乙酸-亞硫酸鹽兩步預處理對楊木木質素吸附纖維素酶的影響
2021-10-20 08:36:40王金葉賈麗麗徐勇張軍華
林業工程學報 2021年5期
王金葉,賈麗麗,徐勇,張軍華,*
(1. 西北農林科技大學林學院,陜西 楊凌 712100;2. 南京林業大學化學工程學院,南京 210037)
楊樹生長迅速,分布廣泛,是我國最主要的造林樹種。在楊木生產過程中所產生的刨花鋸末等廢棄物是一種優良的木質纖維材料,其可以通過纖維素酶水解糖化進而利用微生物發酵制備生物燃料,可減少對化石燃料的依賴[1]。木質纖維素由纖維素、半纖維素和木質素所構成,半纖維素覆蓋在纖維素微纖絲表面,通過氫鍵相互連接,而木質素則通過共價鍵與半纖維素相連,形成穩定的疏水結構[2]。木質素是纖維素酶水解的主要障礙物,其覆蓋在纖維素表面形成物理屏障,且對纖維素酶產生無效吸附,使纖維素酶酶解效率降低[3-4]。因此,為提高纖維素酶水解效率,通常需要將木質纖維原料進行預處理,以破壞其致密的結構,脫除半纖維素和木質素,提高纖維素酶的可及性[5-6]。
Wen等[7]研究表明,采用5%(體積分數)的乙酸在170 ℃下處理楊木可以高效制備低聚木糖,但是預處理后楊木中殘留的木質素含量(質量分數)高達35.6%,當酶劑量為20 FPU/g干物質量(DM)時,預處理后楊木的纖維素水解得率僅22.5%。因此,想要提高乙酸預處理后楊木的纖維素酶解率,需進一步脫除殘留的木質素。堿性亞硫酸鹽預處理技術廣泛應用在制漿造紙工業中,其對木質素的脫除效率較高[8]。因此,采用乙酸預處理楊木制備低聚木糖后,再經亞硫酸鹽預處理移除木質素,可提高楊木纖維素的轉化率。經過兩步預處理后楊木樣品中剩余木質素的結構和分布發生變化,這可能會引起木質素對纖維素酶的吸附及抑制特性發生改變。
筆者在前期的研究中已證實乙酸預處理會增強木質素對纖維素酶水解的抑制作用[9],而亞硫酸鹽二次預處理后木質素對纖維素酶水解的影響有待進一步探究。為探明二次預處理對楊木的酶解效率以及對楊木木質素非生產性吸附纖維素酶的影響,分離了乙酸-亞硫酸鹽兩步預處理前后的楊木木質素,并運用凝膠滲透色譜(GPC)、二維異核單量子關系核磁(HSQC-NMR)、定量磷譜(31P NMR)等技術分析了木質素在兩步預處理過程中的結構和理化性質的變化,考察了預處理后楊木纖維素酶解率以及兩步預處理對木質素吸附/脫附纖維素酶的影響。
1 材料與方法
1.1 試驗材料
楊木(PopulusL.)屑產自江蘇宿遷。首先將楊木屑進行粉碎、過60目篩(孔徑為0.3 mm)制備楊木粉備用。用有機溶劑(甲苯和乙醇體積比為2∶1)對楊木粉脫蠟6 h,將剩余固體在45 ℃下通風干燥24 h。經測定楊木的化學組成(質量分數)為:46.33%葡聚糖、18.25%木聚糖和29.83%木質素(其中包含25.12%的酸不溶木質素和4.71%的酸溶木質素)。參照文獻[10]報道的球磨條件,對上述脫蠟后的楊木粉進行球磨,制得球磨后楊木樣品(BM-poplar)。
