江蘇地區扁豆主要表型性狀的多樣性評價
2021-11-18 15:50:03蘇彩霞孟珊欒春榮顏偉狄佳春朱銀張旭
江蘇農業科學 2021年19期
蘇彩霞 孟珊 欒春榮 顏偉 狄佳春 朱銀 張旭

摘要:利用表型特征,對收集的101份扁豆資源進行多樣性評價。結果表明,花序長短、莢厚、單株莢數等與產量密切相關;播種至始花天數與單株莢數、單株產量的關系較為密切;播種至盛花天數、播種至采收的天數與單株莢數、單株產量的關系則不顯著;而花序長短則影響著莢長、莢寬、單莢質量和產量;聚類分析表明,品種間存在顯著的遺傳差異,在歐氏距離為0.74閾值時,可將101份材料分為5類,形態與地理來源無嚴格的一致性關系。
關鍵詞:扁豆;江蘇省;表型特征;種質資源;多樣性評價;遺傳基礎
中圖分類號:S643.502 文獻標志碼: A文章編號:1002-1302(2021)19-0024-05
扁豆[Lablab-purpureus (Linn.) Sweet]是豆科、扁豆屬的一個栽培種,是多年生或一年生纏繞藤本植物,主要食用嫩莢或成熟豆粒。在我國,扁豆既可當糧食又可以當蔬菜用,在全國的種植面積約21.6萬hm2,單季平均產鮮豆莢約22 500 kg/hm2。近年來,隨著市場的發展以及人們對扁豆營養價值的進一步認識,扁豆的產業化進程正在逐步加速,作為其支柱的品種創新和品質創新工作也被提上了議事日程,而種質資源的收集和鑒定是進行該工作的前提和基礎。因此,筆者所在課題組近年來對江蘇地區的扁豆種質資源進行了廣泛的收集,為了更有效地對所收集的材料進行鑒定和利用,必須對其進行評價。
雖然用于研究植物種質變異和遺傳關系有種子蛋白、同工酶等許多標記[1],但種質評價和分類的第1步仍然是形態特征的鑒定[2],即形態學標記是植物種間或種內分類的重要依據之一[3]。據報道,形態學標記的多樣性分析已用在芭蕉、草莓、甜瓜、甘蔗、菊苣等作物上[4-9]。但在扁豆作物上,對江蘇地區扁豆多樣性評價方面的研究卻少見報道。
因此,本研究于2019年決定對收集自江蘇地區的101份材料(含從上海地區引進的材料)進行植物形態學特征觀測和鑒定,通過多元統計分析數量性狀和質量性狀,并以此為基礎,進行相關性和聚類分析,以找出不同類型材料差異的主要性狀,旨在為鑒別扁豆特異種質,擴大我國扁豆基因資源,以及扁豆新種質的創制等方面提供依據。
1材料與方法
1.1試驗區概況
試驗地設在江蘇省泰興市農業科學研究所內,地理坐標為31°58′12″~32°23′05″N,119°54′05″~120°21′56″E,季風顯著,四季分明,氣候溫和,雨量充沛,日照充足。年平均氣溫15.0 ℃,年平均降水量1 039.7 mm,年平均日照時數1 984.5 h。年平均日照百分率45%。
1.2試驗材料
供試扁豆種質主要來源于江蘇地區第三次農作物種質資源普查材料(含從上海地區引進的5份材料),材料編號、名稱及來源見表1,總計101份。
1.3試驗內容與方法
試驗采用一條龍區組設計,每個小區面積為 30 m2,行距60 cm,穴距1 m,每穴2~3粒種子,最后留苗1株。四周設保護行。測定表型性狀時,每個指標重復10次。具體測量數量指標有12個(播種至始花天數、播種至盛花天數、播種至采收天數、花序長、莢長、莢寬、莢厚、單莢粒數、單莢質量、單株莢數、 單株產量、 百粒質量),質量性狀指標有19個(莖色、葉色、葉脈色、葉片大小、翼瓣色、花色、莢形、縫線色、嫩莢色、莢壁纖維、品質、熟性、抗病性、抗蟲性、抗逆性、干籽粒種皮色、是否帶花紋、粒形、光澤度)。熟性、抗病性、抗蟲性、抗逆性在生長后期觀察記載,干籽粒的種皮色、是否帶花紋、粒形、光澤度、百粒質量,在種子收獲曬干后測定。其他性狀基本在花莢期測定。
