我國巴西橡膠樹炭疽病的研究進展
2021-10-14 04:30:00林春花劉文波繆衛國
熱帶生物學報 2021年3期
林春花,張 宇,劉文波,李 瀟,繆衛國
(海南大學 植物保護學院/熱帶農林生物災害綠色防控教育部重點實驗室,海口 570228)
炭疽菌屬(ColletotrichumCorda),又稱為刺盤孢屬,是子囊菌門糞殼菌綱肉座菌亞綱小叢殼科小叢殼屬(Glomerella)的無性態。炭疽菌屬真菌廣泛分布于熱帶、亞熱帶和溫帶地區,其寄主范圍十分廣泛,包括蔬菜、牧草、果樹、花卉等重要經濟作物,是一類重要的植物病原菌[1]。炭疽菌危害各種農林作物,能夠引起寄主植物葉斑、枝枯、果實腐爛、死苗等炭疽病癥狀,造成重大的經濟損失。基于其經濟意義和科學研究意義,炭疽菌已經被列為世界八大重要病原真菌之一[2]。天然橡膠是重要的工業原料和戰略物資,含有天然橡膠的植物有2 000多種,巴西橡膠樹(Hevea brasiliensisMuell. Arg.,簡稱橡膠樹)是大面積商業化種植生產天然橡膠的植物,其產量占目前世界天然橡膠總產量的99%以上[3]。全世界有40多個國家和地區種植巴西橡膠樹[3]。截至2018年,亞洲橡膠產量占91.13%、非洲占6.49%、美洲占2.38%[4]。其中亞洲以印度尼西亞植膠面積居第一,泰國第二,馬來西亞和中國面積相近,并列第三,越南和印度位居第五和第六,這6國植膠面積約占世界種植總面積的72%[3]。我國橡膠樹種植過程中受到各種病蟲害危害,橡膠樹炭疽病是其中危害最普遍且嚴重的葉部“兩病”之一[5]。該病害在國內外流行范圍逐年擴大,發病趨勢越來越嚴重,造成橡膠產量損失不斷增加,一直以來都受到科技工作者的重視。近年來,我國橡膠樹炭疽菌取得了較好的研究進展,尤其是在病原種類鑒定及田間優勢種的確定、病原菌致病機制研究等方面。筆者對近年來橡膠樹炭疽病發展趨勢、癥狀類型、病原種類、病原菌生物學特性、分子機制和防治措施等研究現狀進行概述,旨在為深入研究橡膠樹炭疽病提供參考。
1 發生與危害
1905年,PETCH首次報道橡膠樹炭疽病在斯里蘭卡發生危害[6]。隨后在馬來西亞、印度、泰國和中國橡膠樹種植地都有報道,為橡膠樹種植區的常見病害[7?8]。在我國,最初記載橡膠樹炭疽病害只在苗圃和新植幼樹上有少量發現[9],屬次要病害。1962年在海南大豐農場橡膠樹炭疽病害引起個別品系開割樹落葉、不能割膠,此為我國首次記錄;1992年在海南暢好農場發生橡膠樹炭疽病的大面積流行,發病面積占開割林地面積75%,造成4~5級落葉的有20多萬株,導致割膠推遲,該農場當年損失干膠250 t[10]。由于大量更新和推廣高產品系,該病發生日趨嚴重,1996年僅海南墾區炭疽病發生面積達73萬hm2,損失干膠15 000 t[11]。2004年,在云南西雙版納、紅河、普洱、臨滄、文山等橡膠種植區不同程度發生橡膠樹老葉炭疽病和開割樹冬春新抽枝葉炭疽病,造成大量落葉和個別林段干膠減產,橡膠炭疽病已經從次要病害上升為重要病害[12]。
2012—2013年,李濤等在云南河口對橡膠樹炭疽病觀察發現,橡膠樹炭疽病一般在橡膠樹嫩葉期、高濕度的情況下發生危害,每年有2~3個發病高峰期,病情有逐年加重現象[13]。近年來,橡膠樹炭疽病和橡膠樹白粉病并稱為我國橡膠樹葉部重要“兩病”[14],是我國農業農村部指定的橡膠樹“兩病”春防工作的防治對象之一[15]。橡膠樹炭疽病從早期危害苗圃,到頻頻危害不同地區開割膠林,近年來被列為橡膠樹重要春防對象之一。橡膠樹炭疽病的發生有越來越嚴重的趨勢,已成為我國橡膠樹重要葉部病害之一。