商品纖維素酶Cellic Ctec2,購自丹麥Novozyme公司,其酶活為137.8 FPU/mL;微晶纖維素Avicel PH-101、木聚糖酶(來自Aspergillusoryzae)、蛋白酶(來自Streptomycesgriseus)、葡萄糖、氯仿-d1、二甲基亞砜-d6、2-氯-4,4,5,5-四甲基-1,3,2-二氧磷雜環戊烷、環己醇、乙酰丙酮鉻(Ⅲ)、吡啶、乙酸酐以及乙酸,購自美國Sigma-Aldrich公司;蛋白檢測試劑盒(二喹啉甲酸,BCA法)購自上海碧云天生物技術公司。
1.2 楊木預處理
將BM-poplar和5%(體積分數)乙酸按照固液比1∶10(g∶mL)在170 ℃反應30 min,反應結束后,將反應釜迅速冷卻至20 ℃。采用真空抽濾收集反應剩余固體,并用蒸餾水將其洗至中性,最后將固體部分凍干即得到乙酸預處理后的楊木樣品(AA-poplar)。
參照文獻[8]報道的條件,采用堿性亞硫酸鹽對AA-poplar進行第2步預處理。具體操作如下:將AA-poplar與含有1%(質量分數)的氫氧化鈉和4%(質量分數)的亞硫酸鈉的水溶液按照固液比1∶10(g∶mL)在121 ℃下反應60 min。反應結束后,用真空抽濾收集剩余固體,并用蒸餾水將其洗至中性,最后將固體剩余物凍干即得到乙酸-亞硫酸鹽兩步預處理后的楊木樣品(AA-AS-poplar)。參照文獻[11]分析預處理后楊木樣品的化學組分,結果為2次重復測定的平均值。
1.3 楊木的X射線光電子能譜分析
采用Escalab 250Xi型X射線光電子能譜儀(XPS,美國Thermo Fisher Scientific)對楊木樣品表層(2~10 nm)碳原子的化學環境進行檢測,且樣品表面木質素的覆蓋率S木質素按下式計算[12]:
S木質素=[n(O/C)-0.83]/(0.33-0.83)×100%
(1)
式中:n(O/C)為XPS測得的楊木樣品表面的氧碳比;0.83為n(O/C碳水化合物)值;0.33為n(O/C木質素)值。
1.4 木質素的制備
用纖維素酶(60 FPU/g DM)和木聚糖酶(299.4 U/g DM)在50 ℃下水解楊木樣品(BM-poplar和AA-AS-poplar)72 h,重復3次,水解結束后,5 000 r/min下離心5 min得到固體殘渣。將上述水解殘渣均勻分散在pH 7.0的磷酸緩沖液中配成固含量為5%(質量分數)的物料體系,添加蛋白酶(20 U/g DM)在37 ℃水解24 h。水解結束后,用蒸餾水對水解殘渣進行充分洗滌,然后5 000 r/min下離心5 min得到固體殘渣,凍干后即得到BM-poplar和AA-AS-poplar的酶解殘渣。經測定,AA-AS-poplar的酶解殘渣中的木質素含量大于93%,無須進一步提取純化,直接將其作為乙酸-亞硫酸鹽預處理后的木質素樣品(AA-AS-lignin)進行后續實驗。參照文獻[13]報道的提取方法,采用二氧六環(二氧六環和水體積比為96∶4)從BM-poplar的酶解殘渣中提取分離得到木質素樣品(BM-lignin)。
