利用數字圖像分析132 份胡枝子種子表型性狀遺傳多樣性
2021-11-22 12:29:20楊正禹陸忠杰張茂董瑞
草業學報 2021年11期
關鍵詞:分析
楊正禹,陸忠杰,張茂,董瑞
(貴州大學動物科學學院,貴州 貴陽550025)
胡枝子屬(Lespedeza)為豆科(Leguminosae)多年生草本、灌木或半灌木,全世界共有本屬植物100 余種,我國約65 種,在全國各地廣泛分布[1?2]。其具有蛋白質含量高、適口性好、抗逆性及水土保持能力強等優點[3?5],富含生物堿、黃酮、萜類、縮合單寧等醫用化學物質,其中縮合單寧能夠促進反芻動物的消化[6?8]。胡枝子(L. bicolor)還是中國草種子出口量較多的優良飼草之一,每年出口量達50~80 t[9]。其在畜牧業、醫藥及水土保持方面都得到了廣泛的應用。
種質資源是育種工作和物種開發的基礎,豐富的種質資源對于物種的收集、保存及改良具有重要意義[10]。物種的表型性狀多樣性是種質資源遺傳變異的外在表現,受環境及遺傳綜合作用,是最基礎、直接和便于掌握的生物學特性[11?13],也是植物遺傳多樣性研究及衡量物種多樣性的重要指標和主要內容,能直觀反映植物多樣性[14]。前人對同一物種不同種質性狀存在的變異進行了廣泛深入研究,如紅砂(Reaumuria soongorica)[15]、箭筈豌豆(Vicia sativa)[16]、大豆(Glycine max)[17]、苜蓿(Medicago sativa)[18]、小麥(Triticum aestivum)[19]等。
植物種子與其他器官相比,所受的外部壓力影響更小,形態更穩定,具有易收集、儲存時間長等多種優點,同時對幼苗的生長發育也有很大影響[20?22]。種子的壽命、萌發等也與種子的形態特征存在著密切的聯系[23]。因此,種子形態特征在分類學、物種遺傳變異等方面均具有重要意義,特別是在物種的鑒定和區分方面[24?25]。Apuan等[26]使用幾何形態計量學(geometric morphology,GM)工具進行圖像分析,將雜草稻與栽培水稻(Oryza sativa)種子性狀進行比較,確定了雜草稻與栽培稻之間的關聯和關系。童躍偉等[27]利用種子的數字圖像分析了6 個種群紅松(Pinus koraiensis)種子表型分化程度和變異規律。
本研究對132 份胡枝子屬種質的種子形態學特征、基因型變異進行分析,旨在對胡枝子的種子表型性狀進行全面了解,為胡枝子遺傳改良提供現實依據。
1 材料與方法
1. 1 試驗材料
試驗材料為132 份胡枝子屬種質種子(表1)。其中131 份種質由美國國家植物種質系統(National Plant Germplasm System of the United States,NPGS)提供,1 份為中國野生種。

