海拔和坡向對唐古拉山土壤胞外酶活性的影響
2021-10-29 03:22:38彭子洋劉衛星黃俊勝楊雅舒劉玲莉
生態學報 2021年19期
彭子洋,劉衛星,田 瑞,楊 森,王 靜,黃俊勝,楊雅舒,劉玲莉
1 中國科學院植物研究所植被與環境變化國家重點實驗室,北京 100093 2 中國科學院大學,北京 100049 3 蘭州大學,蘭州 730000 4 河北大學,保定 071002 5 山東農業大學,泰安 271018
土壤胞外酶是土壤有機質的降解、轉化和礦化的媒介[1],能夠將大分子有機質分解成可以被微生物利用的小分子物質[2],是分解過程中重要的限速因子,在陸地生態系統生物地球化學循環中起著重要的作用。土壤胞外酶活性會受到氣候(溫度、降水、太陽輻射)、地理(地形、土壤)、生物(植物、微生物)等多種因素的影響,但各因子對胞外酶活性的影響尚未得到統一結論。由于長期的動植物殘體積累,青藏高原具有巨大的碳庫儲量,且該區域具有豐富的山脈,氣候、地理、生物等環境變化幅度大,生態系統脆弱,對環境變化十分敏感,因此選取該區域中典型的山地生態系統,研究其土壤胞外酶活性沿海拔和坡向變化的空間分布格局及調控因素,對于解析該區域土壤碳循環對氣候變化的響應具有重要意義。
海拔和坡向的改變可能不會對微生物和土壤胞外酶活性產生直接影響[3],但是二者的變化會改變溫度、降水、光照等環境因子[4-5]。在阿爾卑斯山脈和念青唐古拉山進行的多個研究表明,土壤酶活性隨海拔梯度的升高而降低[6- 9]。根據阿倫尼烏斯(Arrhenius)公式,溫度是影響酶活性的重要因子,低溫可減少酶與底物碰撞的次數,降低酶的反應速率,因此一些研究認為低溫可能是導致高海拔地區土壤酶活性較低的主要原因。同時海拔梯度和坡向的不同也改變了土壤水分和光照[10],從而影響土壤酶活性。不同生態系統中土壤胞外酶活性對土壤濕度的響應不同[6,11- 12]。在土壤水分較高苔原、草甸生態系統中,過高的土壤濕度所導致的厭氧環境通常會抑制土壤酶活性[13];而在干旱半干旱草地生態系統中,較高的水分有利于底物和酶的擴散,提高底物的可接觸性,土壤濕度的增加會提高土壤胞外酶的活性[14-16]。因此,不同海拔和坡向間氣候條件的差異可能對土壤酶活性產生不同影響。
海拔和坡向的改變還可能通過影響植被生長和微生物碳源,間接影響土壤微生物[3]和土壤酶活性,進而調控土壤碳循環和養分循環[17-19],有研究發現,空氣溫度、光合有效輻射、土壤溫度、含水量、土壤碳氮含量、水溶性酚等微氣候和土壤性狀在南北坡均具有顯著差異,南坡的土壤有機質、微生物量和土壤胞外酶活性均高于北坡[20]。但同時也有研究發現,受土壤發育的影響,北坡具有更高的土壤有機碳和微生物量,使得土壤胞外酶活性高于南坡[21]。植物凋落物及根際分泌物是土壤碳的主要來源,土壤碳和土壤養分作為土壤胞外酶作用的底物,其含量與組成也會影響胞外酶活性[22]。米氏方程表明,酶活性會隨底物濃度的升高而升高,并最終達到飽和。大量研究也發現,β-葡萄糖苷酶(BG)、纖維二糖水解酶(CBH)、N-乙酰-氨基葡萄糖苷酶(NAG)活性隨土壤碳、氮含量的增加而增加[3]。較高的土壤養分有利于植物和微生物的生長,從而產生更多的土壤水解酶。植物生物量和群落結構的改變也會影響微生物群落結構和功能,進而影響土壤酶活性[23]。由此可見,目前的研究對土壤胞外酶活性沿海拔梯度的垂直分布格局以及不同坡面的水平分布格局并無統一定論,海拔和坡向對土壤胞外酶活性的影響及其主要驅動因子仍需進一步研究。
青藏高原碳庫儲量巨大,山脈資源豐富,生態系統脆弱,對環境變化十分敏感。唐古拉山是青藏高原的典型山脈,具有很強的垂直地帶性[24],生物、非生物環境條件沿海拔梯度的變化顯著[25]。同時,唐古拉山還阻礙了印度季風向歐亞大陸的傳播,形成強烈的雨影效應[26],顯著改變了兩側坡面的氣候條件[27]。為了探究唐古拉山南北坡土壤胞外酶活性沿海拔梯度的分布格局及其主要驅動因素,進一步理解青藏高原土壤有機質周轉過程,我們沿唐古拉山南北坡不同海拔梯度(2980—5120 m)各選取了3個點,測定土壤胞外酶潛在活性、土壤理化性質等指標,擬解決以下科學問題:1)唐古拉山土壤胞外酶潛在活性沿海拔和坡向的分布規律;2)形成該分布格局的主要驅動因子是什么?
