不同添加劑改善甜高粱青貯質量及其降解性能比較
2021-10-12 11:00:44任海偉劉玉龍馮銀萍張丙云李金平李志忠
農業工程學報 2021年14期
任海偉,趙 藝,劉玉龍,馮銀萍,張丙云,李金平,3※,李志忠
(1.蘭州理工大學生命科學與工程學院/西部能源與環境研究中心,蘭州 730050;2.甘肅省生物質能與太陽能互補供能系統重點實驗室,蘭州 730050;3.西北低碳城鎮支撐技術協同創新中心,蘭州 730050)
0 引 言
甜高粱是一種重要的 C4高糖分飼糧作物和能源作物,具有抗旱、耐澇、耐鹽堿、生物量高等特點,能適應多種氣候與環境條件,在中國甘肅、山東、黑龍江等地均有大面積種植,被廣泛用于動物飼料、生物質能(乙醇、沼氣等)和生物基材等領域[1-2]。由于甜高粱收獲具有明顯季節性和時效性,貯存不及時極易引起水分、糖分流失甚至腐敗變質,嚴重影響其能源轉化利用效率。青貯作為一種經典的濕法保存技術,不僅常用于作物秸稈等生物質原料的貯存[3],而且青貯過程還蘊藏有生化預處理作用[4-5]。研究表明,青貯過程中加入乳酸菌等微生物菌劑、酶制劑、木質纖維功能分解菌等添加劑能發揮一定的強化處理作用,提高生物質的降解性能和能源產出潛力[6]。纖維素酶及產生阿魏酸酯酶的乳酸菌能促進玉米秸稈青貯發酵和木質纖維素降解,提高酶解糖化得率[7]。稻草青貯過程中添加木聚糖酶、植物乳桿菌能提高發酵質量和纖維素轉化率[8]。另一方面,自然進化的鬼斧神工和農業生產智慧提示我們,“鮮活”的反芻動物瘤胃液、厭氧消化沼液等生物載體棲息有大量天然微生物菌群及其代謝酶系,是一類能有效克服木質纖維抗降解屏障的天然特異“生物處理系統”[9]。瘤胃液預處理稻草能有效促進其甲烷產量和有機質降解程度[9]。沼液預處理玉米秸稈能使其木質纖維結構變松散,降低化學反應活化能,加快纖維素分解[10-11]。更重要的是,瘤胃液、沼液這些生物載體還具備與濕法青貯過程進行耦合協同的共同環境基礎(厭氧、低pH值等)、物質基礎和代謝紐帶。若能將富含木質纖維分解菌的沼液、瘤胃液用于甜高粱青貯過程,則有可能通過生物強化作用實現貯存和強化預處理的雙重目標,以時間(貯存)爭取空間(降解糖化產出),提升生物質降解潛力。
甜高粱莖稈表面覆蓋有硅氧蠟質層,以及由木質素、半纖維素和纖維素形成的致密性晶體結構,使其不容易被分解。本文基于全株甜高粱的跨季貯存和能源化利用,從化學組分、發酵品質、微觀結構、酶解特性等方面,比較研究瘤胃液、沼液、纖維素酶和木聚糖酶 4種添加劑對甜高粱青貯品質的動態影響,并考察不同青貯周期時的生物降解潛力差異性,進而評價 4種添加劑在青貯過程中的強化作用效果,篩選適宜的、有應用價值的青貯添加劑。
1 材料與方法
1.1 試驗材料