1.4統計分析
利用Excel軟件對數量性狀進行變異系數分析。利用NTSYSpc軟件對性狀進行材料間歐氏距離分析,質量性狀指標首先在Standardization模塊中標準化為(0,1)矩陣,再在SAHN模塊中采用可變類平均法進行聚類分析(un-weighted pair-group method with arithmetic mean algorithm,UPGMA)。用SPSS 17.0軟件對數量性狀進行方差分析、相關分析,確定反映扁豆差異的主要形態學指標。
2結果與分析
2.1扁豆形態變異分析
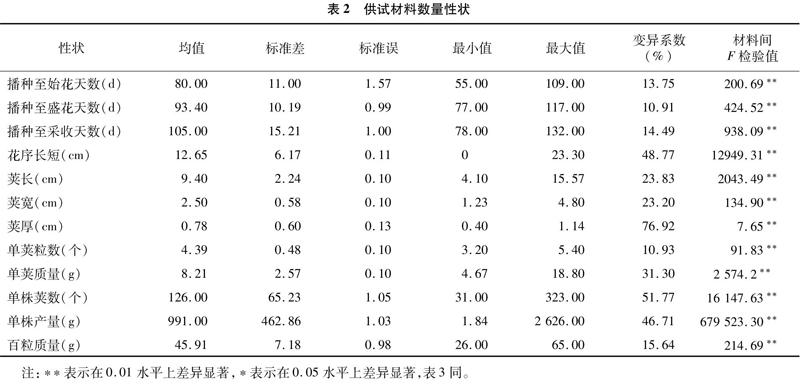
對101份扁豆材料間主要數量性狀變異情況進行分析(表2)。結果顯示,12個數量性狀差異均達到極顯著水平,通過材料間F檢驗值比較,單株產量的F檢驗值最大(679 523.30),莢厚最小(765)。變異系數結果顯示,莢厚的變異系數最大,達到7692%,說明莢厚潛力很大,其次為單株莢數(5177%),播種至盛花天數最少,僅10.91%,是比較穩定的植物學性狀。從整體變異程度來看,播種至始花天數、播種至盛花天數、播種至采收天數等與熟性相關的性狀變異程度要小于花序長短、莢厚、單株莢數等與產量密切相關性狀的變異程度,表明扁豆不同性狀間存在較大的遺傳差異,品種的熟性主要由遺傳因素決定,與產量性狀相關的花序長短、莢厚、單株莢數等表型特征更容易受外界環境的影響。均值、標準差及變異系數等具體參數見表2。
2.2扁豆植物學形態指標之間的相關分析
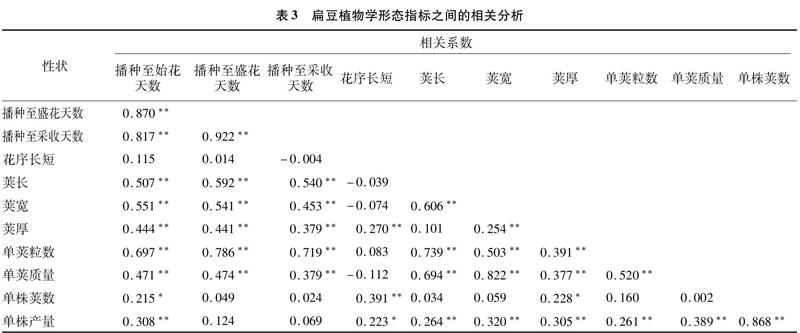
將扁豆不同材料各植物學形態數量指標進行相關分析(表3),結果表明,播種至始花天數與各性狀均呈正相關,除與花序長短相關性不顯著外,與其他數量性狀的相關性均達到顯著或極顯著水平,說明開花期越早,與產量相關的莢長、莢寬、單株產量、單莢質量等也呈正增長;播種至盛花天數則與播種至采收天數、莢長、莢寬、莢厚、單莢粒數、單莢質量呈極顯著正相關,與花序長短、單株產量、 單株莢數正相關性不顯著;播種至采收天數與花序長短呈負相關,但相關性不顯著,與其他性狀均呈正相關,且除單株產量、單株莢數外,正相關性均達到極顯著水平;花序長短與莢長、莢寬、單莢質量呈負相關,與其他性狀呈正相關,說明隨著花序的增長,扁豆莢長、莢寬、單莢質量會減少,其原因可能與養分的供應、分配相關,如果分配給花器官的養分多了,則供應給嫩莢的養分必然有所減少;另外,莢長與莢厚、單株莢數呈不顯著正相關,與莢寬、單莢粒數、單莢質量、單株產量呈極顯著正相關;除單株莢數外,莢寬與其他性狀均呈極顯著正相關;除莢長外,莢厚與其他性狀均呈顯著或極顯著正相關;其他幾個性狀彼此間也呈正相關(表3)。總之,播種至始花天數與單株莢數、單株產量的關系較為密切;播種至盛花天數、播種至采收天數與單株莢數、單株產量的關系則不顯著;而花序長短則影響著莢長、莢寬、單莢質量和產量,說明如何選擇一個適宜的始花期和適宜花序長度的品種,使產量、效益達到最大化,是進行新品種選育時值得關注的一個問題。
2.3101份扁豆種質聚類分析
為了研究扁豆種內親緣關系,采用12個主要數量性狀形態指標以及遺傳上較為穩定、不易受環境影響的質量指標計算材料間的歐氏距離,利用可變類平均法進行聚類分析(圖1)。總體看,品種間存在顯著的遺傳差異。在歐氏距離為0.74閾值時,101份材料分為5組,前3組材料占總材料的9802%,基本是按照莢形分為3個類型, 即:(1)青