表1 胡枝子種質信息表Table 1 Number distribution of Lespedeza germplasm
1. 2 試驗方法
本研究于2019 年7 月進行,分別對L. juncea(7份)、L. cuneata(42 份)、L. capitata(51 份)、L. bicolor(10 份)4 個種及全部132 份胡枝子種質進行分析。共測量7 個種子性狀,分別為種子長(seed length,SL)、種子寬(seed width,SW)、長寬比(width to length ratio,SL/SW)、種子周長(perimeter,Pe)、種子厚(seed thick,ST)、種臍長(hilum length,HL)、百粒重(100-seed weight,SY)。百粒重為100 粒種子重量,其余性狀每份種質測量30 粒種子,3 次重復。
利用掃描儀(EPSON,印度尼西亞)對每份種質種子(每份種子30 粒,3 次重復)進行掃描,獲得種質的種子圖片,圖像分辨率為300 dpi,像素為1024×1024。使用WinSEEDLETM 2011 圖像分析系統對種子圖像進行分析,得到種子長(SL)、種子寬(SW)、長寬比(SL/SW)、種子周長(Pe)。使用數顯游標卡尺(AIRAI SR44,精度0. 01 mm,山東青島)測量種子厚度(ST)、種臍長度(HL),電子天平(Mettler Toledo-MS204S,精度0. 0000 g,瑞士)稱量種子百粒重(SY)。
1. 3 數據分析
數據分析基于:1)利用方差分析來評估胡枝子種質間各表型性狀基因型變異的顯著性和大小。2)利用模式分析,包含聚類和主成分分析(principal component analysis,PCA)[28],以圖形方式來展現所有種質各性狀數據矩陣。
在GenStat 7. 1(2003)中使用殘差最大似然(residual maximum likelihood,REML)進行方差分量分析。隨機線性模型用于分析胡枝子種質間及種間性狀變異。利用最佳線性無偏估計值(best linear unbiased predictors,BLUP)[29]分析,來獲得各種質種子的性狀均值,再利用均值進行主成分和聚類分析。其中方向向量表示各性狀之間的相關性。還進行了以胡枝子不同種質和種作為固定效應的混合線性模型分析,以研究胡枝子不同種質和種之間的性狀差異。
使用REML 分析中的基因型(σg2)和實驗誤差(σε2)的方差分量以及nr(重復次數)估算每個種質各性狀的平均重復性(R)[30]。用變異系數(CV)表示各性狀離散程度。

式中:S為標準差,X為平均值。
各種子性狀之間的表型相關性(rp)使用SPSS 20. 0 分析。