全株鮮綠甜高粱取自中科院近物所武威種植基地,收獲后迅速切碎至1~2 cm備用。瘤胃液取自于武威頂樂生態牧業有限公司第三養殖場,西門塔爾肉牛屠宰過程中收集瘤胃液,經4層無菌紗布過濾后迅速轉移至厭氧袋,并存放至液氮罐中送回實驗室,-80℃保存備用;發酵沼液來源于蘭州牧工商公司荷斯坦奶牛繁育中心的沼氣工程,沼液取回實驗室后用高粱秸稈在(37±1)℃中溫環境下富集馴化一個月;纖維素酶和木聚糖酶(酶活力為50 000 U/g)購自寧夏和氏璧生物技術有限公司。甜高粱原料的干物質、有機組分和有機酸含量如表1 所示。新鮮沼液、瘤胃液的pH值、氨氮和有機酸含量、酶活性等指標如表2所示。
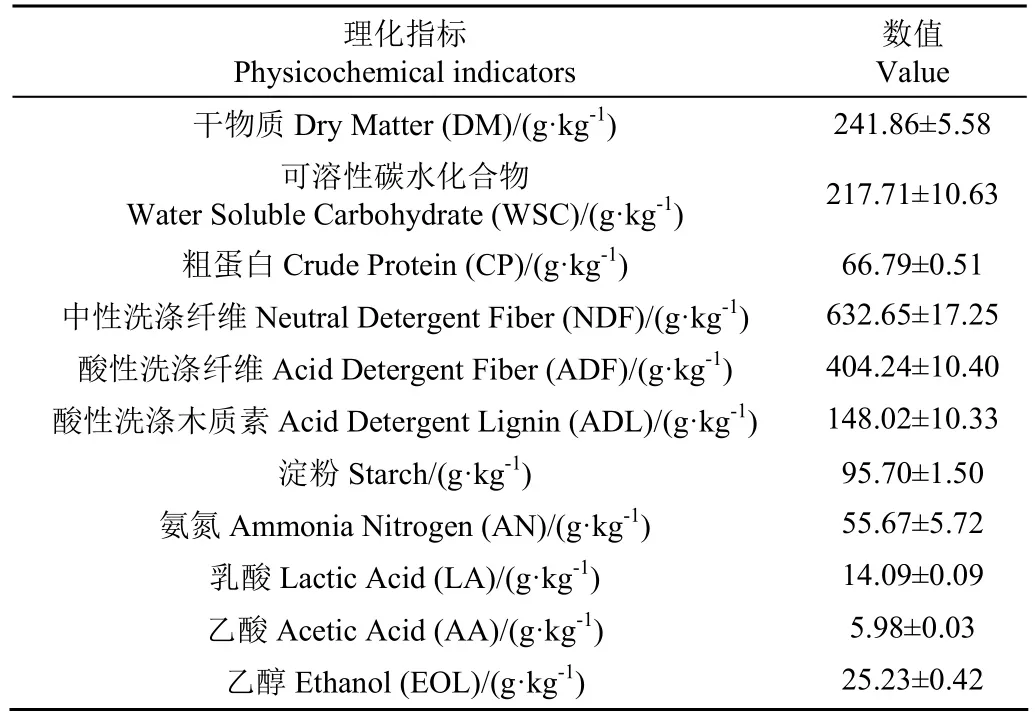
表1 甜高粱的理化指標Table 1 Physicochemical indicators of sweet sorghum
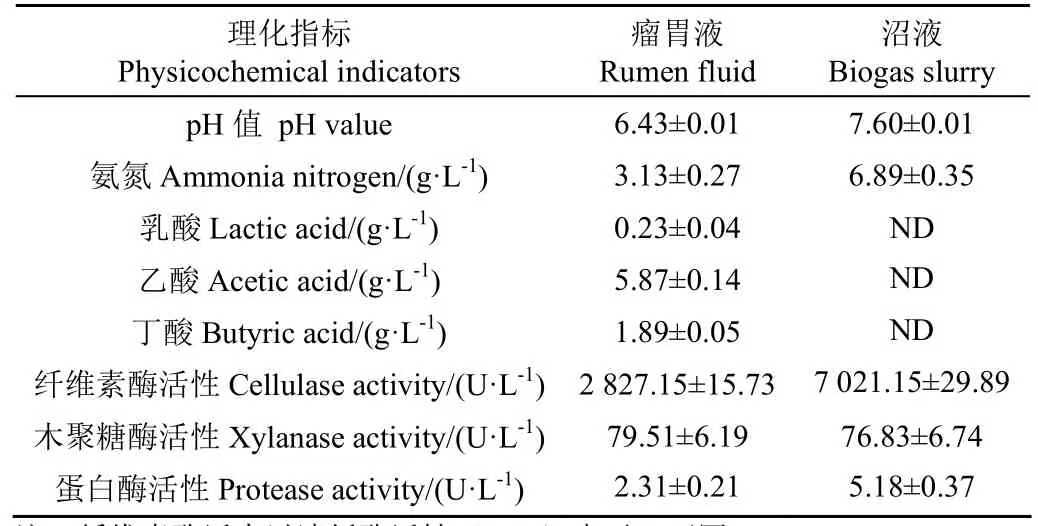
表2 瘤胃液與沼液的理化指標Table 2 Physicochemical indicators of rumen fluid and biogas slurry
1.2 儀器與設備
纖維分析儀(FB800),山東海能科學儀器有限公司;液相色譜儀(Agilent 1260),安捷倫公司;X-射線衍射儀(D/max-2400),日本理學公司;掃描電鏡(JSM-5600LV),日本電子光學公司;傅里葉變換紅外光譜儀(Nexus670),美國Nicolet公司。
1.3 青貯發酵試驗設計與取樣
準確稱取30份1.5 kg切碎的甜高粱進行青貯發酵試驗,設置纖維素酶(CT,3 g纖維素酶溶于70 mL蒸餾水)、木聚糖酶(XT,3 g木聚糖酶溶于70 mL蒸餾水)、瘤胃液(RT,添加量為70 mL/kg)、沼液(BT,添加量為70 mL/kg)4個處理組,以及1個空白對照組(CK,添加 70 mL蒸餾水),將添加劑均勻噴灑至樣品表面后迅速混勻并裝入塑料桶中密封。每個試驗組設3次平行,(18±2)℃恒溫環境中青貯45 d和90 d,定期分析青貯質量和生物降解性。青貯樣品的取樣步驟以及發酵浸提液制備方法參考文獻[12]進行。
1.4 青貯質量分析
1.4.1 化學成分和發酵品質
干物質(Dry Matter,DM)含量測定用105 ℃烘干恒重法,總氮(Total Nitrogen,TN)含量采用凱氏定氮法,可溶性碳水化合物(Water Soluble Carbohydrate,WSC)分析用硫酸-蒽酮法[13],淀粉含量測定采用酸水解法,中性洗滌纖維(Neutral Detergent Fiber,NDF)、酸性洗滌纖維(Acid Detergent Fiber,ADF)和酸性洗滌木質素(Acid Detergent Lignin,ADL)含量通過纖維分析儀測定。纖維素(Cellulose,CL)、半纖維素(Hemicellulose,HC)和綜纖維素(Holocellulose,HoC)組分通過公式計算[14]。pH值采用 UB-7酸度計分析,氨氮(Ammonia Nitrogen,AN)采用苯酚-次氯酸鈉比色法分析,有機酸(乳酸、乙酸、丙酸、丁酸)等小分子發酵產物分析采用HPLC檢測[7,15]。纖維素酶、木聚糖酶、蛋白酶活性的測試均采用南京建成生物工程研究所采購的試劑盒,具體操作參照說明步驟進行。
