土壤水分和植株密度互作對垂穗披堿草地下營養繁殖及生物量的影響
2021-11-22 12:29:08王玲玲庫努都孜阿依吐魯洪孟廣飛郭正剛
草業學報 2021年11期
關鍵詞:植物
王玲玲,庫努都孜阿依·吐魯洪,孟廣飛,郭正剛
(蘭州大學草地農業生態系統國家重點實驗室,蘭州大學農業農村部草牧業創新重點實驗室,蘭州大學草地農業教育部工程研究中心,蘭州大學草地農業科技學院,甘肅 蘭州730020)
垂穗披堿草(Elymus nutans)是青藏高原地區天然草地的主要優勢禾本科植物[1],根系發達,生長期長[2],地下繁殖能力和可塑性較強,不僅是高寒地區天然草地的優良牧草組分[3],而且是青藏高原地區退化草地修復和栽培草地建植的主要草種[4]。大面積的垂穗披堿草栽培草地不僅為青藏高原地區畜牧業生產提供物質資料[5],而且逐漸成為該地區草牧業經濟發展的基石[4]。然而栽培草地具有自然衰退的特征[6],主要是種群自我更新能力弱所致[1]。雖然垂穗披堿草能夠實現種子繁殖[4],但青藏高原地區海拔高、溫度低、生長季短,在這種嚴酷的環境下,垂穗披堿草栽培草地更傾向于營養繁殖[7],實現其通過自我更新而維持持續生產的目標[8]。植物地下營養繁殖是植物營養繁殖的核心,被視為栽培草地種群自我更新和維持持續生產的驅動力[9]。植物地下營養繁殖主要通過克隆繁殖和芽庫繁殖適應嚴酷環境而維持其自我更新[10?11],其中克隆繁殖能力經常用克隆小株數和傳播距離測度[12],而芽庫繁殖能力則用地下芽數量或密度測度[13]。因此,研究垂穗披堿草克隆繁殖和芽庫繁殖,是青藏高原地區垂穗披堿草栽培草地維持穩產和持續利用的需求。
植物克隆繁殖和芽庫繁殖能力不僅受水分和光照等環境因子的影響[14],而且受植株密度的約束[15]。植株密度的變化能引起個體間的相互作用[16]。栽培草地密度往往具有自疏特性[17],出現個體變大而密度降低趨勢,表現出因植株密度降低而栽培草地呈現退化的現象,然而栽培草地又具有產量最后恒值法則[18],即植物個體密度變化引起的種群生物量波動,往往會通過種群的自我調節而逐漸彌補。土壤水分是植物生長的重要限制因子[19]。植株密度與土壤水分的關系既能互惠,又能拮抗。植株密度過大,會加快植物蒸騰作用;植株密度過小,地表蒸發過大,會影響土壤水分利用效率[20]。因此,只有植株密度和土壤水分耦合時,才能既維持植物穩產,又提高水分利用效率。雖然已有研究證實了土壤水分和植株密度對垂穗披堿草單株地上生物量有顯著的互作效應[18],但土壤水分和植株密度互作對垂穗披堿草地下營養繁殖特征是否產生影響,尚需要科學試驗提供證據。因此,本研究采用盆栽試驗,分析土壤水分和植株密度互作對垂穗披堿草克隆繁殖和芽庫繁殖能力以及生物量的影響,不僅為揭示土壤水分和植株密度互作影響垂穗披堿草單株地上生物量的機制提供了重要信息,而且為青藏高原地區天然草地合理管理和垂穗披堿草栽培草地生產力的持續維持提供了理論依據。