1 材料和方法
1.1 研究區概況
為探究海拔和坡向對土壤胞外酶活性的影響,本研究沿唐古拉山脈的南坡(91.02—91.7°E,30.42—32.38°N,海拔4250—5120 m)和北坡(92.6—97.68°E,34.38—36.23°N,海拔2980—4830 m)各選取了3個點進行了取樣(表1)。取樣區域植被類型為高寒草原草地[28],輻射強烈,溫度隨海拔和緯度的升高而降低,日較差大,季節差異明顯。降水主要受西南季風控制,自東南向西北方向逐漸減少;降水季節分配極不均勻,雨季和旱季分異明顯。

表1 研究樣地基本地形和氣候特征
1.2 樣品采集
青藏高原的植被生產力在每年的8月達到最大值,此時的土壤微生物活性最強,能夠更好地表征土壤酶活性最大潛力。因此,我們于2016年8月沿唐古拉山脈不同海拔和坡向進行土壤樣品采集和植被調查,在每個海拔梯度隨機選取3個1m×1m的樣方,每個樣方內隨機選擇1個采樣點,用直徑9 cm的土鉆采集0—5 cm混合土壤樣品,一共采集18個新鮮土樣。所取得的新鮮土樣采用2 mm篩去除粗根系和石子,混合均勻后置于冷藏箱內迅速運回實驗室分為兩份,一份儲藏在-80 ℃條件下用于土壤胞外酶活性和水分的測定,另外一份風干后用于土壤理化性狀的測定。過篩后的根系也置于便攜式冷藏箱內,隨土壤一起寄回實驗室,清洗后60 ℃烘干48 h至恒重,稱取干重并換算成0—5 cm單位面積的根系生物量(Root biomass, RB)。
1.3 樣品測定
樣品前處理:在實驗室測定前首先剔除了土壤樣品中的細根及可見的凋落物碎屑。
土壤含水量:稱取一定質量m1的土壤樣品鮮樣在105℃條件下烘干48h至恒重,然后稱量樣品干重m2,土壤含水量(Soil Moisture, SM)=(m1-m2)/m2×100%。
土壤碳(TC)、氮(TN)含量測定:采用燃燒法測定土壤C、N含量。土壤于避光通風處風干后研磨粉碎,混合均勻后用元素分析儀(Vario EL Ⅲ, Elementar,Germany)測定土壤C、N含量。
土壤胞外酶活性:本研究測定6種常見土壤胞外酶潛在活性,其中碳獲取酶有3種,分別是α-葡萄糖苷酶(α-D-Glucosidase, AG)、β-葡萄糖苷酶(β-D-Glucosidase, BG)、β-木糖苷酶(β-Xylosidase, BX);氮獲取酶有兩種,分別是亮氨酸氨基肽酶(Leucine Aminopeptidase, LAP)、N-乙酰-氨基葡萄糖苷酶(N-Acety1-β-D-Glucosaminidase, NAG);磷獲取酶為酸性磷酸酶(Acid Phosphatse, AP)(表2)。土壤胞外酶活性采用96微孔酶標板熒光分析法[29],使用多功能酶標儀(SynergyH1, BioTek, United States)進行測定。具體方法如下:稱取0.3000g新鮮土壤樣品于離心管中,加入60mL馬來酸鈉緩沖液(pH=6),超聲細胞粉碎機超聲2 min,制備成土壤懸濁液。