1.5 纖維素酶水解實驗
在預處理后的2%(質量分數)楊木樣品中分別添加5,10和20 FPU/g DM的纖維素酶,并在pH 5.0、50 ℃的條件下水解48 h。待水解完成后,用高效液相色譜(HPLC)測定水解上清液中的葡萄糖含量。實驗結果為2組平行實驗的平均值。
為考察預處理后楊木木質素對纖維素酶水解的影響,將BM-lignin和AA-AS-lignin分別添加到2%(質量分數)Avicel的酶解體系中,在50 ℃下水解6~48 h,其中木質素的添加量為200 mg/g纖維素,纖維素酶劑量為10 FPU/g纖維素。待水解完成后,于5 000 r/min下離心10 min取上清液,采用HPLC測定上清液中的葡萄糖含量。實驗結果為2組平行實驗的平均值,且標準誤差以誤差線顯示。
1.6 纖維素酶吸附實驗
將不同含量的纖維素酶(24~480 mg/g 木質素)與1%(質量分數)木質素在pH 5.0、4 ℃、磁力攪拌的條件下平衡1 h。然后,將固液兩相離心分離(4 ℃、5 000 r/min、10 min),上清液中游離蛋白含量的測定方法參照文獻[9]。木質素吸附的酶蛋白量為初始添加的酶蛋白量與上清液中游離蛋白量的差值。根據Langmuir吸附等溫線方程可計算得到木質素對纖維素酶的吸附參數[13]。實驗結果為2組平行測定的平均值,標準誤差以誤差線顯示。
1.7 纖維素酶脫附實驗
將1%(質量分數)木質素與纖維素酶(480 mg/g木質素)在4 ℃下進行吸附-脫附實驗,具體操作流程及纖維素酶脫附回收率計算公式見文獻[14]。采用蛋白質量檢測BCA法測定所得到的吸附和脫附上清液中的蛋白含量。將最終所得的固體殘渣(木質素-纖維素酶復合物)分成2等份,其中一份作為纖維素酶源于50 ℃水解1%(質量分數)Avicel 48 h,待水解結束后,用HPLC測定上清液中的葡萄糖含量;另一半用于氮元素分析。采用有機元素分析儀(德國Elementar Analysen systeme GmbH)對木質素-纖維素酶復合物中的氮元素含量進行測定,測試結果為2組重復測定的平均值。
1.8 木質素的表征
將木質素粉末與乙酸酐/吡啶混合液(體積比1∶1)按照固液比25∶1(mg∶mL)混合,在室溫、磁力攪拌的條件下反應24 h將木質素乙酰化[15],然后將乙酰化的木質素溶解到四氫呋喃溶劑中,采用GPC測定木質素的分子量。流動相為四氫呋喃,流速為1.0 mL/min,所用的凝膠滲透色譜儀型號為Waters1525(美國Waters),檢測器為Waters 2414示差折光檢測器,色譜柱型號為Agilent PL gel 5 μm MIXED-C(美國Agilent)。
