長鏈非編碼RNA小核仁RNA宿主基因1靶向miR-330促進胰腺癌細胞吉西他濱耐藥作用
2021-10-26 08:33:32李銳祝喜萍宋蕾
中華胰腺病雜志 2021年5期
關鍵詞:耐藥
李銳 祝喜萍 宋蕾
黑龍江省醫院消化病院消化三科,哈爾濱 150086
化療耐藥是導致胰腺癌高病死率的重要原因之一[1-2]。隨著化療藥物的長期使用,機體自身的免疫系統產生耐藥性,化療藥物對于腫瘤細胞的殺滅力度明顯降低,影響患者的生存時間[3]。長鏈非編碼RNA(long noncoding RNA, lncRNA)是一種長度大于200nt的非編碼RNA,主要通過內源性競爭RNA的作用方式參與多種疾病的病理生理過程。小核仁RNA宿主基因(small nucleolar RNA host genes,SNHGs)是lncRNA的一個亞群,有報道其成員SNHG1、SNHG 6異常表達參與多種腫瘤細胞生物學過程[4-7]。miR-330是SNHG1的下游靶基因之一,現有的研究表明miR-330在膠質瘤[8]、結直腸癌[9]以及肺癌[10]等癌細胞中低表達。吉西他濱(gemcitabine, GEM)為胰腺癌的一線化療藥物[11-13],目前關于SNHG1和miR-330在胰腺癌GEM耐藥中的作用未見報道。本研究探討SNHG1對胰腺癌GEM耐藥細胞增殖活性以及侵襲遷移能力的影響,驗證SNHG1與miR-330的靶向關系,闡明SNHG1/miR-330信號軸在胰腺癌GEM耐藥中的作用。
資料與方法
一、胰腺癌組織及癌旁正常胰腺組織lncRNA SNHG1和miR-330表達檢測
收集癌癥基因組圖譜(The Cancer Genome Atlas, TCGA)胰腺癌數據集的179例胰腺癌組織和171例癌旁正常胰腺組織的臨床數據,通過GEOtoR軟件分析胰腺癌組織及癌旁正常胰腺組織lncRNA SNHG1和miR-330的表達。
二、GEM耐藥的胰腺癌PANC1細胞株的構建
人胰腺癌PANC1細胞株購自中國科學院上海生命科學研究院細胞資源中心,常規復蘇、傳代。體外采用間歇梯度倍增法[14]用GEM刺激PANC1細胞24周,建立GEM耐藥PANC1細胞株(耐藥株)。預實驗結果顯示100 μmol/L的GEM刺激效果最佳,故以下實驗均采用100 μmol/L的GEM刺激獲得耐藥細胞。
取PANC1耐藥細胞,使用Lipofectamine 3000(Life Technologies,美國)分別將陰性對照siRNA(si-NC)、靶向lncRNA SNHG1的siRNA(si-SNHG1)轉染耐藥細胞,并運用Geneticin(Sigma-Aldrich,美國)進行篩選,建立穩定的si-NC耐藥株、si-SNHG1耐藥株及單純耐藥株。此外,為了驗證SNHG1對miR-330的調控作用,分別將陰性對照抑制劑(NC-inhibitor,NC-inh)和si-NC、NC-inh和si-SNHG1、miR-330-inh和si-SNHG1共同轉染耐藥細胞,建立了穩定的NC-inh+si-NC耐藥株、NC-inh+si-SNHG1耐藥株及miR-330-inh+si-SNHG1耐藥株。所有的miRNA、siRNA以及陰性對照均購自廣州銳博生物技術有限公司。
三、GEM耐藥PANC1細胞的lncRNA SNHG1和miR-330表達檢測