1 材料與方法
1. 1 試驗設計
采用盆栽試驗,設計為雙因素處理,兩個因素分別是土壤水分和植株密度。選用口徑長24 cm,寬10 cm,底徑長20 cm,寬10 cm,高12 cm 的聚乙烯塑料方形花盆,土壤水分設計為3 個水平,分別為土壤飽和含水量的30%,50% 和80%,分別用W1,W2,W3表示,植株密度設計為4,8,12 株·盆?13 個水平,分別用R1,R2,R3表示,共計9 個處理,即W1R1,W1R2,W1R3,W2R1,W2R2,W2R3,W3R1,W3R2,W3R3,每個處理6 個重復,共計54 個花盆。
垂穗披堿草種子來自青海省畜牧科學院,試驗地點為蘭州大學榆中校區的智能溫室,白天室內溫度保持在25 ℃,夜間保持在15 ℃,光照強度>600 μmol·m?2·s?1。試驗采用的土壤pH 為7. 4,有機質含量為0. 66%,全氮含量為0. 11%,速效磷含量為57. 66 mg·kg?1,速效鉀含量為240 mg·kg?1[5]。試驗于2019 年5 月16 日開始,首先,向另外的4 個盆中裝入2. 16 kg 的土,測定土壤的飽和含水量;其次,向54 個盆中裝入2. 16 kg 的土壤,根據土壤飽和含水量的60% 向每個花盆緩慢澆水,使花盆內土壤水分相對均勻分布;第三,每個花盆完成澆水后,于翌日(2019 年5 月17 日)播種,每個花盆播種籽粒飽滿的種子20 粒,此后每隔2 d 通過稱重法監測土壤水分,讓土壤水分繼續維持于土壤飽和含水量的60%,同時觀測種子發芽情況;第四,根據垂穗披堿草種子發芽檢測標準,11 d 認為其完成發芽[21],即2019 年5 月29 日開始按照試驗設計調整每個花盆的土壤水分,讓其達到試驗設計的水平;第五,各個花盆土壤水分達到試驗設計水平后,即2019 年5 月30 日開始采用手工間苗法,將各個花盆的植株密度調整到試驗設計水平。此后至試驗結束,每隔2 d 繼續采用稱重法控制土壤水分到試驗設計水平。試驗期間,每隔7 d 隨機挪動花盆位置,讓每個花盆的植物生長環境相對均勻一致。
1. 2 指標測定
