云南牟定酸漿水中優勢產酸菌的分離 鑒定及生長特性
2021-11-05 10:44:36張宸瑞薛橋麗白彬陽胡永金
食品科學 2021年20期
關鍵詞:生長
張宸瑞,薛橋麗,白彬陽,胡永金,
(1.云南農業大學食品科學技術學院,云南 昆明 650201;2.云南農業大學學報編輯部,云南 昆明 650201)
腐乳作為我國特有的大豆發酵食品,其風味醇厚、濃香,口感滑潤、細膩,營養豐富[1]。云南牟定縣地理環境優越,氣候適宜,從明末清初時期生產腐乳,隨著生產工藝不斷精進,其口感、風味不斷豐富,如今牟定腐乳遠近聞名,成為云南腐乳的代表[2]。
牟定腐乳的獨到生產工藝為酸漿水點漿,酸漿水是豆腐壓制過程中滲出的黃漿水經乳酸菌、醋酸菌、酵母菌等微生物[3-4]發酵形成。自然環境中的微生物接觸黃漿水后,以黃漿水中的蛋白碎片、纖維碎片、多糖等大量營養物質作為營養源,通過酶解代謝產生大量醋酸、乳酸、草酸等幾十種有機酸,因此溶液整體酸價下降[5-6]成為豆腐點漿的酸漿水。酸漿水作為天然凝固劑不僅避免引入無機金屬離子,還賦予腐乳獨特風味,最大限度地開發黃漿水的應用效率,極大地減少黃漿水排放造成的環境污染[7-8]。但由于自然引入的微生物群落結構復雜,含有益菌的同時也摻雜有害微生物,導致產品存在安全隱患[9]。自然發酵的酸漿水受外界環境因素影響較大,難以保證每個批次的穩定品質,因此從自然發酵的酸漿水中分離優勢菌群,分析菌群的生長特性以篩選生長繁殖快、產酸能力強的菌株,采用分離篩選的純種菌發酵黃漿水即可解決上述問題。
目前,國內外對酸漿水中微生物的研究主要集中于篩選鑒定優勢乳酸菌、分析生長特性及測定代謝 產物[10-11],對酸漿水中的其他高產酸菌株及其特性研究鮮見報道。因此,本實驗分離、篩選、鑒定云南牟定2 家腐乳生產公司自然發酵酸漿水中的高產酸菌株,并通過生長特性研究明確不同高產酸菌株的生長條件及產酸性能,從中篩選生長迅速、產酸能力強的菌株,以期為綜合利用高產酸菌發酵生產提供具有潛在應用價值的菌株。
1 材料與方法
1.1 材料與試劑
酸漿水樣品 云南牟定兩家腐乳生產公司(A、B公司);改良MRS固體培養基、MRS液體培養基 上海撫生實業有限公司;增強革蘭氏染色液 北京雷根生物有限公司;新型生化鑒定管 廣東環凱微生物科技有限公司; 無水碳酸鈣、鹽酸、氯化鈉(均為分析純) 天津市天力化學試劑有限公司;納他霉素(食品級) 浙江新銀象生物工程有限公司。
1.2 儀器與設備
BCM-1000A超凈工作臺 蘇州安泰技術有限公司; Eppendorf移液槍 南京貝登醫療股份有限公司;9053A烘箱 上海實研電爐有限公司;HHPX-9272ME恒溫培養箱 上海博訊實業有限公司;Nikon E200-F顯微鏡 南京新飛達光學儀器公司;LX-B高壓蒸汽滅菌鍋 北京市永光明醫療儀器有限公司;TF-B60超低溫冰箱 上海賽默飛世爾科技公司;pH-10 pH計 廣州市銘睿電子科技有限公司;725N分光光度計 南京曉曉儀器設備有限公司。