1.4.2 結構特性表征
利用掃描電鏡觀察樣品的微觀形貌(噴金儀噴鍍40 s,鍍膜厚度4 nm左右,加速電壓20 kV)。利用傅里葉變換紅外光譜儀分析樣品官能團結構(掃描范圍400~4 000 cm-1,分辨率4 cm-1)。采用Rigaku X-射線衍射儀分析結晶性能(Cu-Kα 為靶材,掃描速率為 1°/min,掃描范圍5°~60°,加速電壓 40 kV,電流 100 mA),并計算相對結晶指數[14]。
式中 CrI為相對結晶指數的百分率(%);I002為結晶區的衍射強度(2θ=22°);Iam為無定形區衍射強度(2θ=18°)。
1.5 生物降解性能評價
生物降解性能主要從理論生物降解潛力(Theoretical Biodegradation Potential,TBP)和實際酶解糖化得率兩方面進行評價。TBP為綜纖維素和木質素組分含量之比值(干物質基礎)。酶解糖化能力評價方法:準確稱取0.5 g甜高粱原料或青貯樣品(過100目篩),以1∶20料液比加入pH值4.8、濃度為0.05 mol/L的檸檬酸緩沖液,再依次添加纖維素酶(加酶量1 000 U/g)、半纖維素酶(加酶量500 U/g)、β-葡聚糖苷酶(加酶量1 U/g)進行酶解試驗。恒溫氣浴振蕩酶解72 h(150 r/min,(50±0.5 )℃),間隔12 h測定酶解液中的還原糖濃度,并計算酶解得率(Enzymatic Hydrolysis Yield,EHY,%)[16]。
式中c為還原糖濃度,g/mL;v為上清液體積,mL;m為底物質量,g。
1.6 數據處理與統計分析
用Excel 2007軟件處理基礎數據,結果用平均值±標準差表示,使用Origin Pro 9.1軟件繪圖,SPSS 20.0軟件進行雙因素方差分析,單因子 ANOVO模型處理及Duncan方法對數據進行多重比較分析。
2 結果與討論
2.1 添加劑瘤胃液和沼液的微生物菌群分析
利用Illumina HiSeq 2500 平臺分析瘤胃液、沼液中的細菌菌群多樣性(表3)。

表3 瘤胃液和沼液的門、屬水平上細菌多樣性Table 3 Bacterial diversity of rumen fluid and biogas slurry at phylum and genus level
由表3可知,瘤胃液的門水平細菌主要有厚壁菌(Firmicutes,61.00%)和擬桿菌(Bacteroidetes,31.10%),沼液中的優勢細菌除厚壁菌(Firmicutes,32.58%)和擬桿菌(Bacteroidetes,35.08%)外,還含有一定量互養菌(Synergistetes,17.12%)。研究表明,大多數厚壁菌和擬桿菌的細菌菌群均有木質纖維水解和蛋白質降解能力,且二者存在一種相互促進的共生關系[17]。