2 結果與分析
2. 1 表型性狀多樣性分析
對胡枝子屬不同種的種子表型性狀分析發現,性狀SL、SW、ST、HL、SY、Pe 在L. bicolor中最大,分別為3. 156 mm、2. 469 mm、1. 562 mm、0. 697 mm、0. 558 g 和8. 950 mm。性狀SL、SW、SY/SW、HL、SY 和Pe 在L.cuneata中最小,分別為1. 747 mm、1. 414 mm、1. 238 mm、0. 562 mm、0. 163 g 和4. 880 mm(表2)。 表明L.bicolor的種子最大,而L. cuneata的種子最小。132 份胡枝子種質間各性狀均存在顯著(P<0. 05)差異。在L.juncea、L. cuneata、L. capitata和L. bicolor4 個種中,除SL/SW 外,其余性狀均具有顯著(P<0. 05)差異,表明胡枝子種子表型性狀存在豐富的基因型變異。所有種質中除SL/SW 外,其余性狀的CV 均較高,其中SY 高達61. 53%。在L. juncea、L. cuneata和L. bicolor中,SY 的CV 最高,分別為61. 95%、20. 60% 和40. 63%。上述結果表明胡枝子種間及種內的種子表型性狀多樣性均較高。
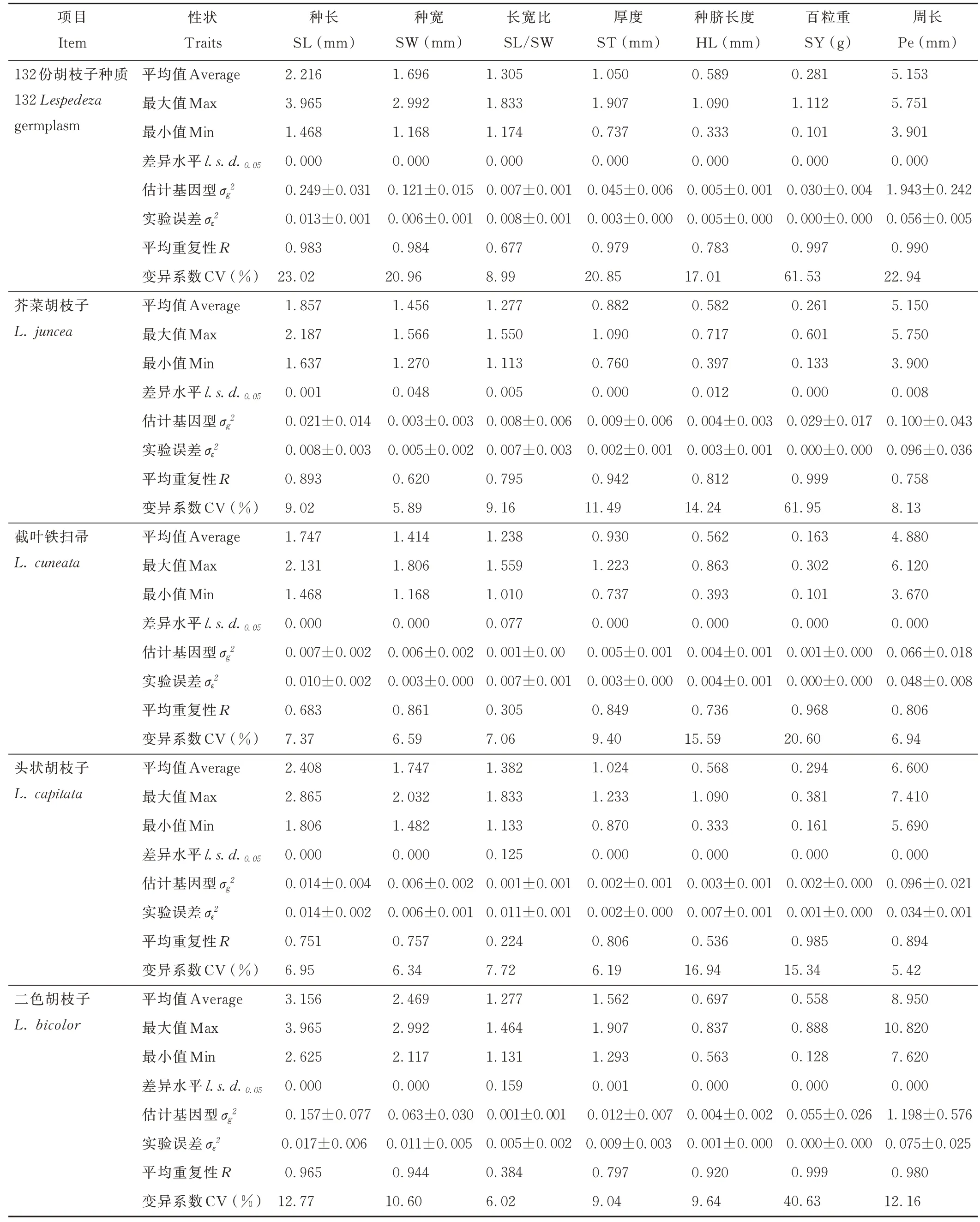
表2 胡枝子種質估計基因型、實驗誤差方差分量及相關標準誤差評估Table 2 Estimated genotype,experimental error variance component and related standard error evaluation of Lespedeza germplasm
在132 份胡枝子種質間,性狀SL、SW、ST、SY 和Pe 具有高的性狀平均重復性(R)值,分別為0. 983、0. 984、0. 979、0. 997 和0. 990,性狀SL/SW 和HL 具有中等R值,分別為0. 677 和0. 783。在L. juncea中,性狀ST、SY 具有高R值,分別為0. 942 和0. 999。在L. cuneata和L. capitata中,僅SY 具有高R值,分別為0. 968 和0. 985。在L. bicolor中,SL、SW、HL、SY 和Pe 具有高R值,分別為0. 965、0. 944、0. 920、0. 999 和0. 980。所有種中性狀SY均有高的R值,表明性狀SY 遺傳性穩定,不易受環境影響,可作為品種選育的重要參考性狀。
2. 2 種子形態特征模式分析及表型相關性