2 危害癥狀
橡膠樹炭疽病可危害苗圃小苗、大田幼樹直至成齡開割膠樹,侵染嫩葉、葉柄、嫩梢和膠果等部位,引起嫩葉脫落、嫩梢回枯、果實腐爛,甚至形成僵果掛在樹上。嚴重時會引起膠樹的重復落葉和嫩梢回枯,推遲開割時間。
通常,橡膠樹炭疽病危害不同部位呈現的癥狀各異。炭疽病主要危害部位是葉部,嫩葉和老葉均可危害。古銅色嫩葉染病后,初期葉尖葉緣呈不規則暗綠色水漬狀病斑,隨后變黑壞死、扭曲,最終導致葉片脫落枝條干枯[10,12,16](圖1-A)。淡綠色嫩葉染病后,葉尖、葉緣呈現圓形或不規則形(圖1-B),暗綠色似開水燙過一樣的水漬狀病斑,病斑較大,有時在病斑邊緣可見黑色壞死錢[17]。在高濕條件下,常在病部長出的粉紅色粘稠孢子堆[10,12,17]。淡綠色后期或老化葉染病后,可呈現多種不同類型癥狀:1)圓形和半圓形型(圖1-C),初期呈現水漬狀小斑,隨后病斑擴大,病斑多在葉尖葉緣處呈半圓形,或在葉片上呈圓形[12,17?18]。2)不規則型(圖1-D),病斑初期呈現灰褐色或紅褐色近圓形,病健交界明顯,后期病斑相連成片,形狀不規則,有的有穿孔[10]。3)葉緣枯型(圖1-E),受害初期葉尖或葉緣退綠變黃,隨后病葉葉緣由外向內表現出先黃后枯,病斑呈現灰白色,病健交界呈鋸齒狀[10,12,18]。4)凸起圓錐型(圖1-F),病斑凸起成小圓錐體,病斑邊緣皺縮,嚴重時可看到整張葉片布滿向上凸起的小點,有穿孔[10,18]。炭疽病在老葉上容易產生小黑點(分生孢子器),小黑點有呈輪紋排列分布,也有分散分布(圖1-G)。嫩稍、葉柄、葉脈染病后,出現黑色下陷小點或者黑色條斑(圖1-H)。嫩稍染病后除了葉片掉落,還容易發生頂芽回枯(圖1-I)。綠果染病后,病斑暗綠色,水漬狀腐爛。在高濕條件下,發病部位長出粘稠狀的粉紅色孢子堆[17]。

圖1 橡膠樹炭疽病癥狀Fig. 1 The symptoms of rubber tree anthracnose
3 病原菌種類和田間優勢種
炭疽菌屬種類繁多,遺傳多態性豐富。由于該屬真菌分類鑒定依據在不斷變化,對橡膠樹炭疽病病原菌的種類鑒定及名稱也隨之發生變更(表1)。
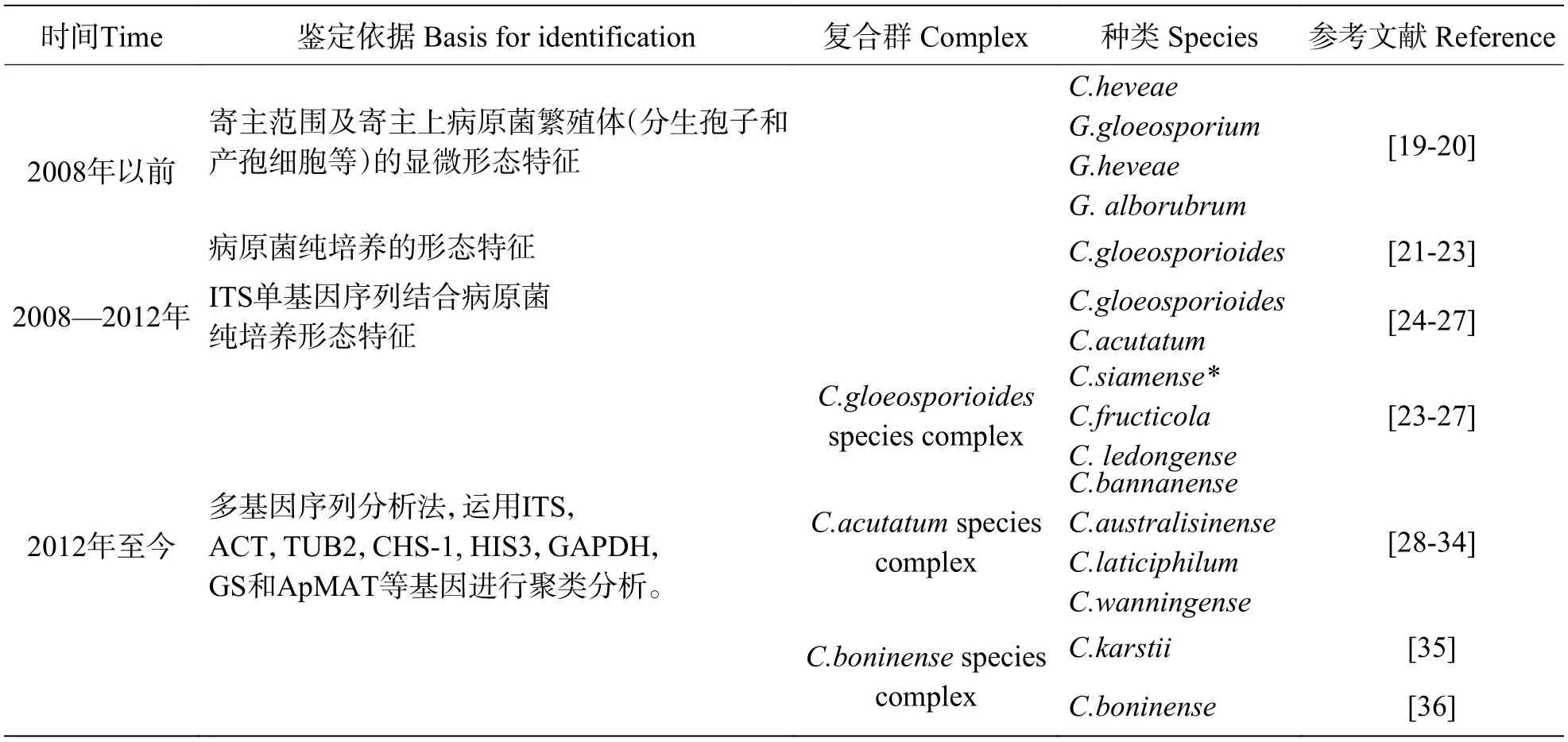
表1 我國橡膠樹炭疽病病原種類鑒定情況Tab. 1 Identification of pathogen of rubber tree anthracnose in China
早期炭疽菌依據寄主范圍或寄主上病原菌繁殖體顯微形態特征進行分類和命名。1905年斯里蘭卡PETCH首次報道橡膠樹炭疽病時,根據寄主范圍和寄主上的分生孢子和產孢細胞,將病原菌鑒定為C.heveaePetch,Gloeosporium heveaePetch和G. alborubrumPetch[19]。戴芳瀾先生在《中國真菌總匯》中將福建橡膠樹炭疽菌記載為C. heveaePetch,將云南橡膠樹的炭疽菌記載為G. alborubrum[20]。根據寄主范圍進行鑒定,易導致許多物種同物異名。隨后以炭疽菌純培養的形態特征(分生孢子、產孢細胞、附著胞等)和寄主范圍,將橡膠炭疽病病原菌無性世代鑒定為C. gloeosporioidesf.heveaePenz.,有性世代鑒定為Glomerella cingulata[21?23]。因此,2008年以前我國橡膠樹炭疽病病原菌主要鑒定為C. heveae或C.gloeosporioides。主要依據病原菌形態特征進行鑒定,容易導致許多相近種難以區分。