根據垂穗披堿草生長習性[22],2019 年10 月1 日取樣。首先,針對花盆內的每株植株,用手輕輕地撥開植株根莖處的表層土壤,讓該植株的根莖充分暴露出來,然后沿著每條根莖的方向,確定其是否有克隆小株,最后記錄每株植物的所有克隆小株數;其次,每株植物有多個克隆小株,采用直尺測定各個克隆小株之間的直線距離,將其中最大的距離,記錄為這個植株的傳播距離;第三,測定完克隆小株和傳播距離后,將花盆中每個植物的地上和地下部分整體取樣(每個植物包括其所有克隆小株),用清水輕輕沖洗植株地下部分附著的土壤,沖洗干凈后,采用電子顯微鏡,鑒定每株植物地下部分附著的地下芽,然后根據地下芽附著的位置,將地下芽分為分蘗芽和根莖芽,統計每株植物上的分蘗芽和根莖芽數量(實際鑒定地下芽時,因垂穗披堿草生長于溫室,尚未發現根莖芽);最后,用剪刀將每株植物的地上和地下部分分離,其中地下芽屬于地下部分,分別裝入信封袋中,做好標記,在65 ℃條件下烘干48 h 至恒重,待冷卻至室溫時再稱重。
1. 3 數據處理
首先,檢驗每個指標數據的正態性和同質性,若數據不服從正態分布,則采用log 進行數據轉化,然后分析。采用一般線性模型對每個指標做雙因素方差分析,其中將土壤水分和植株密度設置為固定因子,克隆小株數、傳播距離、地下芽數量和生物量設置為響應變量,當方差分析結果顯著時(P<0. 05),采用Duncan 法進行多重比較分析,以確定各處理間的差異;若土壤水分和植株密度交互作用顯著時,采用MATLAB 2016b 構建二元二次模型,確定交互作用顯著變量對土壤水分和植株密度的響應過程。最后,采用皮爾森相關性分析確定克隆小株數、傳播距離、地下芽數量和生物量之間的關系。用SPSS 22. 0 進行雙因素方差分析和皮爾森相關性分析。
2 結果與分析
2. 1 克隆小株數和傳播距離
植株密度、土壤水分以及二者互作對垂穗披堿草克隆小株數和傳播距離均有極顯著影響(P<0. 01)。隨土壤水分增加,克隆小株數呈先增加后減小的趨勢,傳播距離呈增加趨勢。隨植株密度增加,克隆小株數和傳播距離均呈減小趨勢(表1)。

表1 土壤水分和植株密度對垂穗披堿草克隆小株數和傳播距離的影響Table 1 Effects of soil moisture and plant density on ramets per plant and spreading length
克隆小株數F(x2)與土壤水分(x1)、植株密度(x2)的關系為F(x1,x2)=?5. 76+0. 5294x1?0. 06079x2?0. 004027x12?0. 0225x22?0. 01158x1x2,F 檢驗時,P=0. 0001,R2=0. 970,說明土壤水分(x1)、植株密度(x2)與克隆小株數具有顯著回歸關系。x1的一次項系數為正數,x2的一次項系數為負數,且土壤水分(x1)系數絕對值大于植株密度(x2)系數絕對值,表明在一定范圍內土壤水分增加克隆小株數的效應要大于植株密度減少克隆小株數的效應;x1、x2的二次項系數均為負數,表明該面為一個開口向下的拋物面(圖1A),說明垂穗披堿草克隆小株數在土壤水分和植株密度互作時理論上存在最佳組合,為W2R1。