取50 mg木質素溶解于1 mL DMSO-d6,將溶解好的樣品轉移至直徑為5 mm的核磁管中待測。采用500 MHz的AVIII核磁波譜儀(瑞士Bruker)對木質素進行HSQC-NMR分析。參數設置為:標準脈沖序列“hsqcetgpsi”;1H維度的采樣點數為1 024,13C維度的采樣點數為256;弛豫時間為1.5 s(D1),累加(NS)64次。以環己醇作為內標,采用定量31P NMR對木質素的各類羥基含量進行測定[16]。測試條件:脈沖延遲時間25 s,累積次數192次。采用TopSpin 3.5軟件對核磁譜圖進行定量積分計算。
采用有機元素分析儀測定木質素樣品的硫元素含量,測試結果為3次重復測定的平均值。木質素的Zeta電位和疏水性的測定步驟見文獻[14]。
2 結果與分析
2.1 預處理后楊木的化學組分及酶水解
楊木經乙酸預處理可以高效制備低聚木糖[7]。預處理后楊木的化學組分見表1。乙酸預處理對木質素的脫除效果較差。為進一步有效脫除乙酸預處理后楊木中的剩余木質素,對其進行了亞硫酸鹽第2步預處理。結果顯示,亞硫酸鹽預處理后殘渣中木質素的含量(質量分數,下同)從34.23%減少到26.75%,這可能是因為木質素以木質素磺酸鹽的形式溶出[17]。經過乙酸-亞硫酸鹽兩步預處理后,楊木木聚糖的含量從18.25%降低到0.65%。這說明在乙酸-亞硫酸鹽兩步預處理過程中楊木半纖維素大量溶出。隨著木質素和木聚糖含量的減少,葡聚糖的相對含量從46.33%增加到72.60%。
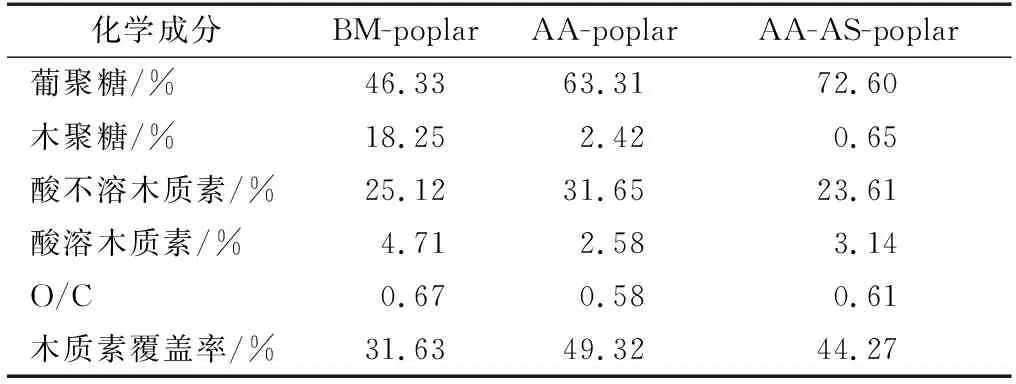
表1 預處理后楊木的化學組分Table 1 Chemical compositions of poplar after pretreatment

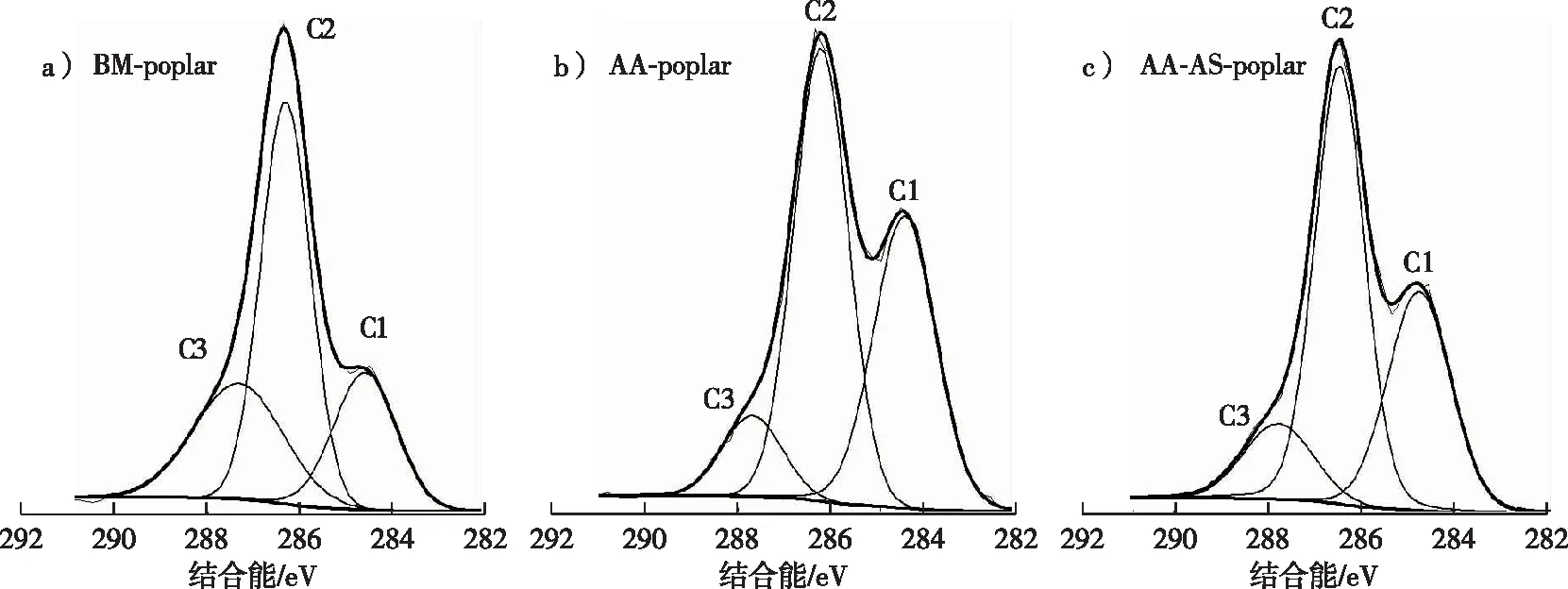
圖1 預處理后楊木的XPS譜圖Fig. 1 XPS spectras of poplar samples after pretreatment
乙酸-亞硫酸鹽兩步預處理后楊木水解的葡萄糖得率有明顯的提高,當酶劑量為20 FPU/g DM時,葡萄糖得率可達到67.1%,而在同等條件下BM-poplar水解的葡萄糖得率僅為32.4%。這歸因于第2步預處理后楊木中的木聚糖和木質素得到了進一步的脫除,增加了纖維素的可及性。盡管兩步預處理對木質素的脫除效果有限,而且AA-AS-poplar樣品表面的木質素覆蓋率高達44.27%(表1),但是兩步預處理后楊木的水解得率仍然有明顯提高,這說明預處理后木質素在原料中的分布以及結構變化均會對原料的纖維素酶水解產生影響。然而,在酶增加到20 FPU/g DM時AA-AS-poplar的水解得率仍未超過70%。因此,有必要對乙酸-亞硫酸鹽兩步預處理后楊木木質素的化學結構及理化性質進行表征,進而探明其對纖維素酶水解的影響。
2.2 木質素的表征