隨著分子生物學的發展,核糖體DNA基因轉錄隔離區(internal transcribed spacer, ITS)在真菌種類鑒定中得到廣泛應用,學者逐步采用形態鑒定法結合分子鑒定方法對橡膠樹炭疽菌進行鑒定。橡膠炭疽菌的鑒定逐漸采用形態特征結合ITS單基因序列分析進行。2008年張春霞等[24]通過觀察分生孢子形態,鑒定出引起云南西雙版納橡膠樹的炭疽菌除了膠孢炭疽菌,還有尖孢炭疽菌(C.acutatum)。隨后李繼鋒(2010)、劉先寶(2011)等[25?26]分別采用rDNA-ITS序列相繼驗證了在海南、廣東、廣西都可從橡膠樹上分離到尖孢炭疽菌。2014年,筆者基于ITS序列設計的特異性引物,對我國橡膠樹炭疽菌進行分子鑒定和分布分析,結果顯示,來自海南、云南和廣東廣西的138株炭疽菌中,72.46%鑒定為膠孢炭疽菌[27]。因此,2008—2012年間,我國橡膠樹炭疽菌鑒定有C.gloeosporioides和C.acutatum2種,其中田間優勢種為C.gloeosporioides[24?27]。
由于ITS序列分析在炭疽菌分類研究中仍存在一定局限性,對于多數近緣相似種,仍不能準確反映和有效識別其親緣關系。多基因系統分類方法在炭疽菌等真菌種類鑒定中得到廣泛應用和認可。當前,炭疽菌鑒定主要利用多基因序列拼接分析法結合形態觀察對其進行種類鑒定。學者們紛紛利用該方法對各種作物炭疽病病原菌種類進行鑒定。本課題組率先對來自海南的橡膠樹炭疽菌進行多基因序列拼接分析,顯示田間有膠胞炭疽菌復合群(C. gloeosporioidesspecies complex)和尖孢炭疽菌復合群(C.acutatumspecies complex),其中膠胞炭疽菌復合群下主要有C. siamense和C. fructicola2個種,以C.siamense為田間優勢種[28?29]。CAO X鑒定出尖孢炭疽菌復合群有C. wanningense[30?31]。LIU X B 分析了來自我國的62株炭疽菌,顯示膠孢炭疽菌復合群下有C. siamense(28株)、C. fructicola(10株)和C.ledongense(2株),尖孢炭疽菌復合群主要有C. bannanense(3株)和C. australisinense(19株)[32]。此外,還有學者鑒定出尖孢炭疽菌復合群下有C. laticiphilum[33]和另一新種存在[34]。除此兩大復合群外,橡膠樹炭疽菌還有博寧炭疽菌復合群的C. karstii[35]和C. boninense[36]危害。以上研究結果表明,我國橡膠樹炭疽菌主要有膠胞炭疽菌復合群、尖孢炭疽菌復合群和博寧炭疽復合群(C.boniensespecies complex),復合群下有多個種組成,膠孢炭疽菌復合群下有C. siamense、C. fructicola、C. ledongense,尖孢炭疽菌復合群下有C. bannanense,C. australisinense,C.laticiphilum和C. wanningense,博寧炭疽復合群下有C. karstii、C.boninense。其中,我國橡膠樹種植區以膠孢炭疽菌復合群的C. siamense為田間優勢種。