取3組對數生長期PANC1耐藥細胞,采用TRIzol(Life Technologies,美國)法提取細胞總RNA,運用PrimeScript RT試劑盒(TaKaRa,日本)將總RNA逆轉錄成cDNA,上Bio-Rad-CFX-qRT-PCR檢測系統(Applied Biosystems Inc.,美國),使用SYBR Green Master(ROX;Roche,加拿大)進行實時熒光定量PCR反應。SNHG1 正向引物序列為5′-TGCAATGTTCAGCCCACAAG-3′,反向引物序列為5′-GCA-GCTGAATTCCCCAGGATA-3′;miR-330 正向引物序列為5′-TCTCTGGGCCTGTGTCTTAG-3′,反向引物序列為5′-CAGTGCGTGTCGTGGAGT-3′,內參U6正向引物序列為5′-GTGCTCGCTTCGGCAGCACATATAC-3′,反向引物序列為5′-AAAAATATGGAACGCTTCACGAATTTG-3′;內參GAPDH正向引物序列為5′-GACCCCTTCATTGACCTCAAC-3′,反向引物序列為5′-CTTCTCCATGGTGGTGAAGA-3′。通過儀器自帶軟件獲取擴增產物的Ct值,采用公式2-ΔΔCt計算其相對表達量,每個樣品設3個復管,取均值。
四、GEM耐藥PANC1細胞增殖活性檢測
選用3~6代si-NC耐藥組、si-SNHG1耐藥組、單純耐藥組、NC inh+si-NC耐藥組、NC inh+si-SNHG1耐藥組以及miR-330 inh+si-SNHG1耐藥組的對數生長期PANC1細胞,以每孔2×103個細胞種植于96孔板,加入GEM(100 μmol/L)孵育48 h,各孔加入10 μl CCK-8染料(中國碧云天公司),置37℃繼續孵育4 h,上酶標儀測量波長450 nm處每孔的吸光度值(A450值)。
取各組1×106個細胞,均勻鋪在含有無菌玻片的6孔板中,加入GEM(100 μmol/L)繼續孵育48 h,4%甲醛固定30 min,隨后加入Ki67抗體(沈陽萬類生物有限公司),按照免疫熒光操作流程進行熒光染色,置熒光顯微鏡下(40倍)觀察細胞熒光,用Imagej軟件定量細胞內Ki67的熒光強度。
五、GEM耐藥PANC1細胞侵襲能力檢測
采用Transwell小室檢測細胞的侵襲能力。Transwell小室隔膜上表面加50 μl基質凝膠過夜。取上述各組對數生長期PANC1細胞,調整細胞密度為1×105/ml,Transwell上室加100 μl細胞懸液,下室加500 μl含10%胎牛血清的RPMI1640細胞培養液,培養48 h后取出小室的隔膜,用棉簽輕輕擦去未穿膜細胞,乙醇固定隔膜5 min,1%的結晶紫染色30 min,置倒置相差顯微鏡下隨機取5個200倍視野計數穿膜細胞數。每組做6個Transwell小室,取均值。
六、GEM耐藥PANC1細胞遷移能力檢測
采用細胞劃痕實驗檢測細胞遷移能力。將各組1×106個/ml細胞接種到6孔板,培養過夜形成單層細胞時,使用10 μl的槍頭垂直于孔板底部劃3條直線,間隔0.5 cm,用PBS洗滌3次,加入無血清培養基培養24 h,置倒置相差顯微鏡下對劃痕前后的相應區域拍照。每組設3個復孔,重復3次。使用Image J軟件計算各組細胞遷移24 h后的遷移距離。
七、lncRNA SNHG1與miR-330的靶向關系驗證
通過Starbase 3.0數據庫搜索lncRNA SNHG1的潛在靶基因。生物信息學研究顯示lncRNA SNHG1和miR-330存在13個互補堿基(圖1),在此基礎上設計野生型lncRNA SNHG1(SNHG1-WT)、突變型lncRNA SNHG1(SNHG1-MT)、野生型miR-330(miR-330-WT)和缺失miR-330的突變型miR-330(miR-330-MT)。熒光素報告酶報告載體購自美國Life Technologies公司。