木質素樣品的化學組分分析表明,兩種木質素樣品的純度較高,木質素含量均大于93%。其中,BM-lignin含有少量的葡聚糖(0.7%)和木聚糖(1.8%),可能是因為木質素-碳水化合物連接鍵在球磨及酶水解過程中不易斷裂[5]。AA-AS-lignin是未經有機溶劑萃取的酶解殘渣,因而有少量的葡聚糖(1.3%)以及木聚糖(0.6%)殘留。木質素分子量測定結果顯示,乙酸-亞硫酸鹽兩步處理后木質素的重均分子量(Mw)和數均分子量(Mn)分別從12 211和6 372 g/mol(BM-lignin)降低到了1 206 和844 g/mol,推測是因為在預處理過程中木質素的醚鍵大量斷裂[19],而且木質素以木質素磺酸鹽的形式溶出[17]。此外,乙酸-亞硫酸鹽兩步預處理后木質素分子量分散系數(PDI)從1.91減小至1.43,說明預處理使木質素的分子量大小變得更加均一。
采用HSQC-NMR對木質素樣品的結構單元及連接鍵進行定量分析(圖2)。木質素的主要結構單元和連接鍵分別在HSQC譜圖的芳環區(δ100.0~140.0/6.0~8.0)和側鏈區(δ50.0~90.0/2.5~6.0)進行定量計算[15],計算結果列于表2中。
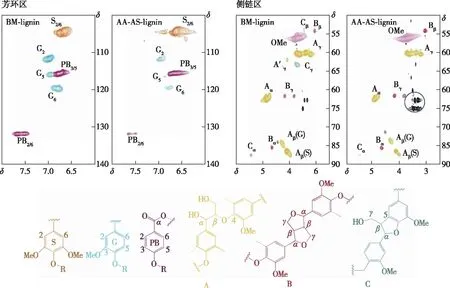
圖2 木質素的二維HSQC核磁譜圖Fig. 2 HSQC-NMR spectras of lignin

表2 采用二維HSQC核磁分析得到的木質素子結構的相對含量Table 2 Quantitative information of lignin substructures by HSQC-NMR analysis
在HSQC譜圖的芳環區,木質素的紫丁香基(S)、愈創木基(G)和對羥苯基(PB)單元的相對含量分別由S單元的C2,6—H2,6相關信號、G單元的C2—H2相關信號以及PB單元的C2,6—H2,6相關信號進行積分計算。其中,各結構單元的含量是被測結構單元占S和G單元總數的百分數。由表2可知,乙酸-亞硫酸鹽兩步預處理后木質素的S單元增加,而G單元的含量減少,因而S/G值從1.4(BM-lignin)增加到4.0(AA-AS-lignin),而PB單元的含量隨著S/G的增加而減少。在木質素的側鏈區,采用β-O-4醚鍵(A)、樹脂醇結構(β-β,B)以及苯基香豆滿結構(β-5,C)的α位碳氫相關信號(Cα-Hα)進行積分計算各連接鍵的相對含量。其中,各連接鍵的含量是指被測連接鍵占β-O-4、β-β和β-5總數的比例。乙酸-亞硫酸鹽兩步預處理后木質素的β-O-4醚鍵減少,而β-β和β-5的相對含量增加。這是因為在酸性或者堿性環境的高溫預處理過程中,木質素醚鍵的斷裂會伴隨著木質素碎片的再縮合反應發生[19]。結合木質素的分子量數據,推測在兩步預處理過程中木質素的解聚和縮合同時發生,而解聚反應占據主導地位,最終導致木質素的分子量減小。另外,在AA-AS-lignin的側鏈區可以看到木聚糖及葡聚糖的相關信號(圖2),位于δ72.8~75.2/3.2~3.5。這是由于AA-AS-lignin是未經有機溶劑萃取提純的酶解殘渣,其中的碳水化合物比如葡聚糖含量相對較高。