按順序將125 μL AG、BG、BX、AP、LAP、NAG 6種胞外酶對應的底物、7-氨基- 4-甲基香豆素(7-Amino- 4-methylcoumarin, AMC)標準物、4-甲基傘形酮(4-Methylumbelliferone, MUB)標準物、超純水分別加入96孔酶標版的相應孔洞中;將125 μL緩沖液、土壤懸濁液分別加入對照板和樣品板的相應孔洞中;25℃避光培養4 h后,在360 nm 波長處激發,460 nm 波長發射的條件下測定熒光。

表2 6種土壤胞外胞外酶種類
1.4 數據計算與處理
1.4.1氣象及植被數據來源
年均溫(MAT)、年均降水(MAP)、紫外線輻射(UV)數據來自于中國區域地面氣象要素數據集(China Meteorological Forcing Dataset)2016年的數據,該數據的空間分辨率為0.01°×0.01°[30- 31]。歸一化植被指數(NDVI)數據來源于中分辨率成像光譜儀(MODIS)數據中MOD13C1 Collection 6的每月NDVI數據,其空間分辨率為0.25°×0.25°[32]。為保證結果的準確性與可信度,只采用質量較好的數據用于后續分析。
1.4.2土壤胞外酶活性計算公式
(1)
式中,EEA為胞外酶活性,單位:nmol g-1h-1;NFU為單位凈熒光值;ε為發射系數(Emission Coefficient),單位:Fluorescence/nmol;VH為土壤懸濁液體積,單位:mL;T為培養時間,單位:h;mS為土壤重量,單位:g;VB為用于制備土壤懸濁液的緩沖液體積,單位:mL。
其中:
(2)
(3)
(4)
式中,NFU為單位凈熒光值;FA為樣品板(底物+土壤懸濁液)的熒光值;FHC為樣品對照(土壤懸濁液+緩沖液)的熒光值;FSC為底物對照(底物+緩沖液)的熒光值;Q為淬滅系數;FQ為土壤懸濁液+標準物的熒光值;FS為緩沖液+標準物的熒光值;ε為發射系數,單位:Fluorescence/nmol;CS為標準物濃度,單位:nmol/mL;VS為標準物體積,單位:mL。
1.4.3數據處理方法
本研究采用二因素方差分析(Two-Way ANOVA)來比較海拔、坡向及其交互作用對根生物量、土壤濕度、土壤碳、土壤氮和6種土壤胞外酶活性的影響,顯著性水平為P<0.05。
本研究采用基于Step-AIC的模型參數篩選,并通過廣義線性混合模型的方法分析地形(海拔)、氣象(MAT、MAP、UV)、生物(NDVI、RB)、土壤(TC、TN、SM、C∶N)等因子對土壤胞外酶活性的作用大小。將各潛在解釋因子標準化后,通過比較方差膨脹系數(VIF)檢驗全模型因子之間是否存在多重共線性,發現MAT、MAP、UV、TN具有較高的VIF,故將此4種潛在解釋因子從全模型中去除,保留了VIF<10的海拔、NDVI、RB、TC、C: N 、SM 6種潛在解釋因子。對該6種潛在解釋因子構建的多模型進行篩選時,通過R語言MuMin包中的“dredge”函數基于ΔAIC ≤ 10的所有模型進行模型平均[33]。