傳播距離F(x1,x2)與土壤水分(x1)、植株密度(x2)的關系為F(x1,x2)=?0. 2551+0. 07884x1+0. 5946x2?0. 0001x12?0. 04375x22?0. 01091x1x2,F 檢驗時,P=0. 007,R2=0. 702,說明土壤水分、植株密度與傳播距離具有顯著回歸關系。x1、x2的一次項系數均為正數,且土壤水分(x1)系數絕對值小于植株密度(x2)系數絕對值,表明在一定范圍內土壤水分增加傳播距離的效應要小于植株密度增加傳播距離的效應。x1、x2的二次項系數均為負數,表明該面為一個開口向下的拋物面(圖1B),說明垂穗披堿草傳播距離在土壤水分和植株密度互作時理論上存在最佳組合,為W3R1。

圖1 土壤水分和植物密度互作下垂穗披堿草克隆小株數和傳播距離曲面圖Fig. 1 3D response surface of ramets per plant and spreading length of E. nutans between soil moisture and plant density
2. 2 地下芽數量
土壤水分、植株密度及其二者互作均顯著影響垂穗披堿草地下芽數量(P<0. 05)。隨土壤水分增加,地下芽數量呈增加趨勢,但隨植株密度增加,地下芽數量呈減小趨勢(表2)。

表2 土壤水分和植株密度互作對垂穗披堿草地下芽數量的影響Table 2 Effects of soil moisture and plant density on underground buds per plant
地下芽數量F(x1,x2)與土壤水分(x1)、植株密度(x2)的關系為F(x1,x2)=6. 747?0. 0403x1?1. 97x2+0. 00074x12+0. 2479x22?0. 0038x1x2,F 檢驗時,P=0. 028,R2=0. 573,說明土壤水分、植株密度與地下芽數量具有顯著回歸關系。x1、x2的一次項系數均為負值,且土壤水分(x1)系數絕對值小于植株密度(x2)系數絕對值,表明在一定范圍內土壤水分減少地下芽數量的效應要小于植株密度減少地下芽數量的效應,x1、x2的二次項系數均為正數,表明該面為一個開口向上的拋物面(圖2),說明垂穗披堿草地下芽數量在土壤水分和植株密度互作時存在最小值,為W2R3,且理論上的最佳組合為W3R1。
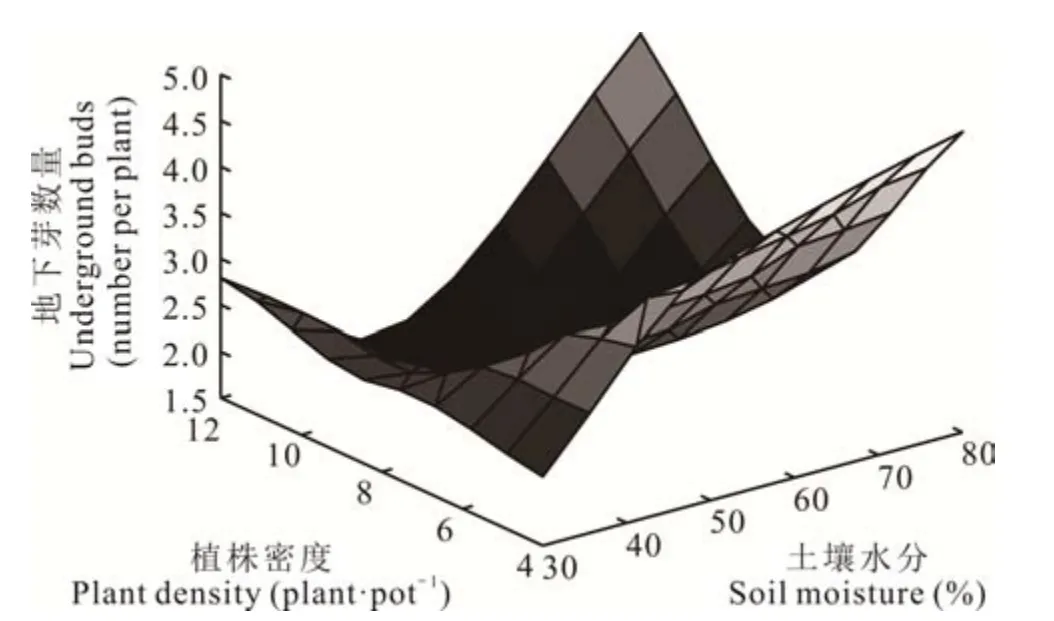