互養菌還能發酵產生有機酸和乙醇,也是一類重要的木質纖維分解菌[18-19]。從屬水平看,瘤胃液的優勢細菌屬主要為克里斯滕森氏菌(Christensenellaceae,20.09%)和瘤胃球菌(22.01%)。這些細菌區系均能有效降解木質纖維素,而且前者還有發酵糖類產生乙酸、丁酸等有機酸的能力[20-21]。沼液的優勢細菌主要有理研菌(Rikenellaceae,26.89%)和少量梭菌(Clostridium,5.44%)、嗜蛋白菌(Proteinip-hilum,4.00%)和假單胞菌(Pseudomonas,2.02%)。其中,假單胞菌具有攻擊芳香族聚合物木質素的能力,盡管其相對豐度相對較低,但它對木質素降解仍有積極作用[10]。
2.2 添加劑對青貯過程中化學組分的影響
2.2.1 干物質含量和干物質損失率
干物質含量是評價甜高粱青貯品質優劣的重要指標。由表1和圖1可知,與原料相比,青貯45 d時5個試驗組的干物質含量均顯著下降(P<0.05),尤其RT組處于最低值;青貯45~90 d期間,CT、XT和BT處理組的干物質含量均隨時間延長而顯著降低(P<0.05),而CK和RT組在此期間未發生明顯變化(P>0.05)。這與Naeini等[16]報道的青貯甜高粱干物質含量隨時間延長而呈下降趨勢結果基本吻合。分析原因,一方面,加入的纖維素酶、木聚糖酶等生物酶和沼液(富含木質纖維分解菌)在青貯發酵期間持續發生作用,將部分有機物質分解為諸如小分子有機酸、乙醇和含氮化合物等揮發性物質,使甜高粱中的干物質含量顯著下降。另一方面,瘤胃液中含有大量諸如產琥珀酸絲狀桿菌、黃色瘤胃球菌、白色瘤胃球菌等木質纖維降解菌以及可以穿透植物細胞壁的真菌,多種微生物菌系協同作用使一部分纖維素和半纖維素快速分解,并轉化為乙酸為主的揮發性有機酸[9]。但隨著發酵時間延長,由于青貯體系pH值下降、乳酸菌等優勢菌群占主導、瘤胃液中噬菌體等微生物病毒發揮抑制作用等諸多因素使得木質纖維分解菌被抑制,從而減少干物質分解,故其含量變化不顯著(P>0.05)。
再者,BT組在90 d時的干物質含量顯著低于CK組(P<0.05),但與其他處理組差異不顯著(P>0.05),這不僅是因為沼液中的微生物菌群及其酶系將淀粉、蛋白質和木質纖維素等組分降解轉化;而且沼液中豐富的氨氮可作為天然氨源,使木質素內部以及木質素與多糖之間的化學鍵斷裂,進而導致木質素膨脹并與纖維素分離,提高纖維素與半纖維素的可降解性,從而使干物質損失率增加(圖1b)[10,22]。
2.2.2 可溶性碳水化合物含量