分別將SNHG1-WT、SNHG1-MT、miR-330-WT、miR-330-MT熒光素酶報告載體通過lipofectamine 3000轉染到GEM耐藥的PANC1細胞中。其中, lncRNA SNHG1以及miR-330中野生型與突變型序列分別在結合位點處納入pMIR報告熒光素酶含有SpeI和HindⅢ載體位點(Ambion,USA)。對于未改變結合位點的lncRNA SNHG1以及miR-330指定為SNHG1-WT和miR-330-WT,結合位點改變的lncRNA SNHG1以及miR-330指定為SNHG1-MT和miR-330-MT。然后,將lncRNA SNHG1以及miR-330分別轉染GEM耐藥PANC1細胞24 h,采用雙熒光素酶報告基因檢測試劑盒(Promega,美國)檢測熒光素酶活性。
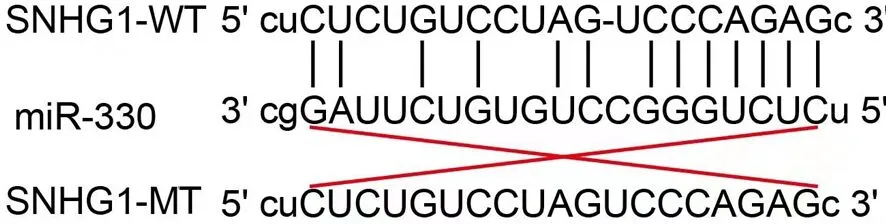
圖1 lncRNA SNHG1與miR-330相互作用的核苷酸結合位點信息
八、統計學處理
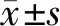
結 果
一、胰腺癌組織和各組GEM耐藥PANC1細胞lncRNA SNHG1和miR-330表達水平
胰腺癌組織及其癌旁正常胰腺組織lncRNA SNHG1的表達量分別為6.54±0.72、5.46±0.54,癌組織顯著高于癌旁正常組織,差異有統計學意義(t=6.00,P<0.01)。胰腺癌組織及其癌旁正常胰腺組織miR-330的表達量分別為2.54±1.85和3.01±1.23,癌組織明顯低于癌旁正常組織,差異有統計學意義(t=2.73,P<0.05)。
si-NC耐藥組、si-SNHG1耐藥組、單純耐藥組細胞的lncRNA SNHG1表達量分別為2.43±0.10、0.26±0.08、3.25±0.31,si-SNHG1耐藥組顯著低于其他兩組(t=1.21,P<0.05),表明抑制lncRNA SNHG1表達的耐藥細胞株構建成功。si-NC耐藥組、si-SNHG1耐藥組、單純耐藥組細胞miR-330的表達量分別為0.47±0.13、1.84±0.12、0.38±0.21,si-SNHG1耐藥組顯著高于其他兩組,提示抑制SNHG1表達能上調耐藥細胞miR-330的表達。
二、抑制SNHG1表達對GEM耐藥PANC1細胞增殖活性及侵襲、遷移能力的影響
與si-NC耐藥組、單純耐藥組比較,si-SNHG1耐藥組細胞A450值、Ki67熒光強度、穿膜細胞數、遷移距離均顯著降低,差異均有統計學意義,而si-NC耐藥組與單純耐藥組細胞的上述指標差異均無統計學意義(圖2,表1)。提示抑制SNHG1表達能夠抑制GEM耐藥PANC1細胞的增殖活性及侵襲、遷移能力。
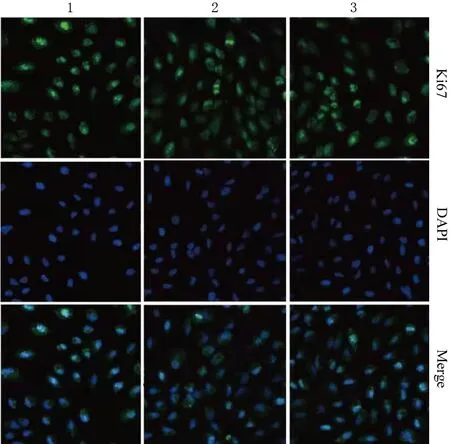
圖2 單純耐藥組(1)、si-NC耐藥組(2)、si-SNHG1耐藥組(3)細胞Ki67熒光強度

表1 單純耐藥組、si-NC耐藥組及si-SNHG1耐藥組細胞增殖活性、Ki67強度、侵襲及遷移能力的比較
三、lncRNA SNHG1靶向調控miR-330
si-SNHG1耐藥組細胞的miR-330-WT熒光素酶活性較si-NC耐藥組顯著升高(3.21±0.22比1.03±0.18,t=2.976,P<0.05),而si-SNHG1耐藥組與si-NC耐藥組細胞的miR-330-MT熒光素酶活性的差異無統計學意義(1.03±0.28比0.98±0.24,t=1.47,P>0.05);miR-330-inh耐藥組細胞的SNHG1-WT熒光素酶活性顯著低于NC-inh耐藥組(0.97±0.21比2.32±0.17,t=2.14,P<0.05),而miR-330-inh耐藥組與NC-inh耐藥組細胞的SNHG1-MT熒光素酶活性的差異無統計學意義(0.99±0.14比1.05±0.14,t=1.01,P>0.05),提示lncRNA SNHG1和miR-330之間可以靶向結合。