1.3 方法
1.3.1 產酸菌的分離
采用平板涂布法,將稀釋后的酸漿水樣品涂布于添加質量分數0.1%納他霉素、質量分數2.0%碳酸鈣的MRS分離培養基上[12],37 ℃恒溫倒置培養24 h,挑取有溶鈣圈的單菌落純化3 代,轉接于斜面4 ℃保存。
1.3.2 高產酸菌的篩選
挑取保存菌株接種于MRS液體培養基中,37 ℃恒溫培養48 h,以總酸含量為指標,產酸量高于20 g/L的菌株為高產酸菌株。總酸測定參考王嘉琪等[13]的方法,以乳酸計,按下式計算待測液產酸量:
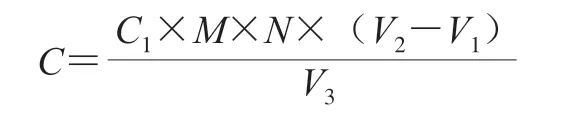
式中:C為待測液產酸量/(g/L);C1為標定的氫氧化鈉濃度/(mol/L);M為乳酸摩爾質量/(g/mol); N為稀釋倍數(10);V1為樣品待測液消耗氫氧化鈉溶液體積/mL;V2為空白待測液消耗氫氧化鈉溶液體積/mL;V3為樣品待測液體積/mL。
1.3.3 高產酸菌株的形態觀察和生理生化鑒定
用革蘭氏染色法對菌株進行形態學觀察[14]。將菌株制成菌懸液接入生理生化鑒定管中,37 ℃恒溫培養24 h后觀察記錄顏色變化,根據《常見細菌系統鑒定手冊》[15]進行判定。
1.3.4 菌株分子生物學鑒定
將菌株送至北京擎科生物科技有限公司,進行DNA提取、聚合酶鏈式反應(polymerase chain reaction,PCR)擴增及16S rDNA測序。
將獲得的測序結果提交NCBI的GenBank數據庫通過BLAST進行比對,確定種屬及親緣關系并進行ITS同源性分析,選取同源性高的菌株16S rDNA基因序列,采用MEGA 7.0構建發育樹。
1.3.5 自然發酵酸漿水及高產酸菌株的活化
將低溫保藏不同產地的的酸漿水樣品按體積分數3%接入MRS肉湯培養基中37 ℃恒溫發酵24 h,連續培養2 代,獲得活化酸漿水發酵液,將A公司活化酸漿水命名為Y,B公司活化酸漿水命名為X。用接種環挑取試管斜面保藏高產酸菌株分別接入MRS肉湯培養基中37 ℃恒溫發酵24 h,取3%發酵24 h的純菌種發酵液接入MRS肉湯培養基中,37 ℃恒溫發酵24 h,連續培養2 代以獲得活化高產酸菌發酵液。
1.3.6 生長曲線測定
取活化后的酸漿水發酵液、純菌種發酵液均按體積分數3%分別接入MRS液體培養基中,37 ℃恒溫發酵48 h,每隔4 h取發酵液,以滅菌后的MRS液體培養基為空白對照,采用分光光度計測定發酵液OD600nm值,重復測定3 次[16]。
1.3.7 產酸能力測定
取活化后的酸漿水發酵液、純菌種發酵液均按體積分數3%分別接入MRS液體培養基中,37 ℃恒溫發酵48 h,每隔4 h取發酵液作為待測液,測定總酸量及pH值,pH值用pH計測定,總酸測定同1.3.2節方法。
1.3.8 高產酸菌耐酸能力測定