以上統計分析均使用R語言(R version 3.5.3)實現,用到的包為MASS、lme4、car、MuMIn。
2 結果
2.1 氣候、植被和土壤性狀沿海拔和坡向的分布
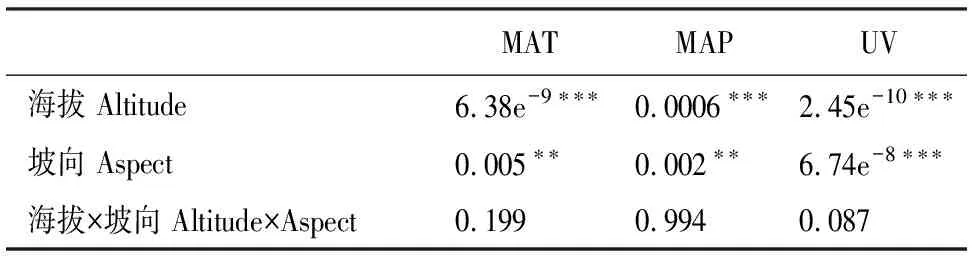
表3 海拔、坡向及其交互作用對環境因子的影響
不同海拔梯度的生物、非生物因子環境具有顯著差異(表3、圖1)。隨海拔梯度的升高,MAP、UV、NDVI、RB、SM、TC、TN均顯著升高(P<0.05),而MAT、C: N隨海拔升高而降低(P<0.05)。
南坡和北坡的氣候、植被和土壤性狀也具有顯著差別(表3、圖1)。實驗區域內南坡的MAP、UV、TC、TN均極顯著高于北坡(P<0.001),分別是北坡的1.6、1.1、2.5、4.7倍;但北坡的MAT、C∶N高于南坡。
NDVI在南坡隨海拔升高而下降,但在北坡隨海拔升高而升高;RB在南坡隨海拔升高而升高,在北坡沒有顯著變化趨勢;SM在南北坡沒有顯著差異,分別隨海拔升高而升高。
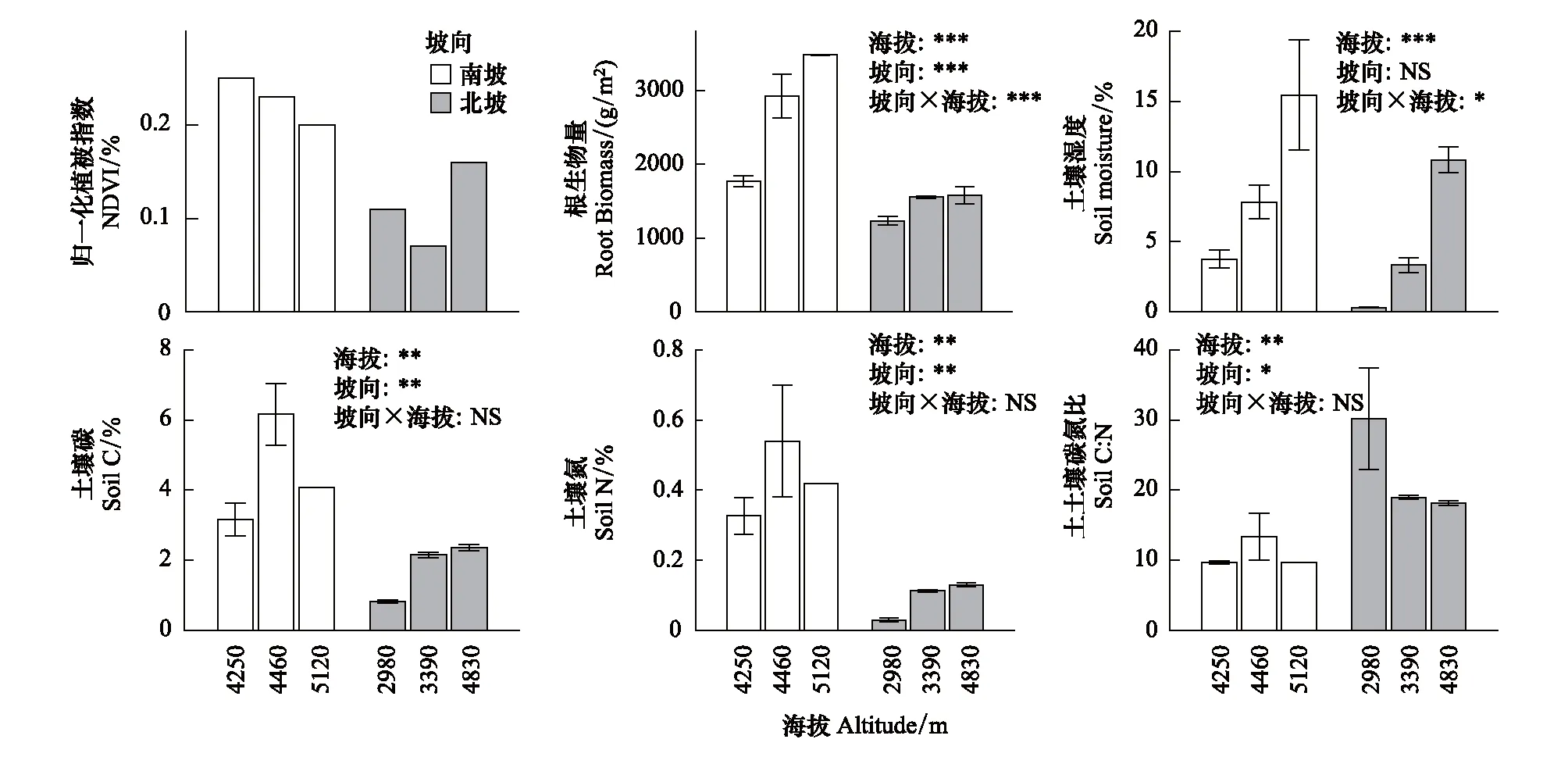
圖1 歸一化植被指數、根生物量、土壤濕度、土壤碳、土壤氮、碳氮比沿海拔和坡向的分布Fig.1 Distribution of NDVI, root biomass, soil moisture, soil carbon, soil nitrogen, C: N along altitudes and aspects白色柱子表征南坡,灰色柱子表征北坡;海拔、坡向及其交互作用對根生物量、土壤濕度、土壤碳、土壤氮、土壤碳氮比的二因素方差分析結果在每張圖的右上角標出(n=3); **, P<0.01; ***, P<0.001; NS, None Significant
2.2 6種土壤胞外酶活性沿海拔和坡向的分布
唐古拉山不同海拔AG、BG、BX、AP、LAP、NAG 6種胞外酶活性在120—3000 nmol g-1h-1之間(圖2)。碳獲取酶AG、BG、BX活性和氮獲取酶NAG活性隨海拔的升高而升高 (P<0.05);其他胞外酶與海拔并無顯著相關關系。
南坡的6種胞外酶活性在200—3000 nmol g-1h-1之間,北坡的6種酶活性在120—1600 nmol g-1h-1之間。碳獲取酶AG、BG、BX和氮獲取酶NAG活性在南坡要顯著高于北坡的酶活性(P<0.01)。磷獲取酶AP和氮獲取酶LAP活性在南北坡沒有顯著差異(圖2)。