圖2 土壤水分和植株密度互作下垂穗披堿草地下芽數量曲面圖Fig. 2 3D response surface of underground buds per plant of E. nutans between soil moisture and plant density
2. 3 生物量
土壤水分、植株密度及二者互作對垂穗披堿草地上生物量、地下生物量及總生物量均有顯著影響(P<0. 05)。隨土壤水分增加,地上生物量、地下生物量及總生物量均呈增加趨勢;隨植株密度增加,地上生物量、地下生物量及總生物量均呈減小趨勢(表3)。
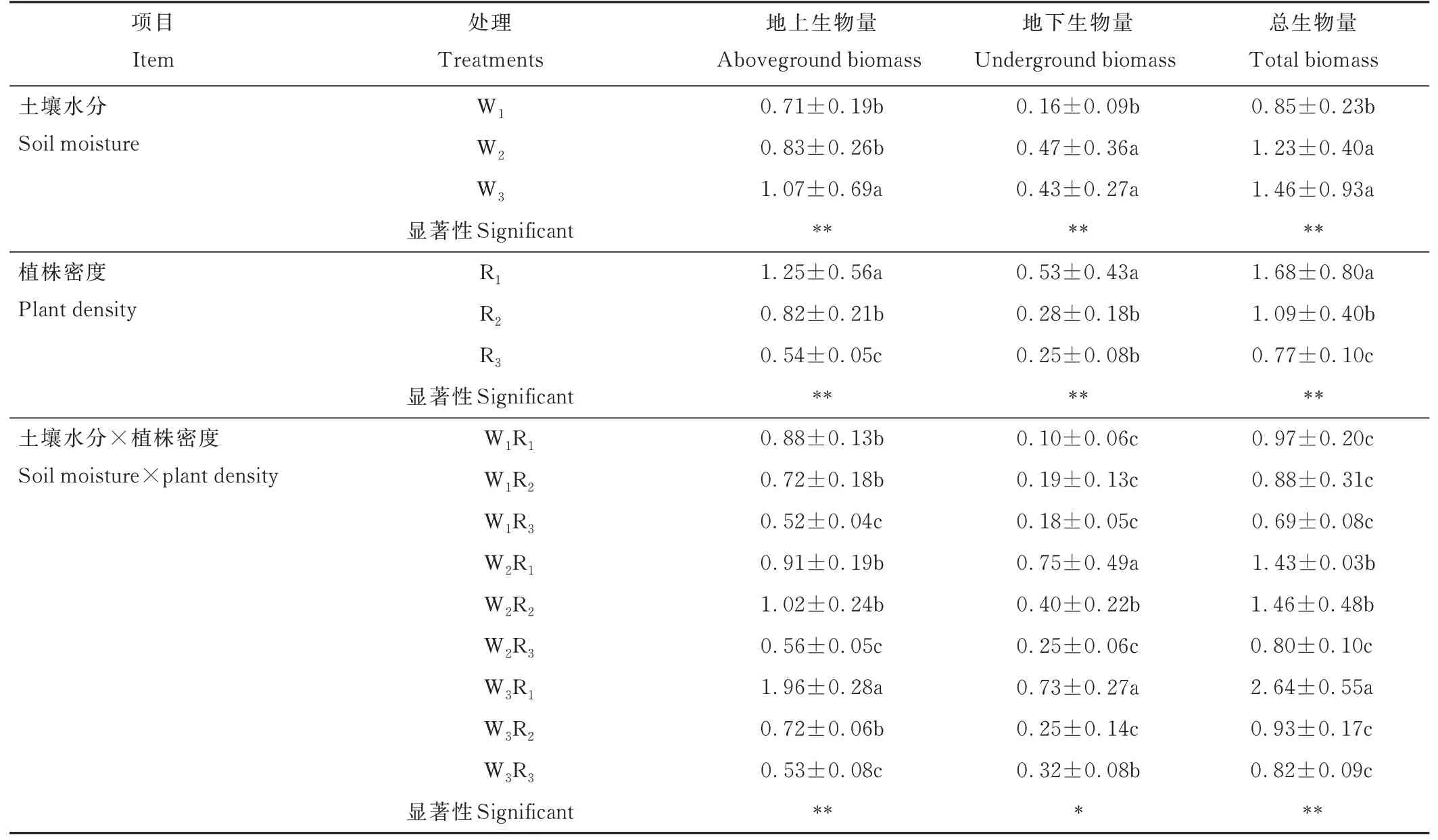
表3 土壤水分和植株密度互作對垂穗披堿草生物量的影響Table 3 Effects of soil moisture and plant density on biomass per plant of E. nutans(g·plant-1)
地上生物量F(x1,x2)與土壤水分(x1)、植株密度(x2)的關系為F(x1,x2)=0. 3316?0. 02579x1?0. 01215x2+0. 00004x12+0. 004583x22?0. 002819x1x2,F 檢驗時,P=0. 00001,R2=0. 912,說明土壤水分、植株密度與地上生物量具有顯著回歸關系。x1、x2的一次項系數均為負值,且土壤水分(x1)系數絕對值大于植株密度(x2)系數絕對值,表明在一定范圍內,土壤水分減產地上生物量的效應要大于植株密度減產的效應,x1、x2的二次項系數均為正數,表明該面為一個開口向上的拋物面(圖3),說明垂穗披堿草地上生物量在土壤水分和植株密度互作時存在最小值,為W1R3,理論上最佳組合為W3R1。
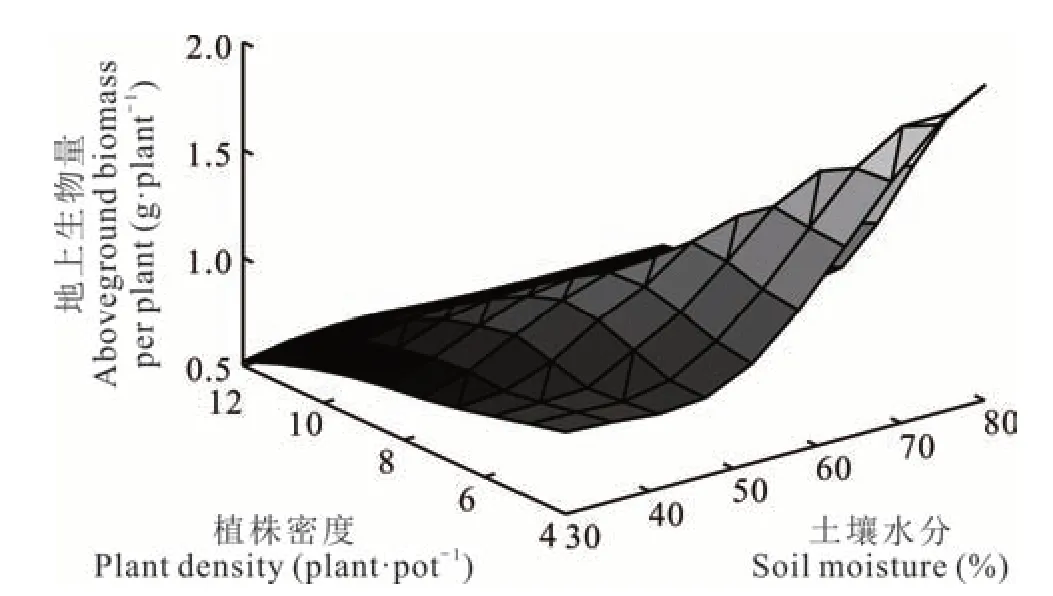
圖3 土壤水分和植株密度互作下垂穗披堿草地上生物量曲面圖Fig. 3 3D response surface of aboveground biomass of E. nutans between soil moisture and plant density
地下生物量F(x1,x2)與土壤水分(x1)、植株密度(x2)的關系為F(x1,x2)=?0. 6377+0. 05087x1?0. 08432x2?0. 00033x12+0. 0068x22?0. 0011x1x2,F 檢驗時,P=0. 043,R2=0. 585,說明土壤水分、植株密度與地下生物量具有顯著回歸關系。x1的一次項系數為正值,x2的一次項系數為負值,且土壤水分(x1)系數絕對值小于植株密度(x2)系數絕對值,表明在一定范圍內,土壤水分增產地下生物量的效應要小于植株密度減產的效應,x1的二次項系數為負數,x2的二次項系數為正數,表明垂穗披堿草地下生物量受土壤水分和植株密度互作影響時較為復雜(圖4)。互作時存在最小值,為W1R1,理論上的最佳組合為W2R1。