取活化后的自然發酵酸漿水和純菌種發酵液,按3%接種量分別接入初始pH值為1.5、2.0、2.5、3.0、3.5、4.0、4.5、5.0、5.5、6.0的液體MRS培養基中,37 ℃恒溫培養24 h測定OD600nm值,重復測定3 次。
1.3.9 耐滲透壓能力測定
取活化后的酸漿水發酵液和純菌種發酵液,按體積分數3%分別接入NaCl質量濃度為0、2、4、6、8、10、12、14 g/100 mL的MRS肉湯培養基中,37 ℃恒溫培養24 h測定OD600nm值,重復測定3 次。
1.4 數據分析
采用SPSS 18.0進行數據分析、Origin 9.0進行繪圖,MEGA 7.0進行同源性分析及建立系統發育樹。
2 結果與分析
2.1 牟定腐乳酸漿水中產酸菌的篩選與鑒定
2.1.1 產酸菌的初篩
由于產酸菌代謝產生的乳酸與培養基環境中的不溶性碳酸鈣發生化學反應形成溶解度較大的乳酸[17],產生肉眼可見的透明溶鈣圈,因此選取產生溶鈣圈效果明顯的菌落于MRS固體培養基上反復劃線并分離、純化,獲得80 株產酸特征明顯的單菌落并進行4 ℃試管斜面保存。
2.1.2 高產酸菌的復篩
將80 株產酸菌進行產酸性能測定,分離自云南牟定A公司酸漿水中的10 株菌產酸量均高于20 g/L,分離自云南牟定B公司酸漿水中的8 株菌產酸量均高于20 g/L。
2.1.3 菌株的初步鑒定
對18 株高產酸菌進行形態學觀察,獲得10 株不同菌群特征、菌體特征的的革蘭氏陽性菌,對云南牟定A公司酸漿水中分離獲得的高產酸菌株重新編號為YQZ1~YQZ5,將活化酸漿水內自然菌群Y、YQZ1~YQZ5統稱為Y組,對云南牟定B公司酸漿水中分離獲得的高產酸菌株重新編號為QJZ1~QJZ5,將活化酸漿水內自然菌群X、XJZ1~XJZ5統稱為X組。
參閱《乳酸細菌分類鑒定及實驗方法》[18]、《伯杰細菌鑒定手冊》[19],根據生化鑒定結果比對《常見細菌系統鑒定手冊》[15]確定菌株YQZ1、YQZ3、XJZ2、XJZ3、XJZ5為乳桿菌屬(Lactobacillus),菌株YQZ4、XJZ1、XJZ4為腸球菌屬(Enterococcus),菌株YQZ2為乳球菌屬(Lactococcus),菌株YQZ5為微桿菌屬(Microbacterium)。
2.1.4 菌株分子生物學鑒定及發育樹建立
2.1.4.1 菌株基因序列測定
以10 株菌的DNA為模板,以27F、1492R為引物進行PCR擴增,產物進行1%尿蛋白普通瓊脂糖凝膠電泳,16S rDNA PCR電泳結果如圖1所示,10 菌株16S rDNA序列長度在1 500 bp有較清晰、明亮的特異性條帶,可進行基因序列測定[20]。
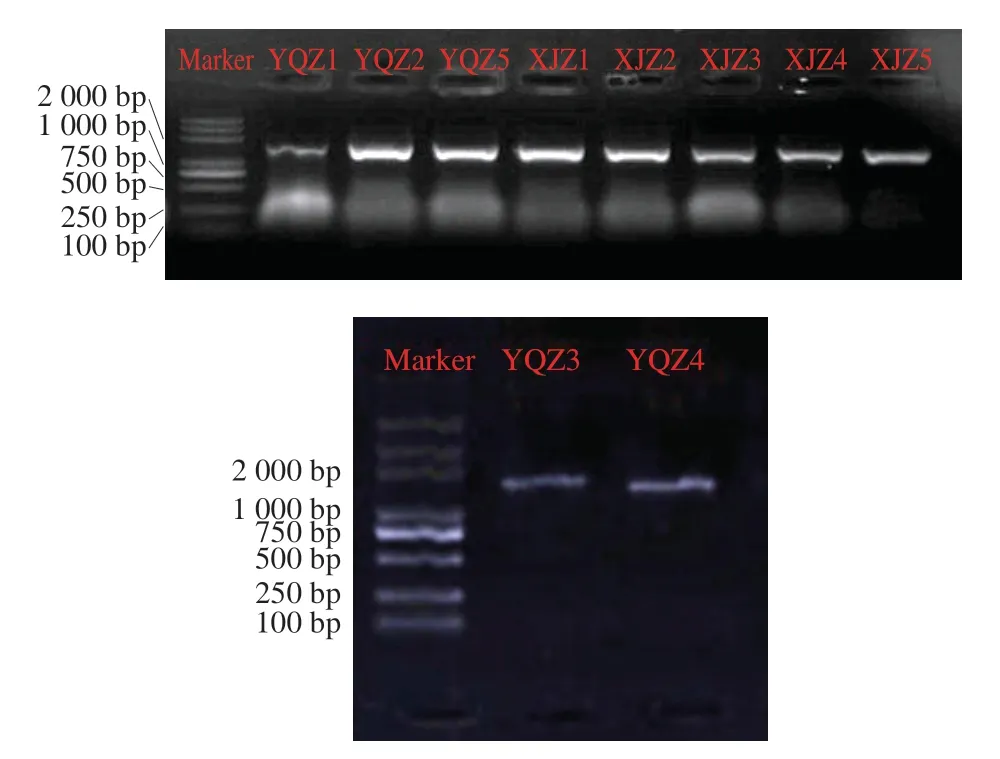