圖2 6種土壤胞外酶活性沿海拔和坡向分布特征Fig.2 The activities of AG, BG, BX, AP, LAP and NAG in six plots along the altitudes and aspects 圖中分別為AG、BG、BX、AP、LAP、NAG沿海拔和坡向分布圖;白色柱子表征南坡胞外酶活性,灰色柱子表征北坡胞外酶活性(n=3);海拔、坡向及其交互作用對AG、BG、BX、AP、LAP、NAG的二因素方差分析結果在每張圖的右上角標出;AG:α-葡萄糖苷酶 α-D-Glucosidase; BG:β-葡萄糖苷酶β-D-Glucosidase; BX:β-木糖苷酶β-Xylosidase; AP:酸性磷酸酶 Acid Phosphatase; LAP:亮氨酸氨基肽酶 Leucine Aminopeptidase; NAG:N-乙酰-氨基葡萄糖苷酶 N-acety1-β-D-Glucosaminidase; **, P<0.01; ***, P<0.001; NS, None Significant
2.3 土壤胞外酶活性與氣候、植被、土壤理化性質的關系
不同海拔和坡向的氣候、植被、土壤等理化性質具有顯著差異,驅動不同土壤胞外酶活性變化的因子也不同。模型平均的結果表明,TC是影響碳獲取酶AG、BG、BX酶活性變化的主要驅動因子之一,對AG、BG、BX酶活性具有正效應(圖3)。NDVI和RB對碳獲取酶BG、BX酶活性也具有正效應,是驅動BG、BX酶活性變化的因子(圖3)。NDVI是磷獲取酶AP活性和氮獲取酶NAG活性變異最重要的驅動因子(圖3)。海拔、NDVI、RB、TC、C: N、SM對氮獲取酶LAP酶活性并沒有顯著的作用(圖3)。
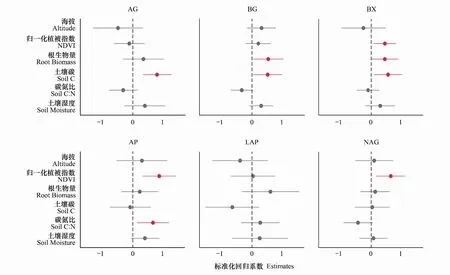
圖3 海拔、歸一化植被指數、根生物量、土壤碳、土壤碳氮比、土壤濕度對土壤胞外酶AG、BG、BX、AP、LAP和NAG活性分布的影響Fig.3 The effect of altitude, MAP, SM, TC, C: N on the activities of soil extracellular enzymes AG, BG, BX, AP, LAP and NAG圖中顯示了模型預測變量的平均參數估計(標準化回歸系數)和相關的95%置信區間(n =3)
3 討論
(1)海拔梯度和坡向對碳循環土壤胞外酶分布的影響
海拔梯度和坡向差異導致的溫度、降水、光照[34]等氣候因子的變化可能會使土壤胞外酶活性隨海拔高度和坡向呈現出一定的空間分布格局[14- 15, 35]。阿倫尼烏斯公式表明,在一定范圍內,溫度與酶活性成正相關。隨著海拔梯度的升高和北坡光照的減少,溫度降低[10],土壤酶活性降低。與此假設相反,我們的研究結果表明,碳獲取酶AG、BG、BX酶活性隨海拔升高而升高,且在南坡的酶活性顯著高于北坡。AG、BG、BX主要參與木糖、纖維素等物質的分解[36],Siles等人認為這幾種胞外酶活性與海拔成正相關可能是因為隨海拔升高,底物濃度上升對土壤酶活性的促進作用抵消了低溫造成的負效應[9]。我們的研究支持了這一觀點。本研究中,TC隨海拔升高而升高,且AG、BG、BX酶活性與TC顯著相關。分析發現,TC和是影響AG、BG、BX酶活性最主要的因子(圖3)。