4 橡膠樹炭疽菌生物學特性
對橡膠樹炭疽菌生物學特性的研究,包括病原菌菌落形態和生長速率、分生孢子形態大小、病原菌培養最佳條件、致病能力及對殺菌劑的敏感性等方面[11,23?24,37?40]。普遍認為橡膠樹炭疽菌存在豐富的生物學特性多樣,主要表現在菌落生長速率、分生孢子形態、營養利用、對藥劑的敏感性以及致病性等方面。菌落生長速率和營養利用方面的研究結果表明,在適生范圍內C.gloeosporioides的菌落直徑普遍大于C.acutatum,C.gloeosporioides在全光照條件下菌落直徑最大,C.acutatum在光照交替的環境性菌落直徑略大些,C.gloeosporioides在麥芽糖上生長最好,C.acutatum在甘露醇和乳糖上生長較好[11,24,38?39]。藥劑敏感性方面,C.gloeosporioides和C.acutatum對甲基托布津的敏感性具有差異顯著,C.acutatum對苯并咪唑類農藥有較強的抗藥性[10]。筆者監測發現橡膠樹炭疽菌對咪鮮胺具有較高的敏感性,不同地域來源的菌株藥劑敏感性有一定差異,例如來自海南的C.acutatum對咪鮮胺敏感性顯著高于來自云南的菌株[41];致病性方面,尖孢炭疽菌和C.gloeosporioides也具有致病力差異,在同類群炭疽菌中,不同菌株間也存在差異[10]。
筆者還比較分析了橡膠樹C.gloeosporioides和C.acutatum在侵染結構發育分化過程差異。結果表明,橡膠樹兩類群炭疽菌不同菌株間分生孢子萌發時間、孢子萌發率、附著胞形成時間和形成率有一定差異,但種間無明顯差異;研究還發現炭疽菌菌絲可誘發形成大量附著枝。分生孢子極易產生,可在菌絲頂端成簇或菌絲側面排列產生,也可由分生孢子形成的芽管產生,或在芽管分化附著胞過程分枝形成分生孢子;附著胞多著生于芽管頂端,少數附著胞頂端可繼續萌發類似短芽管結構,再次分化形成可黑色化的次級附著胞。以上結果表明,橡膠樹炭疽菌具有極強的適生性,這在一定程度解釋了炭疽菌株田間分布廣、危害面積大及炭疽病病情逐年嚴重的原因[42]。
近期,江濤等[34,43]測定了尖孢炭疽菌復合群下4個種(C. bannanense、C.laticiphilum、C.australisinense和歸為新類群的YHLC510)的生物學特性,結果說明,不同種的菌株對不同碳源、氮源利用,在不同溫度、pH、水活度下的生長情況等存在顯著差異;各菌株致病譜也存在明顯差異,C. bannanenseYNML52菌株具有強寄主分化型,其寄主范圍較窄[43]。施玉萍等[44]認為C. laticiphilum最適合生長溫度為26 ℃,最適合生長pH值為6,菌絲致死溫度為50 ℃、10 min,分生孢子致死溫度為51 ℃、10 min,能利用多種碳氮源,咪鮮胺和丙環唑對其具有很強毒力。