在132 份胡枝子的7 個性狀中(圖1),性狀ST、SW、SY、SL 和Pe 之間呈正相關(方向向量間角度<90°),且SW 與SY 呈強正相關。而性狀SL/SW與HL、ST 呈負相關(方向向量間角度>90°)。主成分分析顯示,主成分1 貢獻率為68. 85%,主成分2 貢獻率為15. 95%,累計達84. 80%。
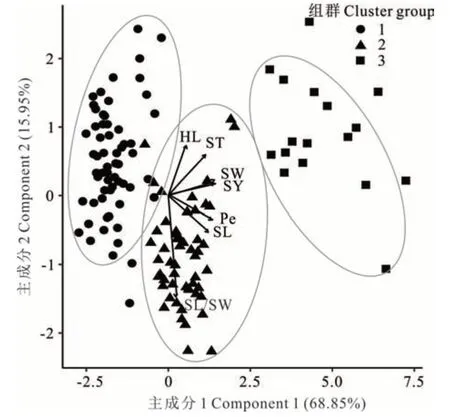
圖1 132 份胡枝子種質7 個性狀的主成分分析及聚類分析Fig. 1 Principle component analysis and cluster analysis for seven traits of 132 Lespedeza germplasm
聚類分析將132 份胡枝子種質分為3 個組群(表3),第1 組包含63 份種質,第2 組包含52 份種質,第3組包含17 份種質。性狀SL、SW、ST、SY 和Pe 在第3組中存在最大值,性狀SL/SW 在第2 組中存在最大值,7 個性狀均在第1 組中存在最小值。
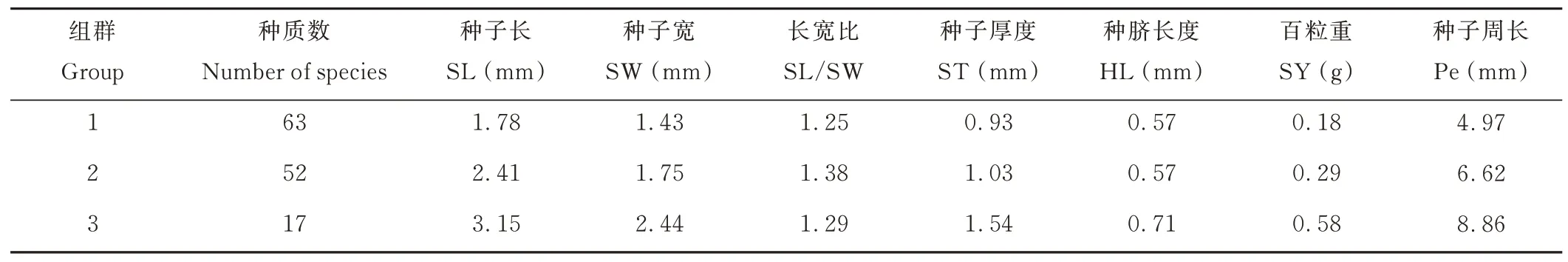
表3 132 份胡枝子種質聚類分析分組信息Table 3 Cluster analysis grouping information of 132 Lespedeza accessions
表4 中132 份胡枝子種質各性狀間的表型相關性(rp)系數進一步證明了圖1 中ST、SW、SY、SL 和Pe之間呈正相關,且差異極顯著(P<0. 01)。正rp的最小值在SL/SW 與SY 間,為0. 058。 最大值在SL 與Pe 間,為0. 991。
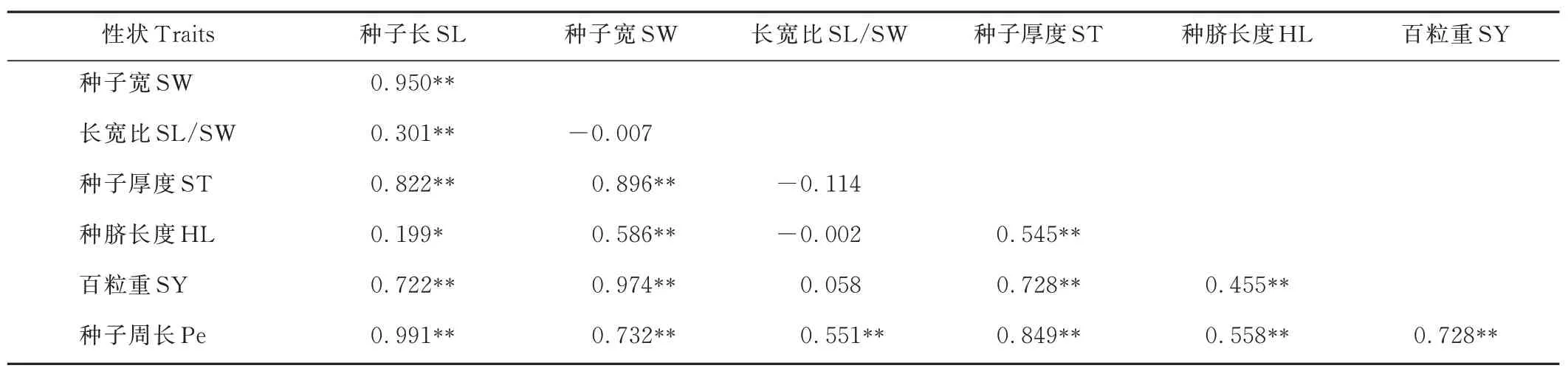
表4 132 份胡枝子種質各性狀間表型相關性系數Table 4 Phenotypic(rp)correlation coefficients among traits based on the 132 Lespedeza accessions
在L. juncea中(圖2),性狀ST 與HL,Pe 與SL 均呈強正相關。SY 與ST、HL 呈正相關(方向向量間角度<90°)。性狀SW 與SL/SW,SL/SW 與Pe、SL 均呈負相關(方向向量間角度>90°)。主成分分析顯示,主成分1 貢獻率為43. 25%,主成分2 貢獻率為36. 57%,累計達79. 82%。
對L. juncea進行聚類分析,可將其分為2 個組群(表5)。第1 組包含3 個種質,第2 組包含4 個種質。第1 組各性狀均值除ST 外,其余均大于第2 組。
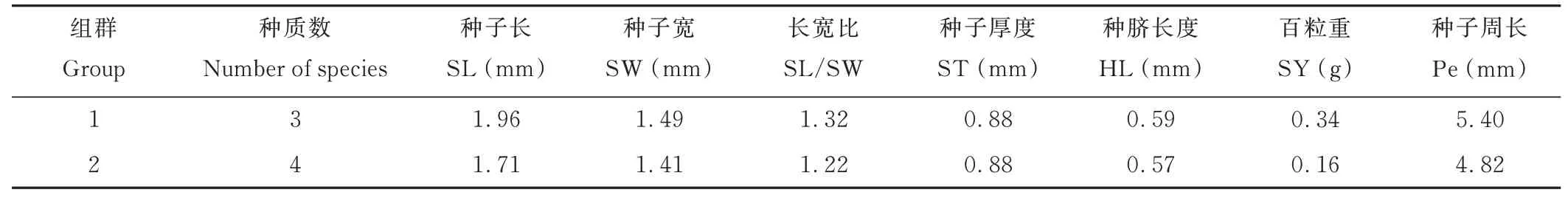
表5 L. juncea 聚類分析分組信息Table 5 Cluster analysis grouping information of L. juncea