四、抑制miR-330表達可逆轉si-SNHG1對GEM耐藥PANC1細胞抑制增殖以及侵襲遷移的作用
NC-inh+si-SNHG1組細胞A450值、Ki67熒光強度、穿膜細胞數、遷移距離較NC-inh+si-NC組均明顯降低,而miR-330-inh+si-SNHG1組較NC-inh+si-SNHG1組均顯著升高,差異均有統計學意義(P值均<0.05,表2),提示抑制miR-330表達能夠逆轉由于SNHG1低表達抑制的GEM耐藥PANC1細胞的增殖活性及侵襲、遷移能力。
表2 NC-inh+si-NC、NC-inh+si-SNHG1組及miR-330-inh+si-SNHG1組的細胞活性、Ki67熒光強度、侵襲及遷移能力比較
討 論
lncRNA是一種“新型”的非編碼RNA,其長度在200~2 000個核苷酸序列。lncRNA并不能編碼蛋白故往往被認為是一種翻譯過程中的“噪音”而未受到重視。然而,現有報道證明lncRNA通過多種作用機制參與腫瘤的惡性進展以及化療藥物耐藥[15],尤其參與胰腺癌GEM耐藥的作用[16]。lncRNA SNHG1作為原癌基因廣泛參與多種癌癥的病理生理過程,如結直腸癌、肝癌、肺癌、前列腺癌、胃癌、食管癌及骨肉瘤等[17]。本研究結果顯示,SNHG1首先在胰腺癌腫瘤組織中高表達,而低表達SNHG1可體外抑制胰腺癌PANC1 GEM耐藥細胞株的細胞增殖活性以及侵襲遷移,這種作用是通過靶向調控SNHG1下游的miR-330實現的。
內源性競爭RNA(competing endogenous RNAs, ceRNA)揭示了一種RNA間相互作用的新機制。ceRNA尤其是在lncRNA參與癌癥細胞化療藥物耐藥中扮演著重要的角色。Liu等[18]研究發現lncRNA HCP5通過吸附miR-214-3p釋放其對于HDGF靶向作用從而調節胰腺癌GEM耐藥性。Xu等[19]證明,lncRNA DLEU2L可以靶向調控miR-210-3p/乳腺癌易感基因2信號軸抑制胰腺癌細胞對GEM的耐藥性。本研究結果發現,在GEM耐藥的PANC1細胞中SNHG1表達明顯升高,抑制SNHG1后GEM耐藥的PANC1細胞增殖活性以及侵襲遷移能力明顯降低。同時,低表達SNHG1后可以靶向調控GEM耐藥PANC1細胞中miR-330的表達。而miR-330往往作為一種抑癌因子以及ceRNA的關鍵作用靶點參與多種癌癥的惡性進展。miR-330作為ceRNA的調控因子參與circHIPK3調控RAS關聯域家族的表達促進胰腺癌細胞對GEM耐藥[20]。本研究結果顯示低表達miR-330能夠使SNHG1-WT的表達升高,而對于SNHG1-MT并未有影響,說明miR-330是SNHG1的靶基因。以上的結果說明SNHG1可靶向調控miR-330的表達,參與胰腺癌細胞的GEM耐藥。
GEM耐藥PANC1細胞共轉染si-SNHG1以及miR-330 inhibitor可逆轉由si-SNHG1誘導的GEM耐藥PANC1細胞增殖活性以及侵襲能力降低。筆者假設這種作用可能是由于SNHG1靶向結合miR-330后釋放了miR-330對下游因子的吸附作用,增強了SNHG1對于胰腺癌GEM耐藥細胞活性作用,為SNHG1/miR-330信號軸成為胰腺癌GEM耐藥的潛在治療靶點提供理論支持。
利益沖突所有作者均聲明不存在利益沖突
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