鑒于炭疽菌分類鑒定的依據發生多次變化,導致不同時期生物學特性測定結果不具有很好的可比性。早期認為的C.gloeosporioides和C.acutatum,現歸入復合群下的不同種。因此,前期測定的“同種”炭疽菌生物學特性結果可能是多個種的結果。因而早期認為炭疽菌存在生物學多樣性,可部分歸因于病原菌種類分類不一致。此外,對橡膠樹炭疽菌生物學測定研究中,通常以1個菌株或者少數幾個菌株代表一類群,供試菌株數量有限,在一定程度限制了結果的代表性。關于橡膠樹炭疽菌的生物學特性,仍需結合炭疽菌新的鑒定方法,補充完善不同種間的生物學特性異同,為防治策略制定奠定基礎。
5 橡膠樹病原菌致病分子機制研究現狀
炭疽菌屬屬于半活體營養寄生菌,在侵染寄主植物前期,并不立即殺死植物,營活體營養寄生;在入侵后期,隨著侵染菌絲的蔓延,殺死植物,營死體營養寄生[45]。關于炭疽菌的致病分子機制在C.gloeosporioides侵染錦葵科Malvaceae雜草[46]、C.dematium侵染豇豆[47]、C.higginsianum侵染擬南芥[48]、C.orbiculare侵染瓜類作物[49?50]、C.graminicola侵染玉米[51]等互作體系得到了廣泛的研究,內容涉及炭疽菌分生孢子粘附在寄主表面所需的黏蛋白及糖蛋白、分生孢子在寄主表面定殖和萌發所需的調控蛋白、穿透寄主的關鍵結構附著胞形成所需的黑色素合成相關酶、侵入寄主繼而產生侵入菌絲相關功能蛋白、抵御寄主防衛反應相關功能蛋白等[45]。
我國橡膠樹炭疽菌致病分子機理研究起步晚,始于近十年,研究內容主要集中于致病相關基因的鑒定和功能分析。初期,中國熱帶農業科學院環境與植物保護研究所利用根癌農桿菌介導T-DNA轉化法(Agrobacterium tumefaciens-mediated Transformation, ATMT),主要采用構建突變體庫的方法篩選致病力喪失或者減弱的相關突變體,進而克隆和研究致病相關基因功能[52?63]。近年來海南大學在與橡膠炭疽菌病菌生長發育和致病力相關的分泌蛋白、效應蛋白、G蛋白信號調控因子和轉錄因子等方面也取得了較好的研究進展[64?79]。
橡膠樹炭疽菌已經鑒定到的致病相關基因,主要影響菌株生長發育、孢子形成和萌發、附著胞形成和穿透寄主能力等方面。自噬相關蛋白CgATG4和CgATG8與菌絲生長、孢子萌發和致病力相關[62];效應蛋白CgSE1和CgSE2與膠胞炭疽菌的菌絲生長、分生孢子產量、穿透寄主能力相關[63];分泌蛋白BAS2(a biotrophy-associated secreted protein)也在一定程度影響菌絲生長、分生孢子的產量和致病能力[65];膠胞炭疽菌中一類二聚體蛋白(Dicer-like proteins)也參與調控病原菌菌絲生長、分生孢子形成和致病力[66];富集半胱氨酸的分泌蛋白CgCP1不影響菌絲的生長,但與分生孢子的產生有密切關系,還會誘發寄主產生活性氧和造成過敏性壞死,影響病原菌致病能力[67];G蛋白信號調控因子CgRGS1、CgRGS2和CgRGS7參與調控膠胞炭疽菌分生孢子產量及萌發[68?69],多個P型ATP酶通過影響膠孢炭疽菌穿透寄主表皮能力來影響致病能力[52,60?60]。Cg4LysM參與膠孢炭疽菌的物質能量代謝及生長發育過程,與致病性相關[70]。轉錄因子CgAP1影響菌體對滲透脅迫反應、細胞壁緊密度和致病能力[71];筆者所在團隊證實了膠胞炭疽菌脂滴包被蛋白CgCap20通過調控附著胞細胞壁孔徑大小,影響附著胞膨壓進而影響致病力[60?61];證實了脂滴包被蛋白CgCap20受到PKAC1的調控,與疏水蛋白互作[72?73];分析了解到炭疽菌HOG MAPK信號途徑下游轉錄因子CsATF1不僅影響滲透脅迫反應能力、吡咯類藥劑的敏感性,還參與致病功能[76?79]。