相關性系數(rp)進一步驗證了圖2 所示的性狀Pe與SL,ST 與HL,SW 與Pe 均在顯著(P<0. 05)水平下呈正相關(表6)。正rp的最小值在SW 與SY 間,為0. 082,最大值在SL 與Pe 間,為0. 871。
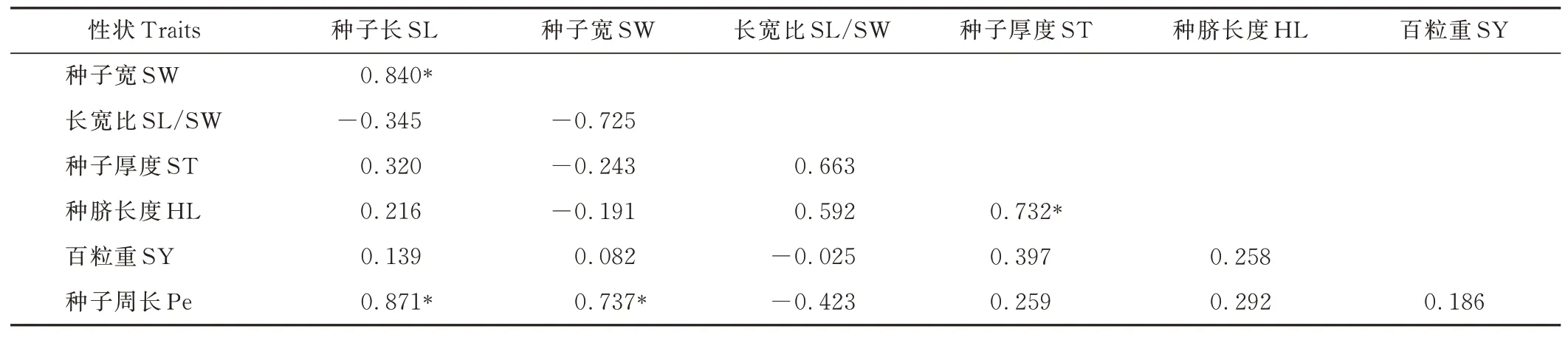
表6 7 份L. juncea 種質各性狀之間的表型相關性系數Table 6 Phenotypic(rp)correlation coefficients among traits based on the 7 L. juncea accessions
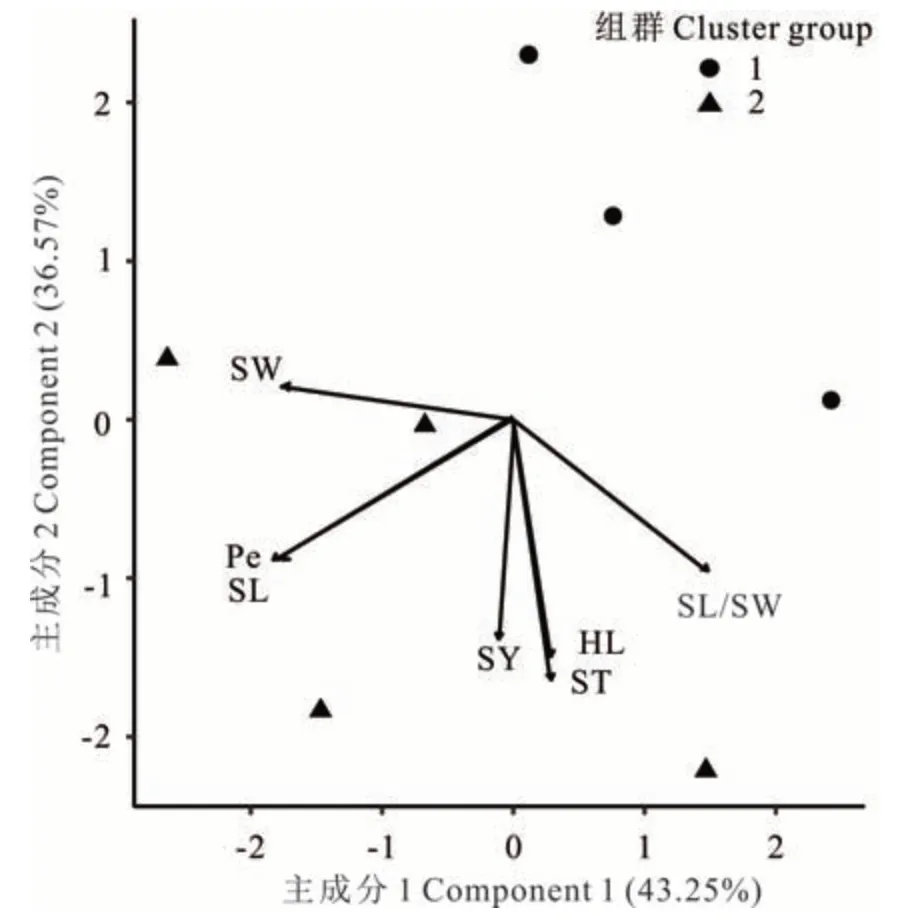
圖2 L. juncea 7 個性狀主成分分析及聚類分析Fig. 2 Principle component analysis and cluster analysis for seven traits of L. juncea
在L. cuneata中(圖3),性狀HL 與SL/SW,ST與SY 間均呈強正相關(方向向量間角度<90°),而性狀SW 與HL、SL/SW、SY、ST 呈負相關。主成分分析結果顯示,主成分1 貢獻率為42. 94%,主成分2 貢獻率為23. 69%,累計達66. 63%。
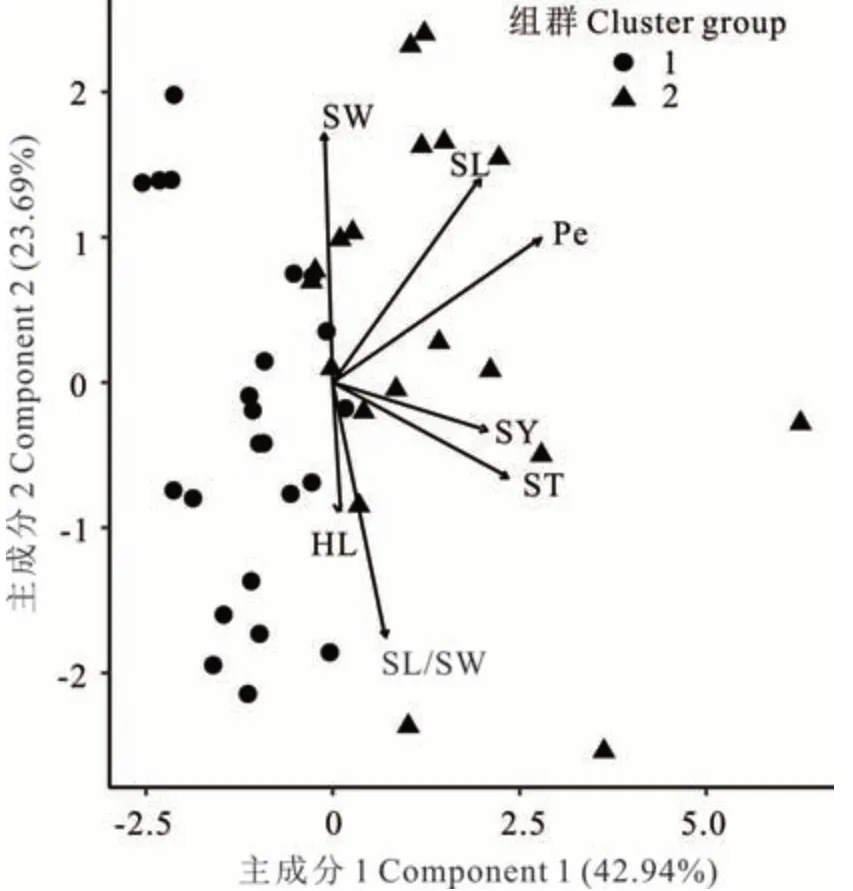
圖3 L. cuneata 7 個性狀的主成分分析及聚類分析Fig. 3 Principle component analysis and cluster analysis for seven traits of L. cuneata
聚類分析將42 份L. cuneata分為2 個組群(表7),第1 組包含23 個種質,第2 組包含19 個種質。由性狀均值可看出,第2 組除性狀SY 外,其余各性狀均值均大于第1 組。
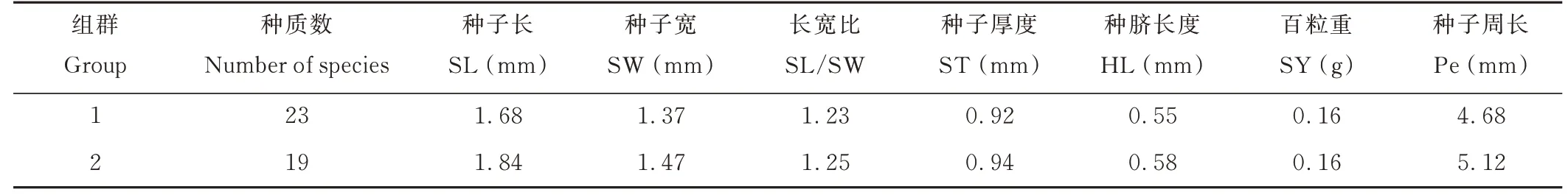
表7 L. cuneata 聚類分析信息Table 7 Cluster analysis grouping information of L. cuneata
L. cuneata各性狀間表型(rp)相關性系數進一步證明了性狀ST 與SY、HL 與SL/SW 均在極顯著(P<0. 05)水平下呈正相關(表8)。性狀Pe 與SL、ST 之間也存在極顯著(P<0. 01)正相關。HL 與SL/SW 間rp最大,為0. 893,最小rp在HL 與ST 之間,為0. 002。
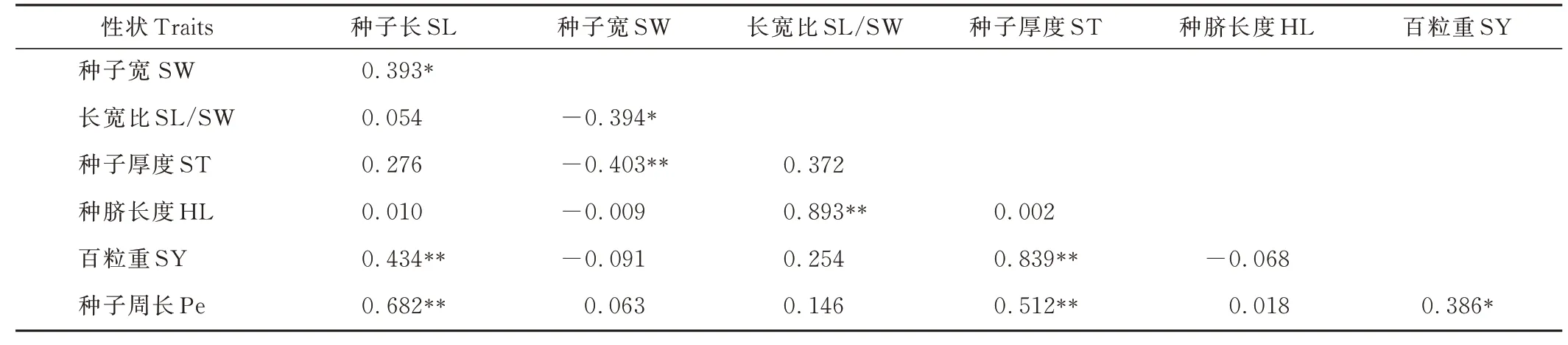