圖1 10 株菌株16S rDNA PCR擴增電泳圖Fig. 1 Electrophoresis of the PCR amplified products of 16S rDNA gene from 10 strains
2.1.4.2 菌株同源性分析及發育樹建立
將10 株產酸菌株測定結果錄入NCBI的BLAST程序上同源性分析比對,結果如表1所示。根據形態學分析、生理生化測定結果,結合同源性比對,確立菌株YQZ1~YQZ5分別為發酵乳桿菌(L. fermentum)、棉子糖乳球菌(L. raffinolactis)、植物乳桿菌 (L. plantarum)、鉛黃腸球菌(E. casseliflavus)、氧化微桿菌(M. oxydans),菌株XJZ1~XJZ5分別屎腸球菌(E. faecium)、L. plantarum、副干酪乳桿菌 (L. paracasei)、海氏腸球菌(E. hirae)、L. paracasei。10 株菌中乳酸菌有6 株、腸球菌有3 株、微桿菌有1 株,可確定乳酸菌為酸漿水中主要菌群。根據同源性比對結果,采用Clustalx裁剪基因序列,采用MAGE7.0對修剪后的基因序列建立發育樹,10 株菌系統進化樹如圖2、3所示。

表1 10 株菌株同源性分析Table 1 Homology analysis of 10 strains
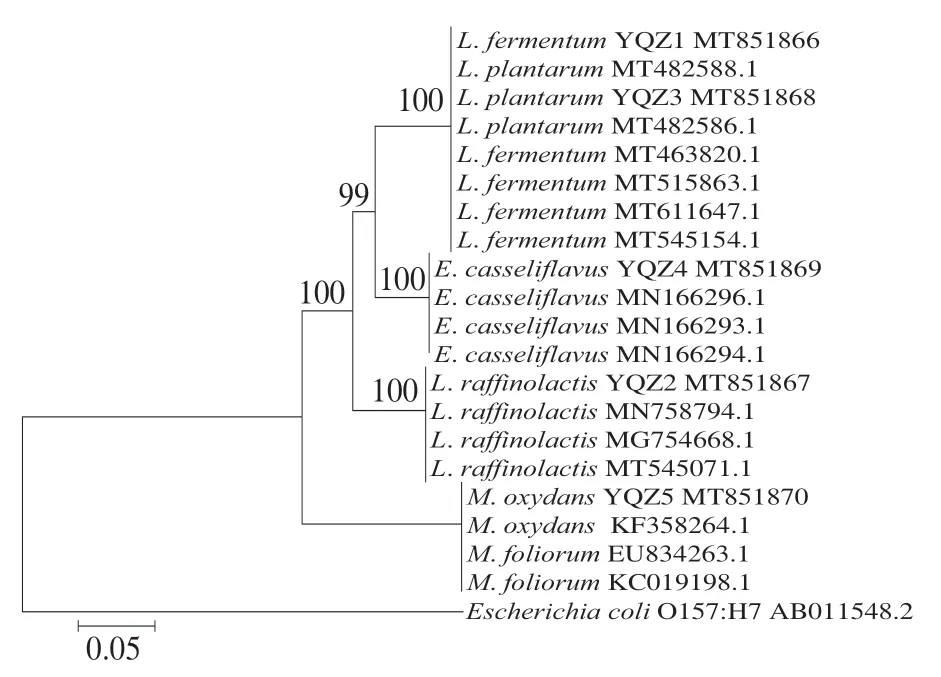
圖2 YQZ1~YQZ5菌株系統進化樹Fig. 2 Phylogenetic tree of strains YQZ1-YQZ5
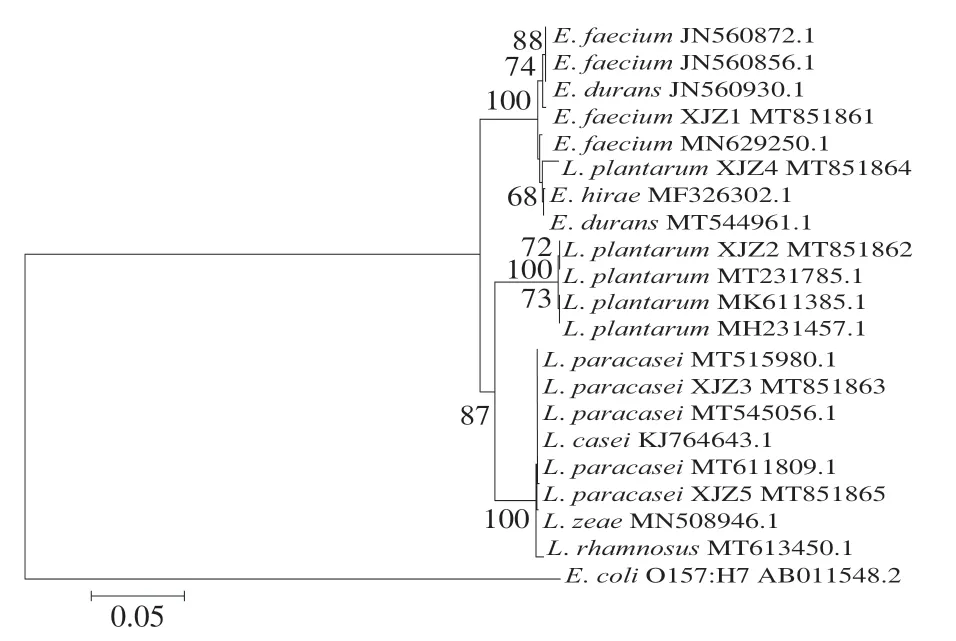
圖3 XJZ1~XJZ5菌株系統進化樹Fig. 3 Phylogenetic tree of strains XJZ1-XJZ5
2.2 產酸菌的生長特性分析
2.2.1 產酸菌生長曲線分析
如圖4所示,不同產地酸漿水中的自然菌群在不同生長周期內相較于大部分單菌株有一定生長優勢。自然菌群Y、X在短時間能迅速進入生長對數期,且對數生長期的生長速率明顯高于大多數菌株,說明自然菌群具有較強的適應能力和較高的繁殖速率。自然菌群Y、X在8 h后進入生長穩定期,且整個生長穩定期均保持較高菌體濃度(OD600nm),其濃度基本穩定在2.2,未出現較大幅度的波動,說明自然菌群具有較強的繁殖能力,且種間不具有多重競爭關系。Y組包含的自然菌群Y、菌株YQZ3在0~8 h呈對數生長,菌株YQZ1、YQZ4在0~16 h呈對數生長,菌株YQZ2在0~24 h呈對數生長,YQZ2較其他菌株的對數生長周期較長,且生長速率較緩。相同培養時間內,菌株YQZ1生長速率僅次于Y、YQZ3,但繁殖能力最強,48 h時菌體濃度為2.26,自然菌群Y次之,YQZ2菌體濃度最低。X組包含的菌株XJZ2、XJZ4、自然菌群X在0~8 h呈對數生長,菌株XJZ5在0~12 h呈對數生長,菌株XJZ1、XJZ3在0~16 h呈對數生長。相同培養時間內菌株XJZ2生長速率最快、活菌數目最高,48 h菌體濃度2.44,自然菌群X次之,XJZ4最低,因此菌株YQZ1、XJZ2均有非常理想的生長繁殖能力。