該分析結果與前人的研究一致[37-38],隨海拔的升高,溫度可能成為地上植被生長的主要限制因子,使得 NDVI 在南坡隨海拔升高而下降。地下根生物量則可能更多受到 SM 的影響,在高寒草地系統中,水分是一個重要的限制因子,隨海拔升高,土壤水分增加,緩解了水分對植物的限制作用,促進了根系的生長[39- 40],同時高海拔的低溫又限制了分解,有利于碳儲量的升高。而南坡的光照和水分條件均強于北坡,使得南坡NDVI和RB均高于北坡。而較高的NDVI和RB能夠促進凋落物和根際分泌物向土壤的輸入[41],TC升高,增加了酶作用的底物濃度,從而促進了土壤胞外酶活性[42-43]。
(2)海拔梯度和坡向對氮、磷循環土壤胞外酶分布的影響
NAG主要參與分解土壤中的幾丁質,被認為是主要的N素獲取酶之一[44]。分析結果表明,NDVI是驅動氮獲取酶NAG酶活性隨海拔梯度和坡向的分布格局最主要的因子。NDVI的提高表征了植物會向土壤輸入更多的新鮮凋落物,一方面,新鮮凋落物的輸入增加了NAG的作用底物,底物的增加可刺激酶促反應速率[16, 42- 43];另一方面,凋落物的輸入也促進了微生物的生長,使得微生物分泌出更多胞外酶,提高酶活性[36, 45]。LAP也是氮獲取酶,但是在海拔和坡向上并無顯著規律,可能是因為微生物群落改變或微生物對環境的適應維持了LAP活性的穩定[46]。
AP作用于土壤中的有機酸,將其分解為植物、微生物可利用的無機磷[47]。磷獲取酶AP在不同海拔和坡向上的分布并沒有顯著的規律性,但模型平均結果表明NDVI對AP有顯著影響,這可能是因為在3390 m處,受水熱條件或營養元素等因子的影響,NDVI和AP均遠低于其他五個樣地。而其他五個樣地的AP活性無顯著差異,這可能與該地區氮磷含量和分布有關。AP的合成需要大量的氮元素[48],雖然在采樣區域內氮分布隨海拔和坡向變化存在差異,但是由于采樣區域生態系統受氮限制,該差異可能并不足以引起AP活性變化;其次,因土壤總氮含量和變化并不能表征土壤有效氮含量和變化,需進一步分析該區域內有效氮含量與AP活性的關系;此外,AP活性與土壤磷含量成反比,當生態系統處于磷限制時,微生物會分泌大量AP來獲取更多的磷,AP活性升高[37],但磷可能不是采樣區域的主要限制因子,所以AP活性沿海拔梯度和坡向未存在顯著差異。
4 結論
本研究探討了參與碳、氮、磷循環的土壤胞外酶活性沿唐古拉山不同海拔和坡向的分布格局及其主要驅動因子。研究發現受到TC含量的驅動,碳獲取酶AG、BG、BX活性沿海拔梯度升高,且南坡的活性高于北坡的活性;氮獲取酶NAG活性在南坡也高于北坡,主要受NDVI的調控;而磷獲取酶胞外酶 AP和氮獲取酶另一種胞外酶LAP活性在不同海拔梯度和坡向上并未表現出顯著差異,可能需要進一步的研究。本研究結果表明,參與碳、氮循環的土壤胞外酶活性在唐古拉山南北坡不同海拔梯度上具有較大差異,可能會影響未來全球變化條件下本區域碳循環和營養循環過程,為預測土壤胞外酶的空間地理格局提供了科學依據。