表8 42 份L. cuneata 種質各性狀之間的表型相關性系數Table 8 Phenotypic(rp)correlation coefficients among traits based on the 42 L. cuneata accessions
在L. capitata中(圖4),性狀SY 與SW 呈強正相關(方向向量間角度<90°),HL 與SY、SW 間呈正相關。性狀ST 與SY、SW、HL 呈負相關(方向向量間角度>90°)。主成分分析顯示,主成分1 貢獻率為43. 98%,主成分2貢獻率為23. 63%,累計達67. 61%。
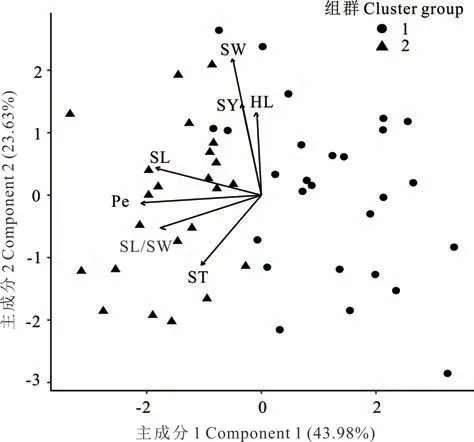
圖4 L. capitata 7 個性狀的主成分分析及聚類分析Fig. 4 Principle component analysis and cluster analysis for seven traits of L. capitata
聚類分析將51 份L. capitata分為2 個組群(表9)。第1 組包含28 份種質,第2 組包含23 份種質。性狀HL 與SY 在2 個組群中的均值相等,第1 組除性狀SL/SW 外,其余性狀均值均小于第2 組。
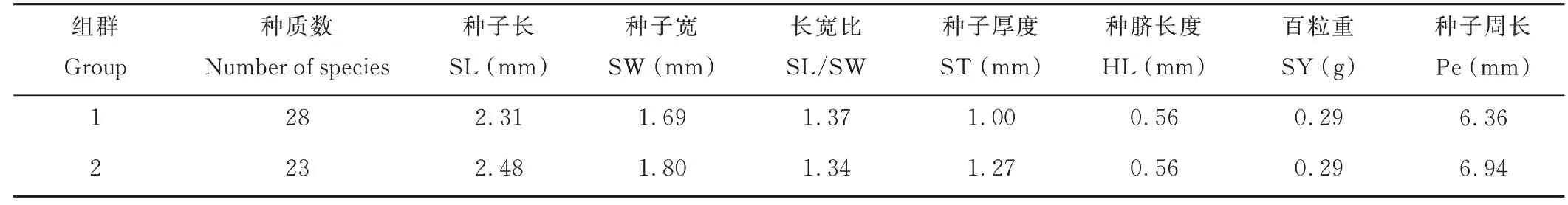
表9 L. capitata 聚類分析信息Table 9 Cluster analysis grouping information of L. capitata
L. capitata各性狀之間的表型(rp)相關性系數進一步證明了SY 與SW 在極顯著(P<0. 05)水平下正相關(表10)。 性狀SL 與SW、SL/SW、Pe,SW 與HL、SY,SL/SW 與Pe,ST 與Pe 間均存在極顯著(P<0. 01)正相關。 性狀SW 與ST 在極顯著(P<0. 01)水平下呈負相關。正rp最小值在SL/SW 與SY之間,為0. 050,最大值在SY 與SW 之間,為0. 945。
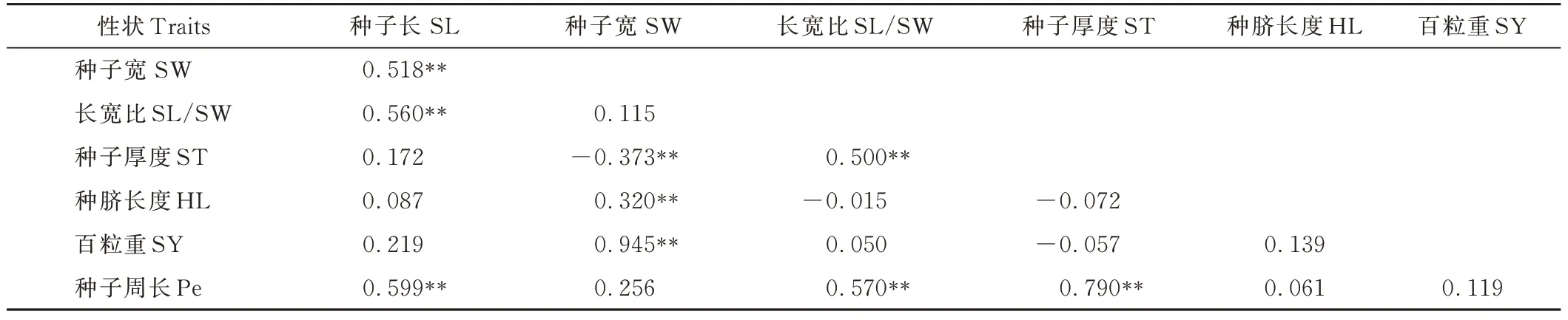
表10 51 份L. capitata 種質各性狀之間的表型相關性系數Table 10 Phenotypic(rp)correlation coefficients among traits based on the 51 L. capitata accessions
在L. bicolor中(圖5),性狀SL、SL/SW、Pe、ST 和HL 之間呈正相關(方向向量間角度<90°)。性狀SY 與SW、SL/SW 呈負相關(方向向量間角度>90°)。主成分分析顯示,主成分1 貢獻率為60. 53%,主成分2 貢獻率為19. 80%,累計達80. 33%。