近期,劉先寶等[80]利用比較基因組學的方法,比較分析了能引起橡膠樹炭疽病兩種不同癥狀類型的C. siamense(菌株HBCG01)和C. australisinense(菌株GX1655),揭示了C. siamense比C. australisinense具有更豐富的分泌性碳水化合物活性酶(CAZymes), 細胞色素氧化酶(CYP450)和次生代謝產物(secondary metabolites),而C. australisinense中具有植物毒性的NLP蛋白(Nep1-Like proteins)成員更多。橡膠樹炭疽菌致病分子機理的研究,從突變體庫的構建、突變體的篩選、功能基因的克隆和分析,正逐步向調控機制、信號網絡分析及組學分析發展。
6 橡膠樹炭疽病的防治技術
近年來,對橡膠樹炭疽病防治技術的研究,主要集中在藥劑的篩選、劑型的研發、抗病育種、防治器械、生物防治等方面。生產上,重點防治歷年重病區和易感病品系林段橡膠樹炭疽病,結合預測測報,采用農業防治和化學防治相結合的防治措施。
6.1 預測預報橡膠樹嫩葉期為炭疽病易感期,每年春季橡膠新抽大量嫩葉時期,炭疽病容易和白粉病共同侵染,造成大量新抽嫩葉的落葉,因此,在此期間應加強病害的預測預報,尤其是歷年重病區和易感病品系的林段。一般從橡膠樹抽葉30%開始進行林段病情調查,若發現炭疽病發生,根據氣象預報在未來10 d內,有連續3 d以上的陰雨或大霧天氣,則要在低溫陰雨天氣來臨前噴藥防治。噴藥后從第五天開始,若預報還有上述天氣出現,則應在第一次噴藥后7~10 d內機械第二次噴藥,指導病情控制為止[14]。
6.2 農業防治對歷年重病林段和易感病品系林段,可在橡膠樹越冬落葉后到抽芽初期,施用速效肥,促進橡膠樹抽葉整齊,以減少侵入機會。在病害流行末期,對病樹施用速效肥,促進病樹迅速恢復生長[16]。苗圃地要注意不要在低洼積水地和山谷地建立苗圃,合理施肥,加強管理,使得膠苗生長健壯,提高膠苗的抗病能力[18]。
6.3 化學防治和防治器械根據預測預報結構進行噴藥防治,科學合理地輪換使用化學藥劑。目前防治的藥劑有28%復方多菌靈膠懸劑,用量為630 mL·hm?2, 兌水75 kg(每畝42 mL,兌水5 kg);20%滅菌靈膠乳劑,用量為450 mL·hm?2,兌水75 kg(每畝30 mL,兌水5 kg);80%的代森錳鋅兌水1 000倍液噴霧;10%百菌清熱霧劑、20%氟硅唑·咪鮮胺熱霧劑、15%咪鮮胺·晴菌唑熱霧劑、16%咪鮮·酮熱霧劑、30%嘧·咪·酮熱霧劑等[18,81?82]。于上午7:00前或下午7:00后,靜風時施藥防治,每隔7~10 d噴1次,噴2~3次[18]。