圖4 產酸菌株Y組(a)、X組(b)生長曲線圖Fig. 4 Growth curves of acid-producing strains from group Y (a) and group X (b)
2.2.2 菌株產酸性能分析
由圖5可知,菌群、菌株的產酸速率與生長趨勢呈正相關,即生長速度越快,產酸速率越快,尤其是處于生長對數期的菌株,能快速吸收培養基中的原營養物質和分解后的次級營養物質以代謝產酸,短時間內有機酸的累積量迅速增加。處于生長穩定期的菌株出生率和死亡率已逐漸達到平衡,營養物質的消耗率和代謝率基本穩定,該階段產酸速率較對數生長期的產酸率緩慢,但酸累積量穩步上升,于第48小時達到最高點。自然菌群Y、X在0~16 h代謝產酸速率高,多種微生物能夠在初始營養環境中最大限度地吸收各種營養滿足自身生長,不同菌株之間可能存在一定互生關系,更利于彼此生長代謝,進一步提高產酸速率。自然菌群Y、X的乳酸積累量分別在28~48、20~32 h出現下降,其原因可能為以有機酸為次生碳源的菌群在該階段生長活躍,有機酸分解代謝率高于產酸率,導致酸累積量下降,因此自然菌群中的乳酸累積并不穩定。從Y組可知,0~48 h內YQZ1的酸累積量最多,但在24~40 h產酸量大幅度下降后回升的原因可能為乳酸、乙酸、酒石酸代謝轉化為其他小分子產物,隨后又繼續累積,有機酸累積量最高,最終產酸量為43.61 g/L。菌株YQZ4產酸速率較低,且產酸量最少,最終產酸量為25.21 g/L。由X組可知,自然菌群X的產酸量在20~36 h先下降后回升,其他菌株菌基本保持較為穩定的遞增趨勢,XJZ2產酸速率最快,有機酸累積量最多,最終產酸量為50.91 g/L,菌株XJZ1的最終產酸量僅次于XJZ2,最終產酸量為49.93 g/L,菌株XJZ4產酸速率最慢,最終產酸量為34.68 g/L。從圖6可知,菌群的pH值在發酵初期隨著有機酸的快速累積,氫離子濃度的迅速升高而快速降低,隨之緩慢降低直至趨于平穩,Y組的自然菌Y、菌株YQZ1、YQZ2、YQZ3、YQZ4、YQZ5的最終pH值為3.98、3.75、3.84、4.01、3.98、4.02。X組的自然菌群X、菌株XJZ1、XJZ2、XJZ3、XJZ4、XJZ5的最終pH值為3.79、3.54、3.48、3.53、3.98、3.49。由圖5、6可知,隨YQZ1、XJZ2的產酸能力明顯高于其他菌株。
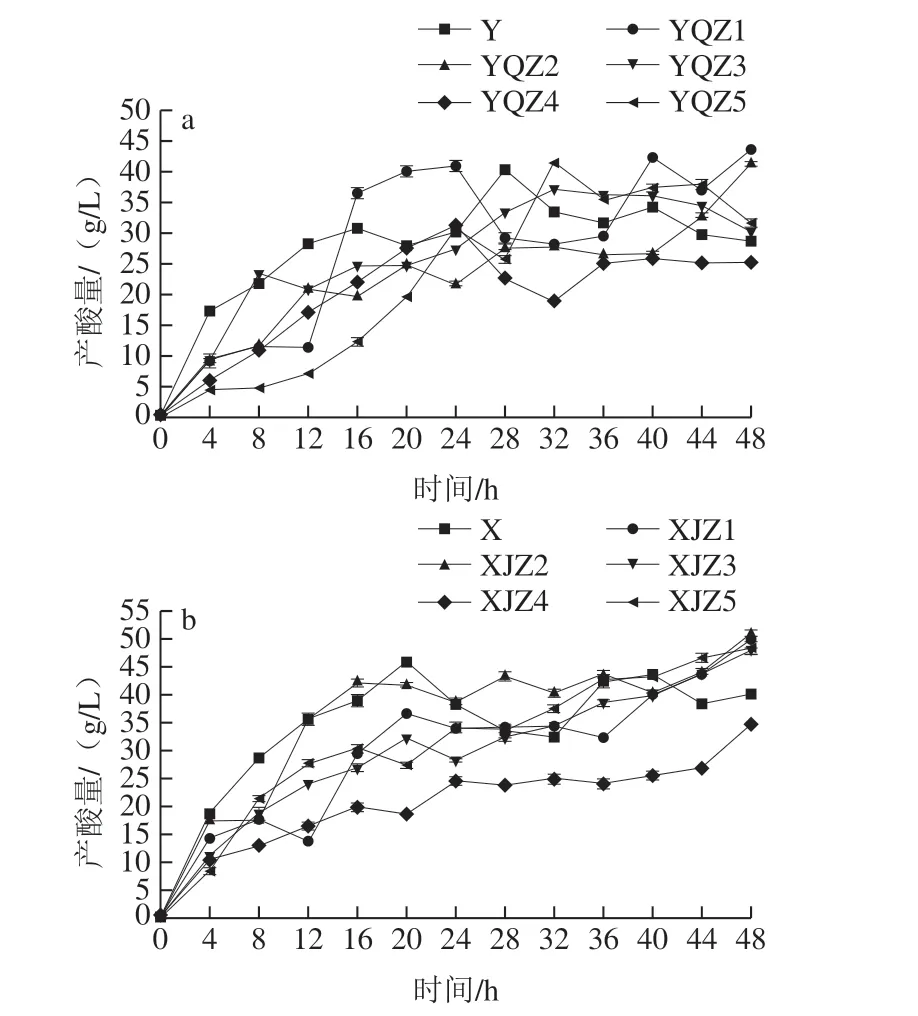
圖5 產酸菌株Y組(a)、X組(b)的產酸量Fig. 5 Acid-producing capacity of acid-producing strains from group Y (a) and group X (b)