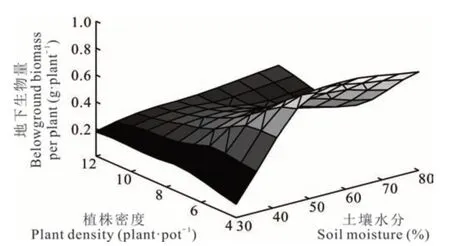
圖4 土壤水分和植株密度互作下垂穗披堿草地下生物量曲面圖Fig. 4 3D response surface of belowground biomass of E. nutans between soil moisture and plant density
總生物量F(x1,x2)與土壤水分(x1)、植株密度(x2)的關系為F(x1,x2)=?0. 39+0. 06889x1?0. 03875x2?0. 00022x12+0. 008437x22?0. 003938x1x2,F 檢驗時,P=0. 00001,R2=0. 861,說明土壤水分、植株密度與總生物量具有顯著回歸關系。x1的一次項系數為正值,x2的一次項系數為負值,且土壤水分(x1)系數絕對值大于植株密度(x2)系數絕對值,表明在一定范圍內,土壤含水量增產總生物量的效應要大于植株密度減產的效應,x1的二次項系數為負值,x2的二次項系數為正數,說明垂穗披堿草總生物量受土壤水分和植株密度互作影響時較為復雜(圖5)。互作時存在最小值,為W1R3,理論上的最佳組合為W3R1。

圖5 土壤水分和植株密度互作下垂穗披堿草總生物量曲面圖Fig. 5 3D response surface of total biomass of E. nutans between soil moisture and plant density
2. 4 克隆繁殖和芽庫繁殖與生物量間的關系
皮爾森相關分析結果表明,地上生物量、地下生物量與地下芽數量、克隆小株數和傳播距離均呈顯著正相關(表4),說明地下營養繁殖能力與垂穗披堿草栽培草地生產力具有密切關系,一定程度上說明垂穗披堿草地下營養繁殖能力提高,不僅有助于增加垂穗披堿草的產量,還有助于增加垂穗披堿草的生產力。

表4 各因素的相關系數Table 4 The correlation coefficient of parameters
3 討論
垂穗披堿草作為優良禾本科牧草[1],其種群自我更新能力事關高寒地區栽培草地的持續生產[12]。本研究采用盆栽試驗從克隆繁殖和芽庫繁殖兩個方面,研究了不同土壤水分和植株密度互作下垂穗披堿草的克隆小株數,傳播距離,地下芽數量,生物量的變化特征,以及克隆繁殖和芽庫繁殖與生物量之間的關系,解析垂穗披堿草栽培草地適應青藏高原地區水分環境變化和種植密度變化的可能途徑,為垂穗披堿草栽培草地穩定持續生產提供理論依據。