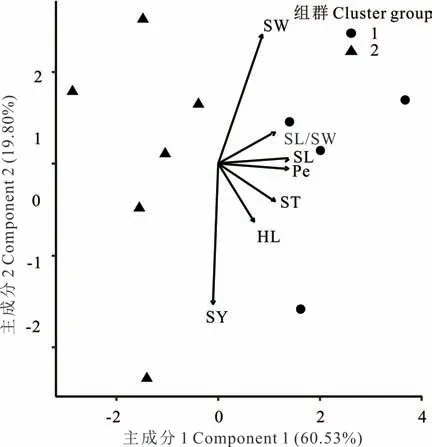
圖5 L. bicolor 7 個性狀的主成分分析及聚類分析Fig. 5 Principle component analysis and cluster analysis for seven traits of L. bicolor
聚類分析將10 份L. bicolor分為2 個組群(表11)。第1 組包含4 份種質,第2 組包含6 份種質。兩個組群的各性狀均值除HL 相等外,其余性狀均值第2組均大于第1 組。
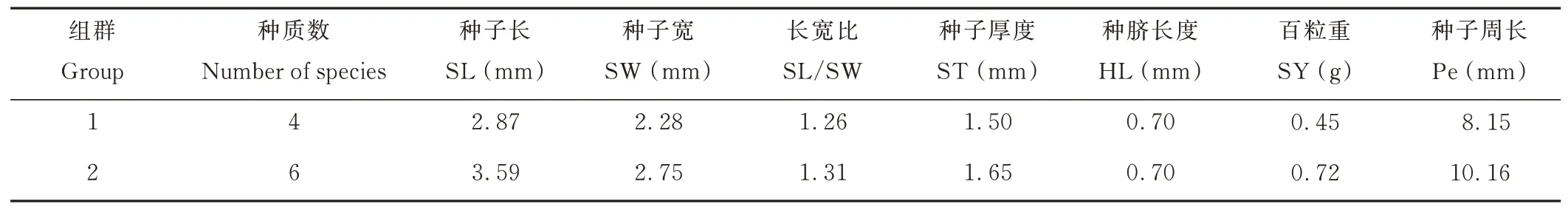
表11 L. bicolor 聚類分析信息Table 11 Cluster analysis grouping information of L. bicolor
L. bicolor各性狀之間的表型(rp)相關性系數進一步證明了性狀Pe 與ST、SL,SL 與ST 在極顯著(P<0. 01)水平下呈正相關,SL/SW 與HL、SL 與SW 在顯著(P<0. 05)水平下呈正相關(表12)。正rp的最小值在SL 與SY之間,為0. 049,最大值在SL 與Pe 之間,為0. 997。
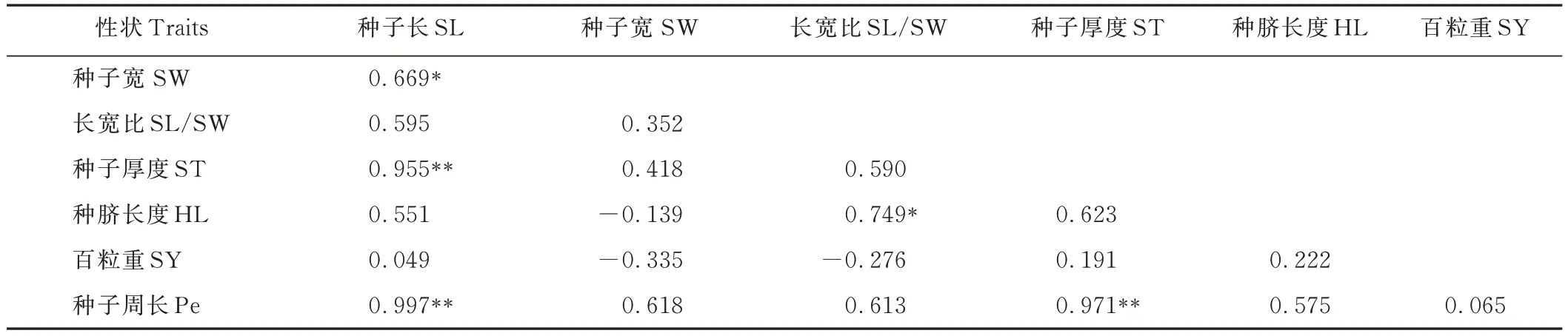
表12 10 份L. bicolor 種質各性狀之間的表型相關性系數Table 12 Phenotypic(rp)correlation coefficients among traits based on the 10 L. bicolor accessions
3 討論
自然資源的遺傳變異是經過長期自然選擇形成,從而獲得了相對穩定且豐富的表型變異特征[31]。本研究對胡枝子屬132 份種質及4 個種L. juncea、L. capitata、L. bicolor和L. cuneata的7 個種子性狀進行分析。結果表明,除性狀SL/SW 外,其余種子性狀都存在顯著(P<0. 05)的基因型變異,這些種子性狀的基因型變異和R值表明,在各種質內存在潛在可用的遺傳變異。張吉宇[32]對14 個胡枝子屬野生居群植株形態特征進行了遺傳多樣性研究,發現胡枝子的表型性狀豐富。Bécquer 等[33]和Ocampo 等[25]也從不同植物種子數字圖像中提取了長度、寬度、周長以及種子形狀等數據來分析其遺傳多樣性和物種分類。本研究中各種子表型性狀的CV 均較高(>5%)。有研究表明,性狀的CV 超過5% 可視為產生了較大變異[34],而CV 越大說明植物對于環境的適應能力越強,能適應的環境范圍也越廣[35]。本研究中132 份胡枝子種質CV 均較高,這是由于132 份胡枝子種質來自全球不同地區,經過了長期的地理隔離及不同海拔和環境氣候的影響。而環境條件是種質資源遺傳變異的主要因素之一。