皮、白皮嫩莢、直刀皮條白花型;(2)紫花、紫紅嫩莢、鐮刀型;(3)青白紅邊嫩莢、紫花型。第1組材料37份(材料編號主要為1、2、5、8、12、18、19、20等),約占總材料的36.63%,這部分材料大多為白皮扁豆,皮薄,籽粒大,糯性強,但纖維稍多,易老化,口感稍差;第2組材料62份(材料編號主要為3、21、53、24、75、71等),約占總材料的61.38%,這部分材料大多花青素含量高,肉質厚,籽粒大,纖維含量少,口感優良;第3組材料4份(材料編號為15、89、90、91),這部分材料早期為青白紅邊,生長中期見光部分逐步由沙紅色到紅色,老熟時紅色又逐步消失,可能是第1組和第2組的中間過渡型;第4組是87號,名為句容紅邊扁豆,花色紫,無花序或超短花序,鮮豆莢長鐮刀形,紅色縫線,青白帶沙紅色嫩莢。但嫩莢比一般扁豆更長、更寬;第5組是40號啟東刀扁豆,花色白,中長花序,鮮豆莢蔥管形、青綠色縫線,青白色嫩莢,在扁豆的莢型中也是獨樹一幟。從圖1中還可以看出,上海市的5個品種(編號為92、93、94、95、96)莢形雖均為豬耳朵,嫩莢較厚,但均在第2組,可見形態與地理來源無嚴格的一致性關系。
3討論與結論
種群內遺傳變異的大小及種群結構決定了一個物種的進化潛力和抵御不良環境的能力[10]。表型性狀是種質遺傳多樣性分析的最常用方式[11]。本研究中,扁豆不同材料間存在顯著的遺傳變異,各性狀間均有一定程度的相關性,且大部分達到極顯著相關性。其中,與熟性相關的性狀變異程度總體小于與產量相關的性狀變異程度,說明產量性狀的遺傳潛力巨大,可以通過栽培措施來影響產量相關部分性狀,尤其是莢厚、單株莢數等,從而提高產量,增加效益。姜永平等曾指出,在評價品種間遺傳差異時,不能僅以地理來源為依據[12],對扁豆主要農藝性狀變異系數的分析表明,莢厚、單株產量、單株莢數等性狀的變異系數較大,超過了30%,而播種至始花天數、播種至盛花天數、播種至采收天數性狀的變異系數較小,說明扁豆種質資源中,莢厚、花序長短、單株產量的變異較為豐富,雖然熟期有些單一,但可引進不同成熟期的種質資源以豐富變異類型,從而選育不同生育期的品種。
表3中,播種至始花天數與單株莢數、單株產量有顯著或極顯著的相關性;播種至盛花天數、播種至采收天數與單株莢數、單株產量的關系則不顯著;說明播種至始花天數的重要性。生產上,我們常用播種至采收天數確定品種的熟性,本研究認為,應以播種至始花天數作為其主要的依據更為恰當。在進行扁豆新品種選育時,我們本能地以長花序品種作為優良親本的首選,本研究表明,要結合莢厚、單株莢數綜合考慮,使產量效益達到最大值時的花序長度才是最合適的。
在馴化物種演化過程中,地理因素發揮了重要的作用[13],用形態性狀進行聚類分析,能粗略地反映出材料之間的親緣關系。本研究所用101份材料,幾乎涵蓋了目前長三角地區絕大部分扁豆品種,通過聚類分析圖1,可將其分為5類,第1類、第2類數量較多,87號句容紅邊扁豆、40號啟東刀扁豆則各自成為一類,說明他們與其他品種性狀間存在顯著差異;上海市的5個品種(編號為92、93、94、95、96)均在第2類,表明本研究中收集的扁豆資源不存在地域特異差異,各資源分類并不與地域相關。這與國內外同行研究的結果[14-16]相同。
由于目前扁豆尚未進行全基因組測序,可用的標記太少,本試驗只進行了表型分析,至于通過分子標記來檢測和分析遺傳多樣性結果如何,有待進一步考證。
參考文獻:
[1]何慶元,王吳斌,楊紅燕,等. 利用SCoT標記分析不同秋眠型苜蓿的遺傳多樣性[J]. 草業學報,2012,21(2):133-140.