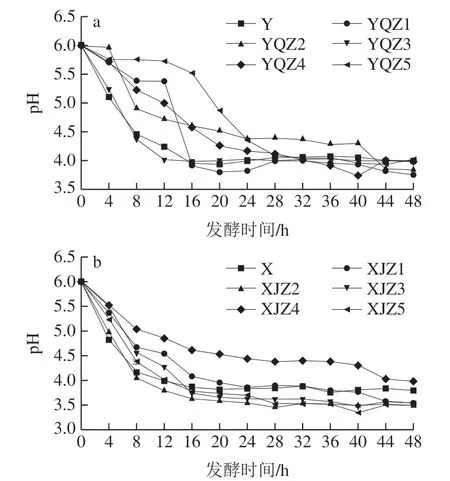
圖6 產酸菌株Y組(a)、X組(b)的pH值變化趨勢Fig. 6 pH trends of acid-producing strains from group Y (a) and group X (b)
產酸能力是衡量產酸菌發酵活力的重要指標,決定著發酵周期的長短。發酵過程產生的有機酸不僅抑制部分有害微生物的生長繁殖[21],而且對酸漿風味形成及酸漿豆腐的品質和風味均有重要影響[22]。僅X組菌株XJZ2產酸量高于50 g/L。Y組菌株YQZ1、YQZ2和X組菌株XJZ3的產酸量在40~50 g/L之間。Y、X組產酸量平均值為34.46、46.41 g/L,均高于葉青[23]優化后的最高產酸量34.25 g/L,2 組菌株從整體上表現較強的產酸能力,具有深入研究價值。
2.2.3 產酸菌耐酸性能分析
采用一定酸度的酸漿水點漿是大豆蛋白是否能夠酸凝、豆腐壓榨成型后能否保持一定品質的關鍵,因此產酸菌應該具有一定耐酸能力以保證酸漿酸度達到點漿要求及后續工藝順利進行。
如圖7所示,隨pH值下降,菌體濃度呈現不同幅度的降低,相同pH值表現不同的耐受能力。自然菌群在pH 3.5~4.5的最佳點漿酸度范圍內[24-25],相較于其他菌株表現較為脆弱的生長趨勢,自然菌群Y、X的活菌數下降幅度明顯,其原因可能為自然菌群中的大部分耐酸能力較差的菌群隨酸度下降的生長受到明顯抑制,因此整體體現較差的耐酸性。從Y組可知,菌株YQZ3、YQZ5的耐酸性最強,pH 2.5的菌體濃度仍保持在1.64、1.39的較高水平。菌株YQZ1在pH 4.0時生長狀況保持良好,培養24 h后,菌體濃度為1.47。當pH值降至3.0時,YQZ1、YQZ2、YQZ4菌體濃度下降明顯,pH值低于2.5則幾乎不生長,其原因可能為pH值過低,環境中的氫離子濃度過高,菌株無法維持細胞內的電荷平衡,導致生長受到抑制,甚至死亡。從X組可知,pH值降至3.5前,XJZ2菌株耐酸能力始終高于其他菌株,pH 3.5時仍能保持最高繁殖能力,XJZ4菌株耐酸性最差,當pH值降至4.0時,菌體濃度僅為0.31。因此菌株YQZ1、YQZ3、YQZ5、XJZ2具有較好的耐酸能力。
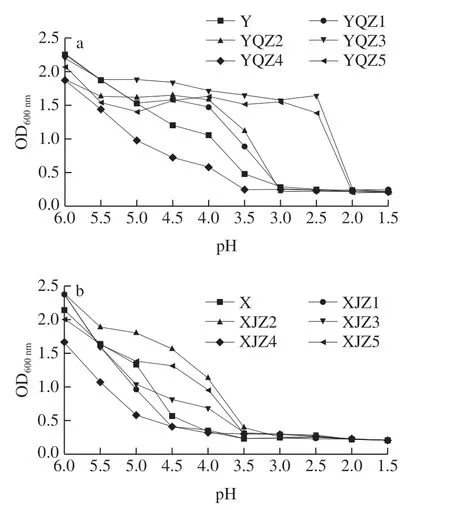
圖7 產酸菌株Y組(a)、X組(b)的耐酸性Fig. 7 Acid tolerance of acid-producing strains from group Y (a) and group X (b)
2.2.4 產酸菌耐滲透壓能力分析