可溶性碳水化合物(Water Soluble Carbohydrates,WSC)是青貯發酵過程中乳酸菌群繁殖代謝的重要底物,含量多寡及其變化規律一定程度上也能反映不同種類添加劑在青貯過程中的強化作用效果。由圖2和表1可知,青貯45 d時5個試驗組的WSC含量均顯著低于原料(P<0.05),且CT和XT組的WSC含量顯著低于其他試驗組(P<0.05),BT組的WSC含量最高,這可能是因為活性較強的沼液中富含木質纖維素分解菌及纖維素酶等復合酶系,相較于纖維素酶或木聚糖酶等單一酶制劑而言,更能充分有效地分解甜高粱中纖維素、半纖維素、淀粉等大分子碳水化合物,使WSC生成速率高于消耗速率,形成可溶性糖的累積效應[9]。此外,沼液還能促進秸稈木質纖維素結構的膨脹崩解,使其通透性增加,更容易被微生物及其酶系降解[22]。另一方面,青貯90 d時CT組的WSC含量較45 d時明顯增加(P<0.05),這可能是纖維素酶能降解秸稈中不與木質素存在化學鍵聯結的細胞壁成分所致;而且青貯環境中乳酸菌占優勢、腐敗微生物菌群被抑制等因素也會使CT組中WSC消耗速率降低,導致WSC積累增加。
總之,整個青貯發酵期間,RT和 BT組中的 WSC含量始終高于其他試驗組,尤其BT組在45和90 d時均為最高值,說明甜高粱青貯體系中添加沼液的生物強化作用效果最為明顯。
2.2.3 木質纖維組分和淀粉含量
由表4可知,添加劑、時間及其交互作用均對木質纖維組分有極顯著影響(P<0.001)。中性洗滌纖維(Neutral Detergent Fiber,NDF)包括了植物細胞壁的大部分成分,主要有不溶性纖維素、半纖維素等非淀粉多糖和木質素。與原料相比,45 d時各試驗組(除BT組)的NDF含量均未發生顯著變化,但在 90 d時顯著低于原料(P<0.05),說明無論是否加入添加劑或何種種類,長時間青貯發酵均有助于NDF組分的降解。尤其添加沼液的BT組中NDF組分含量隨時間延長呈顯著下降趨勢,且在整個青貯期間均顯著低于其他處理組(P<0.05)。半纖維素(Hemicellulos,HC)是組成植物細胞壁的結構性多糖,由不同類型單糖組成,常與纖維素和木質素緊密交聯形成木質纖維素,該組分的含量變化也存在與纖維素相類似的趨勢。因為沼液中本身存在有大量腐殖酸和可降解木質纖維素的微生物菌群,能使難降解的高聚體木質素以及被包裹的纖維素和半纖維素發生分解[23],這些組分的降解都會使NDF含量隨之下降。
酸性洗滌木質素(Acid Detergent Lignin,ADL)是影響纖維素和半纖維素等結構性碳水化合物降解的主要屏障,較高的 ADL 含量會導致生物降解性能下降,進而影響酶解糖化效率。與原料相比,5個試驗組在青貯期間的 ADL含量均明顯下降(P<0.05)。尤其,RT組和BT組中的 ADL含量在 90d時分別降至 78.11 和79.01 g/kg,顯著低于其他處理組(P<0.05),這得益于瘤胃液和沼液天然具備的木質纖維分解能力,二者富含能降解植物細胞壁的微生物菌群及其酶系[23]。這與 Guo等[10]報道的玉米秸稈經沼液預處理后木質纖維素明顯下降的結果一致;Candia-García等[24]也發現瘤胃液能明顯降低稻草中的木質纖維組分。可見,青貯過程中加入沼液、瘤胃液能發揮明顯的生物強化預處理作用,有效降低抗降解屏障組分ADL含量,促進半纖維素降解,最大限度地保存纖維素組分[11]。這與纖維組分隨青貯時間延長而顯著升高的變化趨勢相吻合(表4)。此外,上述木質纖維組分的聯動變化也間接影響能量組分綜纖維素的含量多寡。青貯90 d期間,BT組中的綜纖維素含量最低,RT組中的綜纖維素含量最高,因為瘤胃液中的纖維分解菌對pH值敏感,在酸性青貯環境中活性較弱,降解作用趨于溫和;而沼液中的纖維分解菌對pH值變化適應性較好,使綜纖維素組分的含量處于最低值。