以環己醇為內標,對磷化后的木質素樣品進行定量磷譜分析(圖3),譜圖的歸屬情況見前期報道[9]。結果顯示,乙酸-亞硫酸鹽兩步預處理后木質素的脂肪族羥基含量減少,而酚羥基(S—OH、G—OH及PB—OH)的含量明顯增加(表3),這可能是由于木質素β-O-4醚鍵斷裂后形成了新的酚羥基[13]。乙酸-亞硫酸鹽兩步預處理后隨著木質素S—OH和G—OH的含量增加,PB—OH的含量則從0.45 mmol/g降低到0.08 mmol/g,這與表2中木質素PB單元的相對含量的變化趨勢一致。此外,AA-AS-lignin的譜圖中可以看到一些雜峰位于δ144~145,這可能是木質素樣品中殘留多糖的羥基峰[16]。
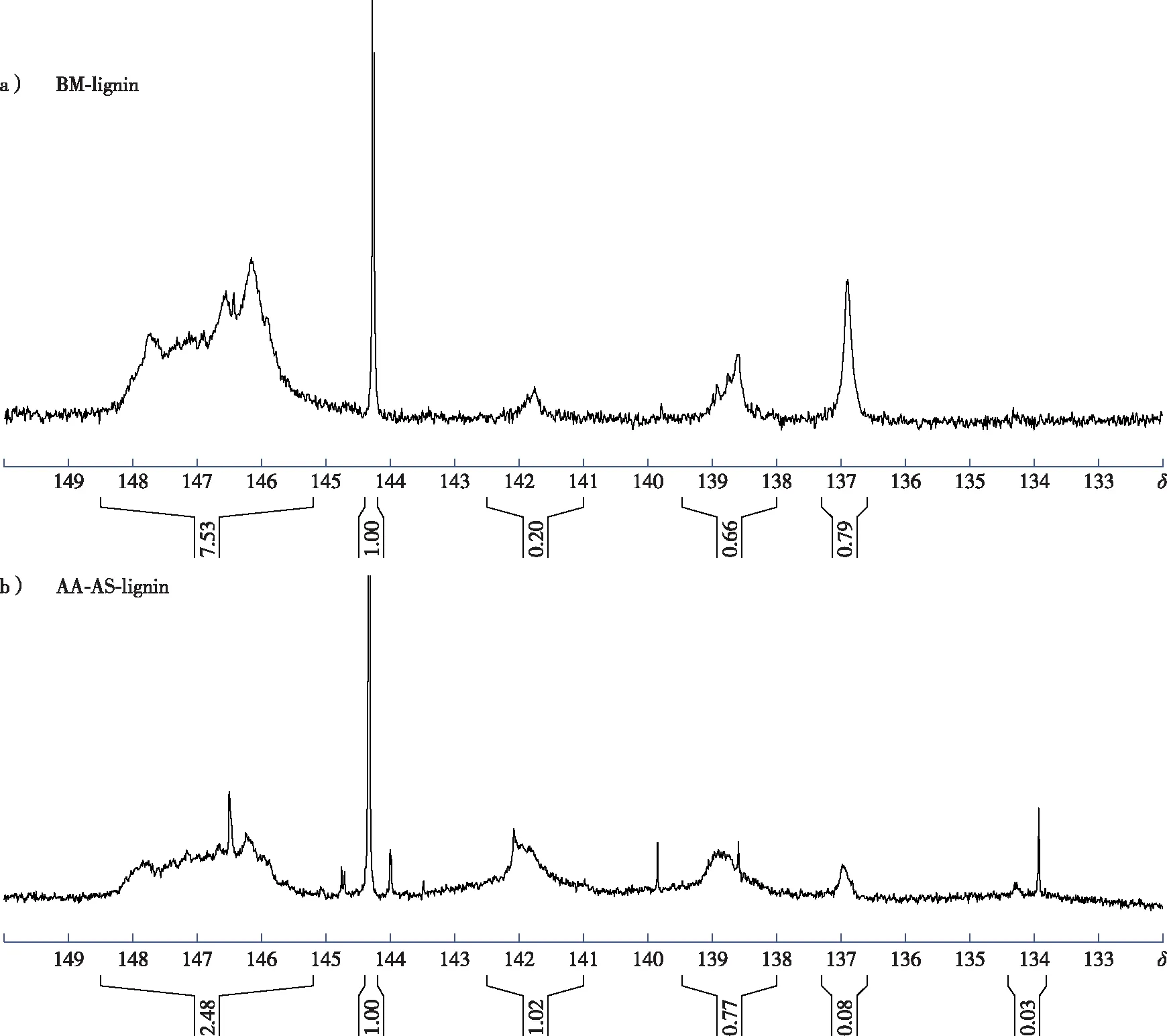
圖3 木質素的定量磷譜圖Fig. 3 Quantitative 31P NMR spectra of lignin
木質素的靜電特性和疏水性會影響其對纖維素酶的吸附能力[20]。兩個木質素在水相環境下均帶負電荷,且兩步預處理后木質素的Zeta電位從-47.63 mV(BM-lignin)略降低到-44.23 mV(AA-AS-lignin)。據報道,木質素羥基和羧基可以在水相環境下解離而帶負電荷[21]。與BM-lignin(0.05%)相比,AS-AA-lignin的硫元素含量增加到0.76%,說明亞硫酸鹽預處理后木質素中發生了磺化。因而AS-AA-lignin的負電荷可能來自于磺酸基、羥基和羧基的解離。木質素基團的解離能力與其所處化學環境有關[21],故而乙酸-亞硫酸鹽兩步預處理后木質素Zeta電位略降低的原因是很可能與其脂肪族羥基、酚羥基、羧基和磺酸基含量變化有關(表3)。未預處理的BM-lignin的疏水性較高為3.7 L/g,而經過兩步預處理后木質素的疏水性降低至0.89 L/g。木質素的疏水性基團(如甲氧基和芳基等)和親水性基團(如羥基和羧基等)的比例決定其總的疏水性大小。因此,預處理后木質素官能團的形成與消去會影響其整體疏水性的大小。BM-lignin較高的疏水性表明楊木的原本木質素含有較多的疏水性基團。AA-AS-lignin疏水性的降低可能是由于木質素側鏈斷裂之后形成了大量的親水性官能團,如羥基和羧基(表3)。此外,木質素的磺化也會降低其疏水性[8,22]。

表3 采用定量磷譜對木質素各羥基含量的測定結果Table 3 Quantitative information on OH groups in lignin by a quantitative 31P NMR method mmol/g
2.3 木質素對纖維素酶水解的影響