由于橡膠樹樹體高,可濕性粉劑和水劑等難以送達樹冠層,目前使用藥劑的劑型為粉劑或煙霧劑,防病機械包括噴粉機和煙霧機。中國熱帶農業科學院近年來開展了橡膠樹無人機植保飛防技術,利用無人機噴施硫磺粉防治具有施藥效率高、噴藥均勻、易控制和專業化等優點[83]。該實驗團隊稱,利用FBH300無人直升機每小時可以完成16.67 hm2(250畝)以上的作業面積,每公頃僅需要“保葉清”超低容量微乳劑加“熱飛”植物油飛防專用助劑60 L(每畝4 L),藥劑利用效率高,施用安全、高效[83]。
6.4 抗病育種品系的感病性是橡膠樹炭疽病發生的基礎,在病害流行頻率高的地區選種抗病高產品系,是預防橡膠樹炭疽病最經濟有效的方法。各植膠地均重視品系抗病性評價工作。我國主要采用室內離體接種鑒定法和大田調查法。華南熱帶作物學院肖倩莼通過苗圃鑒定和大田調查,從34個已推廣的橡膠品系中鑒定出對炭疽病具有較好抗性的品系有:“熱研44-9”、“熱研11-9”、“熱研88-13”、“熱研7-31-89”、“保亭933”、“南強1-97”[84]。蔡志英等[85]對我國46份我國大規模和中規模推廣品系進行田間2年評價,顯示表現抗性的品系有8份,為“IAN873”、“幼1”、“熱研8-79”、“熱研88-13”、“文昌217”、“云研77-2”、“云研77-4”、“云研277-5”。
6.5 生物防治目前,針對橡膠樹炭疽病的生物防治還在室內實驗或盆栽苗實驗探索過程中。賀春萍等[86?87]開展生防菌枯草芽孢桿菌Czk1與化學殺菌劑“根康”協同防治橡膠炭疽病,顯示Czk1與根康的協同作用在防治上效果均比施用單劑高;劉文波等[88]開展了解淀粉芽孢桿菌對橡膠炭疽菌的抑菌作用,等等。
7 展 望
橡膠樹炭疽病危害有逐年加重趨勢,目前是我國橡膠樹葉部重要“兩病”之一。現有研究結果表明,橡膠炭疽病田間危害癥狀、病原種類、病原菌生物學特性均呈多樣性。由于炭疽菌分類依據的變更,不同時期病原菌種鑒定結果不同,相應的研究結果是否具有很好的可比性值得商榷,如早期病原菌生物學特性的研究主要是對C. gloeosporioides和C. acutatum進行分析,隨著多基因序列分析法在炭疽菌種類中的應用,已知我國橡膠炭疽病病原菌歸屬3個復合群下的多個種,早期報道C. gloeosporioides和C.acutatum的生物學特性實為復合群的生物學特性。病原菌的準確鑒定及其對生物學特性等的了解可為田間防控提供指導。橡膠樹炭疽病危害部位和危害癥狀多樣,是否由不同的病原種引起?不同的橡膠樹栽培區,是否有不同的優勢種?不同炭疽菌種侵染橡膠樹的致病力和致病機制是否有差異?病原菌不同種間流行規律和對藥劑的敏感性是否有差異?不同寄主對不同種炭疽菌的抗病性如何?等等,都值得進一步分析。
炭疽菌種類繁多、遺傳多態性豐富。炭疽菌危害寄主多、地理分布廣,是重要病原真菌之一。近年對炭疽菌生長發育、致病和抗藥機制研究取得了較好的進展,今后有關分子機制研究仍是重點。關于橡膠炭疽菌的致病分子機制研究已從單個基因的功能研究逐步發展為信號網絡、調控機制分析及組學分析。分析橡膠炭疽菌致病機理,有望為橡膠樹炭疽病的綠色防控提供參考,也有助于促進人們更好理解植物病原菌炭疽菌的生長發育、致病、適生性等方面的分子機制。
目前,橡膠炭疽菌的防控技術仍以傳統的農業防治和化學防治為主。隨著技術的不斷創新,安全高效的防治技術需求,將快速推進無人機防治術、生物防治和選種抗病品系的發展。