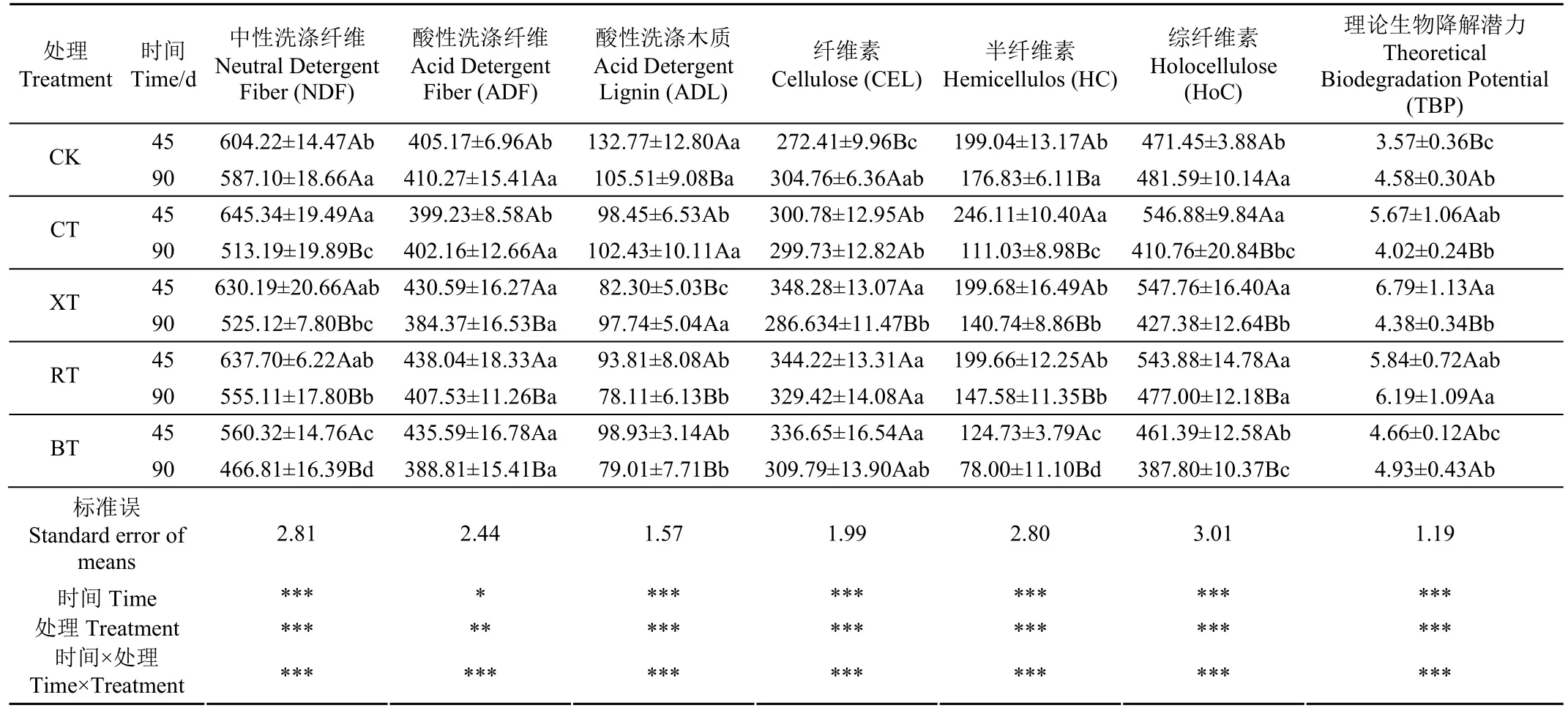
表4 青貯過程中木質纖維組分的變化Table 4 Changes of lignocellulosic components during ensiling (g·kg-1)
結合理論生物降解潛力(TBP)分析,青貯45 d時,4個添加劑組的TBP均明顯增加(P<0.05),而CK組則無明顯變化(P>0.05)。青貯90 d時,CT和XT組的TBP不增反降,而RT和BT則保持不變。說明,添加劑的加入均有助于提升青貯甜高粱的理論降解潛力,起到明顯的強化處理作用,而且添加瘤胃液、沼液的作用效果相對較好,但實際改善效果需通過酶解糖化試驗進行驗證。
淀粉是全株甜高粱中重要的非結構性碳水化合物。由圖3和表1可知,青貯45 d時,CK和CT組的淀粉含量與原料相比差異不顯著(P>0.05),而XT、RT和BT組的淀粉含量在青貯45 d時顯著高于原料。然后,5個試驗組的淀粉含量均隨青貯時間延長而呈顯著下降(P<0.05),90 d時的5個試驗組的淀粉含量均明顯低于原料(P<0.05),但組間差異不明顯,說明4種添加劑對淀粉含量的影響無顯著性差異(P>0.05)。
2.3 添加劑對青貯發酵品質的影響
2.3.1 pH值
青貯pH值的下降速率和程度是反映發酵品質優劣的重要指標[25]。由圖4可知,發酵45 d時,由于青貯體系內乳酸菌等有益菌及其他微生物的厭氧生化反應,將可溶性糖轉化為乳酸(pKa=3.86)和乙酸(pKa=4.75)等有機酸[26],使5個試驗組的pH值均迅速下降至3.67~3.90范圍,明顯低于原料(pH 值5.62),均達到優良青貯標準(3.8~4.2)。低 pH值能有效抑制蛋白酶活性,減少蛋白質降解,抑制對青貯有害細菌的生長,從而保障青貯品質。當青貯周期延長至90 d時,CK、CT和XT組的pH值繼續顯著下降(P<0.05),維持在 3.52~4.29;RT和 BT組則呈顯著增加趨勢(P<0.05),pH值分別為 4.29和 3.93。分析原因,纖維素酶、木聚糖酶對結構性碳水化合物的直接催化降解作用進一步促進了乳酸等有機酸的發酵累積,導致青貯pH顯著下降[17,27]。另一方面,瘤胃液、沼液因其特有的微生態菌群結構,容易生成較高含量的乙酸[9];而且青貯發酵生成的乳酸又會被反過來被沼液、瘤胃液中微生物菌群作為代謝底物而消耗減少[28],導致乳酸和乙酸含量的此消彼長,這些因素的疊加造成了RT和BT組pH值隨時間延長而顯著升高,這與表5中的有機酸變化趨勢相吻合。
