質和途徑具有Ⅰ型IFN的特征,但與巨噬細胞中的IFN-α不同。IFN-ε誘導了大量的吞噬和活性氧(reactive oxygen species,ROS)殺滅作用,從而阻礙了HIV的復制[58]。除此之外,IFN-ε具有抗增殖的作用,重組犬IFN-ε可激活JAK-STAT信號通路,具有顯著的抗增殖作用[59]。有學者通過噻唑藍(3-(4,5)-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide,MTT)細胞增殖檢測、形態學觀察和細胞凋亡試驗發現,包涵體表達的重組駱駝IFN-ε以劑量依賴性抑制乳腺癌細胞MDA-MB-231和MCF-7生長,與對照組相比,IFN-ε處理過的MDA-MB-231細胞Caspase-3的表達量明顯升高[60]。
Ⅰ型IFN都具有結構上的同源性,并與由IFNAR1 和IFNAR2亞基構成的異二聚體受體結合,該類受體在大多數細胞表面均有表達[61]。IFN受體兩個亞基IFNAR1和IFNAR2存在兩個獨立的配體結合位點,這種雙重結構保證了不同類型IFN被識別并結合啟動干擾素信號轉導[62]。IFN-ε與IFNAR結合,激活ISGs的表達,進而激活多種抗病毒蛋白表達抑制病毒復制。但是,在體內和體外試驗中,病毒誘導不能產生IFN-ε[63],而IFN-ε在生殖系統黏膜和支氣管黏膜上皮細胞組成型高表達,這些黏膜部位存在著不同的解剖結構、生理作用和微生物的差異,因此,要明確IFN-ε對不同病原體在不同黏膜組織內的保護作用需要進一步深入研究其作用機理。在女性生殖系統中IFN-ε表達水平受雌激素和孕激素的調節,是否存在其它的特異性作用機制調節IFN-ε的表達也有待進一步證實。
6 展 望
干擾素是國內外第一個被批準投放市場用于臨床抗病毒、抗腫瘤治療的基因工程藥物,其治療的適應癥有數十種[64-65],目前,也用于治療COVID-19感染的臨床試驗[44, 66-67]。而IFN-ε作為Ⅰ型IFNs中新發現的一個獨特家族成員,在多種細胞上具有明顯的抗病毒作用,其在黏膜免疫和對神經調節的作用明顯優于其它Ⅰ型IFNs。IFN-ε正在被研究用于由HPV感染引起的尖銳濕疣、尋常疣、喉乳頭狀瘤、宮頸癌等多種疾病,女性泌尿生殖系統感染以及部分腫瘤的輔助治療,有望成為其他治療用干擾素不良反應的替代品。目前的研究表明,病毒感染不能誘導機體和體外細胞產生IFN-ε,盡管IFN-ε對生殖道、腸道和肺部氣管黏膜有預防微生物感染的保護作用,這種保護作用可能通過某種組織特異性的機制進行調節,例如在雌性生殖系統通過分泌雌激素和孕激素調節IFN-ε的表達。其他黏膜部位的IFN-ε表達是通過什么機制調節的?是否存在依賴于細胞或特定組織中表達的其他輔助因子?IFN-ε對不同病原體在不同黏膜組織內保護作用的機制、廣度和潛力都需要進一步探索。