一般認為,從球磨后的木質纖維原料中分離得到的木質素樣品保留了原料中木質素的結構特征[13]。結果表明,添加BM-lignin后Avicel水解的葡萄糖得率幾乎沒有降低,而添加AA-AS-lignin后對Avicel酶水解的抑制率達到了16.5%(圖4)。Kellock等[23]的研究表明纖維素酶對原本木質素的吸附能力較弱,這可以解釋BM-lignin對纖維素酶水解較弱的抑制作用。然而,乙酸-亞硫酸鹽兩步預處理后木質素對纖維素酶水解的抑制作用明顯增強。據報道,木質素磺酸鹽可以表現出表面活性劑的特性,通過靜電斥力減少木質素對纖維素酶的非生產性吸附,進而促進纖維素酶水解[17]。盡管AA-AS-lignin也被磺化,但是AA-AS-lignin卻表現出較強的抑制作用,導致二者對纖維素酶水解產生不同影響的原因很可能是其結構差異較大。木質素磺酸鹽是亞硫酸鹽預處理水解液中的游離態木質素衍生物,而AA-AS-lignin是經過乙酸-亞硫酸鹽兩步預處理后的剩余木質素固體殘渣。兩步預處理過程中木質素分子側鏈的斷裂產生的大量酚羥基很可能是導致其抑制作用增強的原因之一[13]。此外,AA-AS-lignin較高的S/G比值也可能是導致其對纖維素酶水解抑制增強的原因[24-25],究其原因可能是木質素S單元比G單元對纖維素酶水解有更顯著的抑制效果[24]。
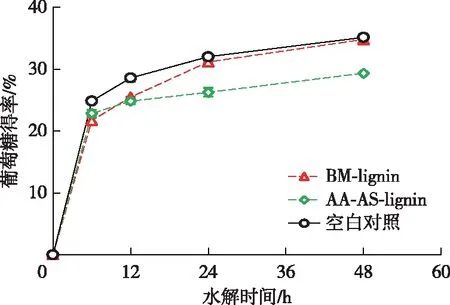
圖4 木質素對纖維素酶水解的影響Fig. 4 Effects of lignin on enzymatic hydrolysis of cellulose
2.4 木質素對纖維素酶的吸附
木質素對纖維素酶的吸附符合Langmuir吸附等溫線(決定系數R2>0.8)(圖5)。BM-lignin的最大吸附容量(Pads,m)為55.7 mg/g,而AA-AS-lignin的Pads,m則增大到了175.0 mg/g,表明兩步預處理后木質素對纖維素酶的吸附位點增多。結合強度是由Pads,m乘以吸附平衡常數(Kp,BM-lignin為0.44,AA-AS-lignin為0.41)所得,它可以更加全面地反映木質素對纖維素酶的吸附強度。兩步預處理后木質素的結合強度從24.7 mL/g(BM-lignin)增大到72.1 mL/g。木質素的疏水性被認為是其吸附纖維素酶的主要推動力[20],而帶負電荷較多的木質素與纖維素酶之間存在較強的靜電斥力[17,20]。
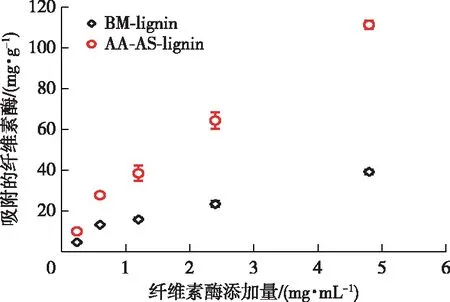
圖5 木質素對纖維素酶的吸附等溫線Fig. 5 Adsorption isotherm of cellulase on lignin
實驗結果顯示,雖然AA-AS-lignin有較低的疏水性,但是卻表現出較大的吸附纖維素酶的能力。另外,AA-AS-lignin和BM-lignin的Zeta電位很接近,但是二者卻表現出不同的纖維素酶吸附能力,說明乙酸-亞硫酸鹽預處理后木質素的疏水性和Zeta電位不是影響其對纖維素酶吸附能力的主導因素。據報道,木質素發生磺化后所帶的磺酸基與帶負電的纖維素酶之間可產生較強的靜電斥力,進而緩解其對纖維素酶的吸附[8],然而,被磺化的AA-AS-lignin卻表現出較大的纖維素酶吸附能力。結合木質素羥基官能團的分析結果(表3),推測導致AA-AS-lignin對纖維素酶吸附能力增強的原因很可能是兩步預處理后木質素的酚羥基含量的增多(表2),因為木質素酚羥基的增加會使其對纖維素酶的吸附能力增強[13,22]。