微生物的生長繁殖需理想的滲透壓境條件,滲透壓過大會抑制微生物的生長繁殖,甚至脫水衰竭[26-27]。產酸菌株的耐鹽性結果如圖8所示。自然菌群雖具有一定豐度,各類微生物參與整個代謝環境,但隨著鹽質量濃度的增加,并未體現理想的耐受性和生存能力,NaCl質量濃度由0 g/100 mL增至4 g/100 mL時,活菌數大幅度下降,8 g/100 mL NaCl的菌落基本停止生長,其活菌數遠小于不含NaCl培養基內的活菌數。由Y組可知,菌株YQZ2、YQZ3耐受性最弱,4 g/100 mL NaCl或6 g/100 mL NaCl條件下,菌落的生命活動受到明顯抑制;NaCl質量濃度大于6 g/100 mL時,菌落無法生長而死亡。4 g/100 mL NaCl條件下菌株YQZ4生長良好,活菌數最高,培養24 h時,其OD600nm為1.53,當NaCl質量濃度大于4 g/100 mL時,生長受到明顯抑制,但在NaCl質量濃度為10 g/100 mL時仍能存活。X組菌株整體耐鹽性較差,菌株XJZ3、XJZ5對鹽的耐受性最差,4 g/100 mL NaCl條件下基本不生長。菌株XJZ2在NaCl質量濃度由4 g/100 mL增至8 g/100 mL時表現理想的耐鹽性,活菌數下降幅度小,8 g/100 mL NaCl條件下仍能夠存活。因此菌株YQZ4具有非常理想的耐鹽性。
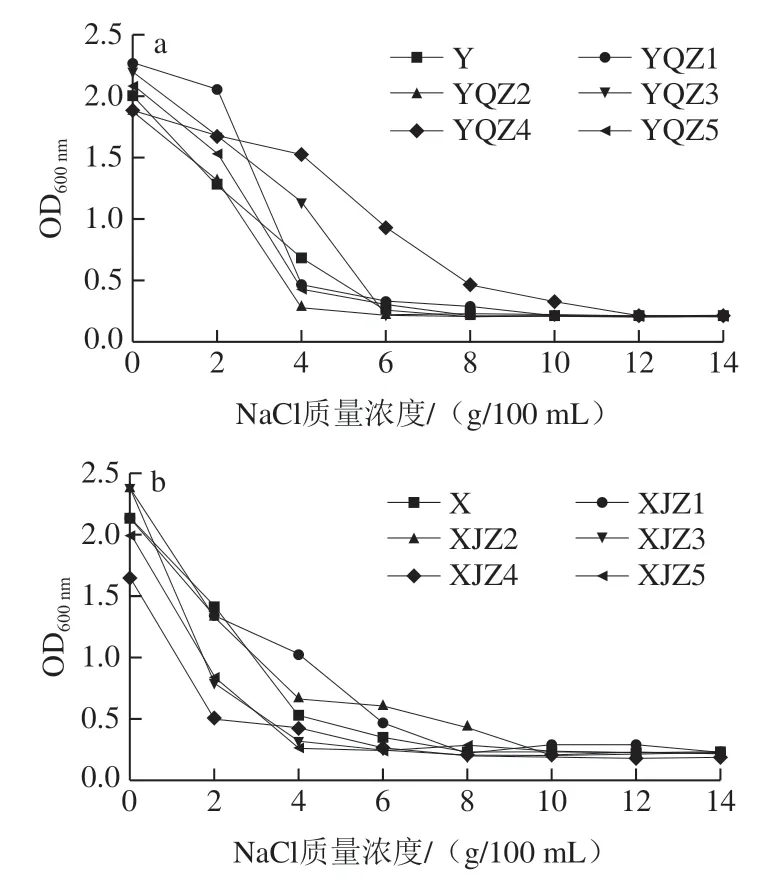
圖8 產酸菌株Y組(a)、X組(b)的耐鹽性Fig. 8 Salt tolerance of acid-producing strains from group Y (a) and group X (b)
3 討 論
乳酸菌是有重要生理功能的益生菌,廣泛分布于乳制品、腌制肉類、泡菜醬菜等各類發酵食品中,乳酸菌發酵不僅能夠產生酸乳酸、甲酸、乙酸、γ-氨基丁酸等賦予食物獨特的風味,而且大部分還能產生乳酸菌素,抑制甚至殺死許多導致食物腐敗、變質的致病菌,不影響食物的品質風味,且延長食物的貯藏保鮮期[28-29]。目前國內外對酸漿水中的優勢乳酸菌已有諸多研究[4,30-33]。Qiao Zhihong等[4]通過研究酸漿水中的不同有機酸含量得出乳酸菌為酸漿水中優勢菌群的結論。Xu Yunhe等[30]通過高通量測序及研究大豆蛋白凝固機理,獲得1 株凝乳能力強、產酸量大的干酪乳桿菌YQ336。Fei Yongtao等[31]優化酸水中分離獲得菌株的發酵條件,獲得酸度值為110 °T的淀粉乳桿菌L6。本研究經分離篩選鑒定、生長特性研究獲得產酸量高達50.91 g/L的菌株XJZ2,是佟梓沂等[32]從酸漿水中分離得到的植物乳桿菌培養48 h產酸量40.80 g/L的1.25 倍。分離獲得產酸量高達43.61 g/L的菌株YQZ1,其產酸性能強于賀云[33]從酸漿水分離得到的發酵乳桿菌。菌株XJZ2、YQZ1均有理想的產酸能力,均能大幅縮短酸漿水發酵周期、提高經濟效益、發揮潛在應用價值。