2.3.2 氨氮含量
青貯蛋白質的水解通常是由植物蛋白酶將其水解為肽和游離氨基酸,然后通過微生物菌群活動進一步降解為酰胺、胺和氮等因素引起的。
由圖5可知,青貯90 d期間,各試驗組(除CT組不變外)的氨氮含量均隨時間延長而顯著下降(P<0.05),這可能是因為較低的酸性 pH青貯環境有效抑制了芽孢桿菌、沙門氏菌等腐敗菌生長以及甜高粱植株中的蛋白酶活性,從而減少蛋白質分解[29];添加纖維素酶對限制氨氮的產生并無顯著影響,這與Guo 等[30]研究結果一致。另一方面,青貯45 d時RT和BT組中氨氮含量顯著高于其他試驗組(P<0.05),但仍低于優質青貯的氨氮限值(10%)。瘤胃液、沼液中自身本底含有一定量的氨氮,而且這兩類添加劑也富含蛋白質降解菌,極易引發蛋白質分解和氨氮增加[9]。當青貯周期延至90 d時,RT和BT組中的氨氮含量又反而處于較低水平,這與青貯體系中較高濃度乙酸等抗菌物質有效抑制蛋白質分解菌群存在一定聯系[31]。
2.3.3 有機酸及乙醇含量
小分子有機酸和醇類物質是青貯發酵過程中微生物菌群的主要代謝產物。由表5可知,隨著青貯時間的延長,CK、CT和XT組中的乳酸含量顯著增加(P<0.05),這是由于外源纖維素酶、木聚糖酶的加入直接促進了纖維素、半纖維素等結構性碳水化合物的催化分解,進而通過優勢乳酸菌群發酵產生并積累乳酸所致[27]。這與Zhao等[17]、陳鑫珠等[32]在柳枝稷、甘蔗梢青貯中的研究結果吻合。相反,RT和BT組中的乳酸含量隨時間延長而顯著下降(P<0.05),這主要與瘤胃液微生態系統中的乳酸代謝菌能將其轉化為乙酸、丙酸等物質有關。此外,優勢乳酸菌群從同型發酵類型向異型發酵的轉變也可能是原因之一[33]。另一方面,5個試驗組的乙酸、丙酸含量均隨時間延長而顯著升高,且整個青貯期間 CT、XT和BT組中二者的含量均顯著低于CK和RT組(P<0.05),而在RT組中含量則相對較高。這可能是因為RT組中加入的瘤胃液存在大量產琥珀酸絲狀桿菌、黃色瘤胃球菌、白色瘤胃球菌以及少量梭桿菌,這些微生物在厭氧條件下能產生較多的乙酸和丙酸[9]。Zhang等[34]也發現稻草厭氧發酵過程中添加瘤胃液,乙酸含量明顯提高。另外,青貯90 d時RT和BT組中檢測出少量丁酸,這可能是由于瘤胃液、沼液中的氨氮物質會促進梭狀芽孢桿菌生長,從而產生具有損耗青貯品質的丁酸所致[9,11,17]。乙醇也是甜高粱青貯過程的重要產物,和乙酸類似均能起到一定抑菌作用,抑制腐敗菌生長,有助優勢乳酸菌占據主導[23]。整個青貯期間,4個添加劑組的pH值均低于4.3,乳酸/總有機酸(乳/總)值始終高于0.58,乳酸/乙酸(乳/乙)值始終大于2.0,均達到優良青貯發酵范疇[35](除個別組),乳酸菌群發酵占主導,有效抑制了酵母菌繁殖代謝,從而顯著減少了乙醇含量(P<0.05)。其中,CT和XT組的乳/總和乳/乙比值均顯著高于CK組,而RT和BT組的這兩個比值在45 d時顯著高于CK組,90 d時卻顯著低于CK組,這可能與瘤胃液、沼液中含有復雜多樣的微生物菌群有關,這些微生物群落的引入打破了原本相對簡單的青貯乳酸發酵型態,將乳酸作為代謝底物利用轉化為乙酸、丙酸等小分子揮發酸[36]。