研究結果表明,垂穗披堿草克隆小株數隨土壤水分增加呈先增加后減小的趨勢,這與苔草屬植物[23]和禾本科植物玉米(Zea mays)[24]的克隆小株數響應土壤水分的過程趨同,說明土壤水分較低或過高,均不利于植物克隆小株數的形成,只有土壤水分適宜時才能促進植物形成更多克隆小株數,從而通過增加垂穗披堿草克隆繁殖能力而很好地適應環境。垂穗披堿草傳播距離隨土壤水分增加而增加,這不同于美麗箬竹(Indocalamus decorus)克隆分株與土壤水分的關系[25],但類似于淫羊藿(Epimedium brevicornum)傳播距離與土壤水分的關系[14],這主要與植物自身的生物學習性有關。若植物屬于濕生植物,其傳播距離受土壤水分的影響較小,但植物屬于中生或旱生植物時,其傳播距離受土壤水分影響較大,以幫助植物在更大空間內獲取更多資源[23,26?27]。研究結果表明,垂穗披堿草地下芽數量隨土壤水分增加而增加,這與Aarssen[28]提到的地下芽數量或密度響應土壤水分的變化趨勢一致,主要是土壤水分能夠調控地下芽的萌發。植物克隆小株數和地下芽是一個互作系統[29],當土壤水分適宜時,地下芽則會萌發為克隆小株,此時克隆小株數增加,而地下芽數量或密度維持在較低水平,而當土壤水分過大時,此時地下芽萌發為克隆小株的速度受抑,導致克隆小株數數量降低,而地下芽數量或密度維持在較高水平,因此垂穗披堿草克隆小株和地下芽數量對土壤含水量的響應本質是植物自我調節的機制之一。垂穗披堿草地上和地下生物量隨土壤水分增加呈增加趨勢,這與藜屬植物在智利地區響應水分條件的結果趨同[30],當土壤水分能夠滿足植物生長需要時,則會增加植物地上和地下生物量[31],然而本研究亦發現垂穗披堿草地上和地下生物量增加時所需的土壤水分存在一定的差異,植物地上生物量增加所需的土壤水分要大于植物地下生物量增加所需的土壤水分標準,主要是植物根系首先獲取土壤水分,滿足自身生長需求,然后才通過自我調節機制向地上部分輸送水分。當土壤水分條件滿足植物生長需要的時候,垂穗披堿草通過增加克隆小株數而增加地上生物量。水分能改變禾本科植物根莖形態特征,土壤水分增加會增加垂穗披堿草根莖體積和數量[5],從而增加垂穗披堿草地下生物量,而地下生物量的增加,會促進垂穗披堿草更好地利用土壤養分資源和水分資源[25],反哺地上生物量[32]。地上地下生物量隨土壤水分增加,促進了垂穗披堿草總生物量的增加。