2.5 木質素對纖維素酶的脫附
木質素脫附纖維素酶的定量分析結果見表4。當20.0 mg的木質素與9.6 mg的纖維素酶在4 ℃下平衡60 min后,約有0.39和1.11 mg的纖維素酶分別被BM-lignin和AA-AS-lignin吸附。采用新鮮的緩沖液對木質素吸附的纖維素酶進行洗脫,纖維素酶的回收率分別達到61.1%和28.8%,說明兩步預處理后木質素對纖維素酶的脫附能力降低。吸附-脫附之后所得的木質素-纖維素酶復合物中氮元素含量的高低可以間接地反映木質素表面吸附的酶量多少[23]。由表4可知,AA-AS-lignin與纖維素酶復合物的氮元素含量較高,這進一步證實了兩步預處理后木質素對纖維素酶的脫附能力降低。以木質素-纖維素酶復合物作為酶源對Avicel進行水解,并測定葡萄糖得率。結果表明,盡管AA-AS-lignin結合了較多的纖維素酶,但是采用AA-AS-lignin-纖維素酶復合物水解Avicel卻獲得較低的葡萄糖得率(3.0%)(表4)。木質素-纖維素酶復合物的酶活損失可能是歸因于木質素與纖維素酶之間較強的結合力,這會導致纖維素酶與纖維素有效接觸的機會減少[23]。因而AA-AS-lignin與纖維素酶較強的結合強度可能是導致AA-AS-lignin-纖維素酶復合物水解能力較低的原因。
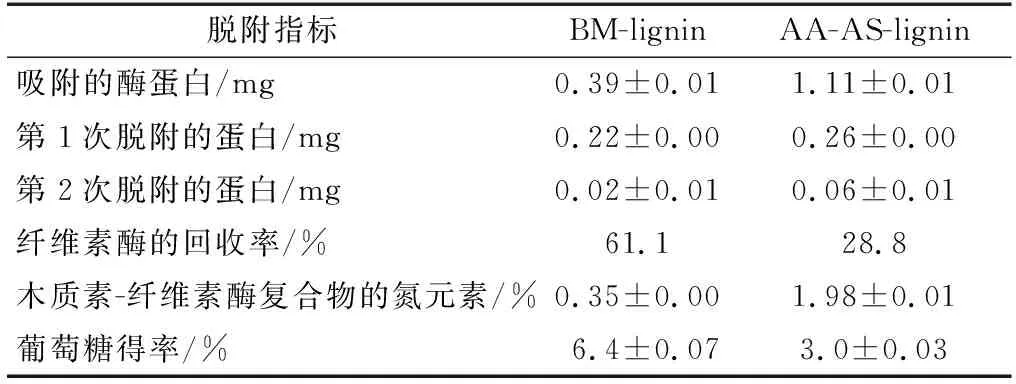
表4 木質素對纖維素酶的脫附Table 4 Desorption of cellulase from lignin
3 結 論
采用亞硫酸鹽對乙酸預處理后的楊木殘渣進行二次預處理,重點探究了乙酸-亞硫酸鹽兩步預處理對楊木化學組分、酶水解率、殘渣中木質素的結構及其對纖維素酶的吸附和抑制的影響。主要的結論如下:
1)乙酸-亞硫酸鹽兩步預處理后楊木的木質素含量減少,預處理后楊木的水解得率從32.4%提高到67.1%。兩步預處理使木質素嚴重降解,預處理后木質素發生磺化并且其S/G比值及酚羥基含量增加。兩步預處理后木質素對纖維素酶的結合強度從24.7 mL/g增大到72.1 mL/g,而其對纖維素酶的脫附能力降低,預處理后木質素對纖維素酶水解的抑制率從1.0%增至16.5%。
2)乙酸-亞硫酸鹽兩步預處理后楊木中剩余木質素對纖維素酶的非生產性吸附及空間阻礙都對后續的纖維素酶水解不利。相比草本原料,闊葉材楊木的纖維結構更為致密堅硬,致使其木質素更難脫除。另外,在170 ℃的高溫乙酸預處理過程中會形成大量的木質素縮合結構。以上都可能是導致乙酸-亞硫酸鹽預處理后楊木木質素脫除率低,進而使預處理后楊木纖維素水解率低的主要原因。因而,為實現楊木的多組分高效轉化利用,第2步亞硫酸鹽預處理的條件(如乙酸及亞硫酸鹽兩步預處理的溫度和試劑濃度等)需要繼續優化。