[2]Smith J S C,Smith O S.The description and assessment of distances between inbred lines of maize:the utility of morphological,biochemical and genetic descriptors and a scheme for the testing of distinctiveness between inbred lines[J]. Maydica,1989,34:151-161.
[3]周波,江海東,張秀新,等. 部分引進牡丹品種的形態多樣性[J]. 生物多樣性,2011,19(5):543-550.
[4]Ortiz R. Morphological variation in Musa germplasm[J]. Genetic Resources and Crop Evolution,1997,44(5):393-404.
[5]Swennen R,Vuylsteke D,Ortiz R.Phenotypic diversity and patterns of variation in west and central African Plantains(Musa spp.,AAB Group Musaceae)[J]. Economic Botany,1995,49:320-327.
[6]Harrison R E,Luby J J,Furnier G R,et al. Differences in the apportionment of molecular and morphological variation in North American strawberry and the consequences for genetic resource management[J]. Genetic Resources and Crop Evolution,2000,47(6):647-657.
[7]Muthy H N,Manohar S H.Estimation of phenotypic divergence in a collection of Cucumis melo,including shelf-life of fruit[J]. Scientia Horticulturae,2012,148:74-82.
[8]劉新龍,馬麗,蔡青,等. 云南甘蔗品種表型性狀的遺傳多樣性分析[J]. 植物遺傳資源學報,2010,11(6):703-708.
[9]梁小玉,張新全,白史且,等. 菊苣主要表型性狀的多元統計分析[J]. 草業學報,2013,22(6):257-267.
[10]Grant P R,Grant B R. Hybridization of bird species[J]. Science,1992,256(554):193-197.
[11]Fufa H,Baenziger P S,Beecher B S,et al. Comparison of phenotypic and molecular marker-based classifications of hard red winter wheat cultivars[J]. Euphytica,2005,145(1):133-146.
[12]姜永平,吳春芳,陳惠. 12個鮮食大豆數量性狀的主成分和遺傳距離分析[J]. 中國農學通報,2007,23(8):193-197.
[13]Jaradat A A,Shahid M. Patterns of phenotypic variation in a germplasm collection of Carthamus tinctorius L. from the Middle East[J]. Genetic Resources and Crop Evolution,2006,53(2):225-244.
[14]Maass B L,Jamnadass R H,Hanson J,et al. Determining sources of diversity in cultivated and wild lablab purpureus related to provenance of germplasm by using amplified fragment length polymorphism[J]. Genetic Resources and Crop Evolution,2005,52(6):683-695.
[15]Esther N K,Francis N W,Miriam G K.Molecular diversity of Kenyan lablab bean [Lablab purpureus (L.) Sweet] accessions using amplified fragment length polymorphism markers[J]. Amercian Journal of Plant Sciences,2012,3:313-321.
[16]姚陸銘,武天龍. 利用SSR標記及表型多樣性對扁豆遺傳多樣性的研究[J]. 上海農業學報,2016,32(5):1-7.
基金項目:江蘇省現代農業體系項目(編號:JATS[2021]274);2019年泰興市高層次人才培養工程培養對象科研資助項目(編號:泰人才辦[2019]14號);第三次全國農作物種質資源普查與收集行動(編號:111821301354052028)。
作者簡介:蘇彩霞(1974—),女,江蘇泰興人,碩士,高級農藝師,主要從事糧經作物新品種選育及配套栽培技術研究與示范。E-mail:2794785451@qq.com。
通信作者:欒春榮,碩士,農業推廣研究員,主要從事糧經作物新品種選育及配套栽培技術研究與示范工作。E-mail:lcrtx@163.com。