腸球菌常見于乳制品、發酵肉制品、腸道內及糞便中,酸漿水中鮮見報道[34-36]。M. oxydans多從土壤中分離獲得[37],酸漿水中未有報道。本研究從酸漿水中分離到3 株高產酸腸球菌,1 株微桿菌。菌株XJZ1(E. faecium)產酸量49.90 g/L,耐酸性較理想的菌株YQZ5(M. oxydans)產酸量31.59 g/L。本研究證明酸漿水中的優勢菌群除乳酸菌外,還有其他產酸性能力強的菌群,其均對增加酸漿水酸度發揮重要作用,對酸漿水風味的形成可能具有重要影響。因此還需進一步研究3 株腸球菌、M. oxydans與乳酸菌協同發酵黃漿水的代謝機制、對人體有潛在危害代謝物、具有重要生理活性功能的代謝物及其對酸漿水風味的影響,以為改善酸漿豆腐品質、口感提供理論參考。
4 結 論
為探明云南牟定酸漿水中優勢產酸菌株及其生長特性、高效應用黃漿水、減少排放污染,本實驗從云南牟定A、B公司自然發酵酸漿水中分離篩選到10 株高產酸菌株,并對其進行形態學觀察、生理生化實驗、16S rDNA序列測定分析、同源性比對及發育樹的構建。通過形態學觀察、生理生化實驗,可將10 株菌可分乳桿菌屬、乳球菌屬、腸球菌屬、微桿菌屬。將10 株菌進行16S rDNA序列測定分析及同源性比對得到菌株YQZ1為L. fermentum、菌株YQZ2為L. raffinolactis、菌株YQZ3為L. plantarum、菌株YQZ4為E. casseliflavus、菌株YQZ5為M. oxydans、菌株XJZ1為E. faecium、菌株XJZ2為L. plantarum、菌株XJZ3為L. paracasei、菌株XJZ4為 E. hirae、菌株XJZ5為L. paracasei。10 株高產酸菌種乳酸菌6 株、腸球菌3 株、微桿菌1 株,因此可確定乳酸菌為云南牟定酸漿水中優勢菌群。
10 株菌及自然菌群的生長溫度、生長曲線、產酸能力、耐酸能力的分析結果表明,菌株XJZ4最適生長溫度為32 ℃,其余菌株的最適生長溫度均為37 ℃。相同培養時間內,菌株XJZ2生長速率最大,菌株YQZ1、XJZ2繁殖能力最強;產菌株YQZ1、XJZ2產酸能力最強,有機酸累積量最多,培養至20 h時pH值分別降至3.80、3.58,二者于48 h產酸量分別高達43.61、50.91 g/L;菌株YQZ3、YQZ5的耐酸性最強;菌株YQZ1、XJZ2耐酸性良好,在pH 4.0時仍能保持菌體濃度高于1.00;菌株YQZ4耐鹽性最強,菌株YQZ4在4 g/100 mL NaCl條件下仍能保持菌體濃度1.53,其余菌株在NaCl質量濃度大于4 g/100 mL時生長受到明顯抑制;通過生長特性分析,菌株YQZ1、XJZ2均具有較強的繁殖能力和較高的產酸性能,二者可用于后續混合菌種發酵黃漿水研究。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