密度制約是自然植物種群普遍存在的特征,通過改變種群內每個植株可獲得資源總量,即植物種群內部個體特征在很大程度上依賴于密度變化狀況。本研究發現,垂穗披堿草克隆小株數隨植株密度增加呈減小趨勢,這與Makihiko 等[33]采用方格模型(lattice model)模擬的結果一致。植株密度增加,每株植物的克隆小株數之間的競爭亦會增加,這會反過來抑制克隆小株數的增加[34]。因此植物密度和克隆小株數的變化過程實質是一個權衡效應過程,主要目的是維持栽培草地的產量相對穩定。垂穗披堿草傳播距離隨植株密度增加而減小,這與Wong 等[35]采用隨機個體模型(stochastic individual-based model)模擬的結果一致,植株密度較高時,資源匱乏,較短的傳播距離有助于抵抗這種情況下的外界干擾。垂穗披堿草地下芽數量隨植株密度增加呈減小趨勢,地下芽是地上部分生長的補償[36],密度較低時植株會產生更多克隆小株,并且地下芽數量或密度維持在一個較高水平;而密度較高時,資源相對受限,植物增加克隆小株及地下芽數量或密度的能力受抑[37],從而導致地上生物量、地下生物量、總生物量均呈減小趨勢,這亦與老芒麥(Elymus sibiricus)生物量對密度的響應結果一致[38]。密度較低的生境內,植物個體能夠充分利用環境資源,個體之間的競爭較弱或不存在;但當種群密度較大時,個體間維持正常生長,必然會形成資源競爭格局[16],且隨植株密度增加,個體間在競爭光照和養分資源方面的程度變強,光合作用減弱,從而地上生物量降低[39],至密度較高時,就會引起植物個體積累碳水化合物的能力下降,個體生長受阻[16],地下生物量亦減少。個體繁殖能力是栽培草地生產力的基礎,然而個體繁殖能力往往與植株密度有關,植物密度維持相對穩定時,則意味著栽培草地能夠持續生產[40]。個體繁殖能力因植物密度自疏而發生改變,這種改變若處于一定的密度閾值之內,則會維持栽培草地的持續生產,然而當密度過小時,則因栽培草地不能充分利用自然資源[16?17],而導致垂穗披堿草總生物量減小。
垂穗披堿草克隆小株數、傳播距離、地下芽數量與其地上和地下生物量呈顯著正相關關系,說明土壤水分和植株密度對垂穗披堿草生物量的增加具有積極效應,但同步存在明顯的互作效應,并不是土壤水分越大,植株密度越大,其克隆小株數、傳播距離、地下芽數量、地上生物量、地下生物量以及總生物量越大,而是具有最佳的理論組合,本研究設置的9 個處理中,克隆小株數、傳播距離、地下芽數量、地上生物量、地下生物量、總生物量對應的最佳組合分別為W2R1、W3R1、W3R1、W3R1、W2R1、W3R1。因此,垂穗披堿草栽培草地的管理,并不是種植密度越大越好,亦不是土壤水分含量越高越好,而是需要根據垂穗披堿草栽培草地的管理目標,采用合理的種植密度,維持合理的土壤水分水平,這為青藏高原地區垂穗披堿草栽培草地的穩定持續生產提供了理論基礎。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