根據R值可判斷各表型性狀的遺傳穩定性。L. juncea中性狀SL、SL/SW、Pe、HL、SY 和ST 具有較高(R>0. 7)的R值;L. cuneata中性狀SW、Pe、HL、ST 和SY 具有較高的R值;L. capitata中性狀SL、SW、Pe、SY 和ST具有較高的R值;L. bicolor中性狀SL、SW、Pe、HL、SY 和ST 具有較高的R值。性狀的R值越高表明其受遺傳因素的影響越大,越不易受環境因素影響,遺傳更具穩定性。L. bicolor各性狀的R值均高于其他種,表明L.bicolor與其他種相比遺傳更穩定。在4 個種中,性狀SY、ST、Pe 的R值均較高(R>0. 9),相比其他性狀,其遺傳穩定性較好,不易被環境因素所改變,能夠在后期的繁育過程中穩定遺傳,保持優良性狀。
種子表型相關性分析表明,性狀SL/SW 與SY、Pe 之間相關性較弱,而種子的SL/SW 主要用來描述種子的形狀,與種子的大小性狀無關,因此胡枝子種子大小與種子形狀獨立遺傳,這與對青稞(Hordeum vulgare)[36]、小麥[37]的研究結果相似。主成分分析結果表明,在132 份胡枝子種質中,第1 主成分解釋了總變異的68. 85%,第2主成分解釋了15. 95%,累計達84. 80%,說明這兩個主成分可反映大部分信息。第1 主成分包括SL、SW、ST、SY、Pe,第2 主成分包括HL、SL/SW。表明第1主成分主要描述種子大小,第2 主成分主要描述種子形狀。 楊生華等[38]和侯維海等[36]分別對蠶豆(Vicia faba)和青稞種子表型性狀進行的主成分分析都表明第1 主成分涵蓋了籽粒大小信息,第2 主成分涵蓋了籽粒形狀信息。在特定環境中物種種子大小對其幼苗的建植和存活具有顯著影響。因此,植物種子性狀遺傳變異的信息可為新品種選育提供重要的數據,尤其是對種子產量的提高以及幼苗建植和存活情況的改良具有十分重要的意義[39]。在本研究中,胡枝子屬各種質間和種內種子性狀均具有顯著的(P<0. 05)基因型變異,這些性狀可用于胡枝子屬植物分類學研究。
4 結論
種子豐富的表型性狀信息對于優良品種的選育及保存具有重要意義,且種子的大小對植物幼苗的建植和存活具有顯著影響。本研究結果表明,胡枝子種質及種間均存在顯著的基因型變異,表明胡枝子各種質種子遺傳多樣性豐富。性狀SY、ST 和Pe 遺傳穩定,可作為后期品種選育的參考指標。這不僅為胡枝子種質資源的收集、保存提供基礎數據,還有利于胡枝子改良育種工作的開展。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