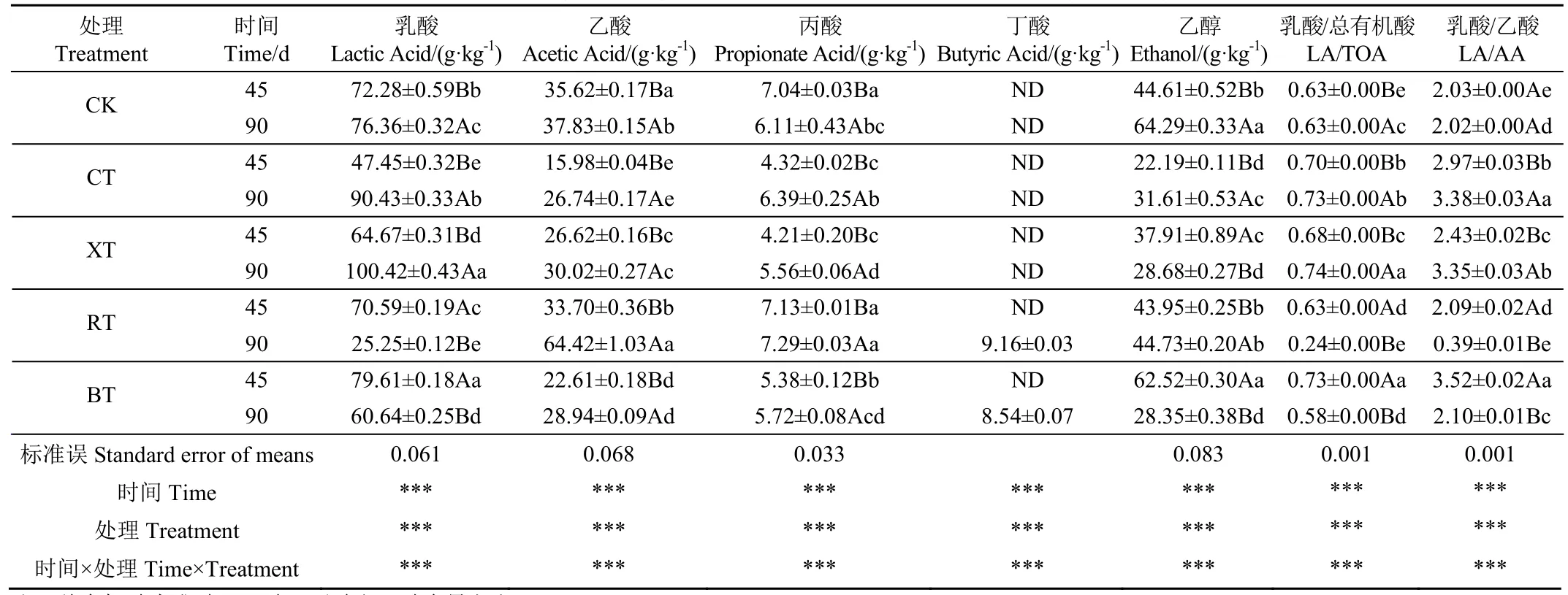
表5 青貯過程中有機酸與乙醇含量的變化Table 5 Changes of organic acid and ethanol contents during ensiling
2.4 甜高粱青貯前后的結構表征
2.4.1 微觀形貌分析
由圖6可知,未青貯的甜高粱莖稈結構較為致密,表面光滑平整,角質層形成的包埋結構無孔隙裂痕。青貯45 d時,所有試驗組的甜高粱表面莖稈結構均遭受破壞。其中,CT和XT組的木質纖維結構發生微弱瓦解,RT和BT組則出現大量微孔洞、裂痕甚至斷裂分層現象,形成許多凹凸不平的溝槽,并呈現條紋鋸齒狀,說明青貯過程中添加瘤胃液、沼液能更有效地破壞甜高粱木質纖維結構。因為瘤胃液和沼液自身富含復雜多樣的古菌、真菌及原蟲等微生物菌群,青貯發酵期間能穿透莖稈表面的蠟質層和木質化細胞壁,破壞莖稈內部半纖維素和木質素之間的酯鍵并形成裂痕[9]。
隨著青貯周期的延長,90 d時五個試驗組的孔洞、裂縫愈發明顯,尤其添加沼液的BT組出現大面積崩解塌陷,這是因為甜高粱在青貯過程中除受到微生物菌群的持續作用外,長時間的沼液浸潤也能使木質纖維結構膨脹崩解,這與Guo等[10]結果相吻合。
2.4.2 官能團分析
由圖7紅外光譜分析可知。
6個甜高粱樣品的紅外光譜圖總體表現出相似特征,明顯吸收峰值主要發生在3 413、2 926、1 733、1 604和1 050 cm-1。其中,3 413 cm-1處的寬峰代表纖維素羥基的伸縮振動[37],相較于未青貯的原料(SS組)而言,5個青貯樣品在3 413 cm-1處的吸收峰強度均有所減弱;青貯45 d時BT組的吸收峰強度明顯低于其他處理組。說明纖維素組分在青貯期間均有不同程度地降解,其中青貯過程中添加沼液進行強化處理可在較短時間內有效降解或破壞甜高粱莖稈的結晶纖維素結構,使其發生分解作用。2 926 cm-1處的特征吸收峰表征木質素中甲基、亞甲基(C-H)的伸縮振動[37],青貯90 d時4個處理組在此處的吸收峰均明顯低于45 d,說明青貯周期的延長對木質素脫除具有積極效果,且與青貯周期呈正相關。1 733和1 050 cm-1分別代表半纖維素-木質素復合物的特征吸收峰以及半纖維素中C-O鍵的伸縮振動[38-39]。青貯90 d時,RT、BT和XT組在這兩處的吸收峰與45 d相比明顯減弱,說明青貯周期延長后瘤胃液、沼液對木質素以及半纖維素的解聚效果明顯增強,青貯過程中添加木聚糖酶對半纖維素組分分解、木質素組分釋放解聚有積極效果。1 604 cm-1處的伸縮振動來源于吸附水分子氫鍵的彎曲振動[22]。青貯后所有試驗組在該處吸收峰強度均有不同程度地升高,且隨青貯時間延長強度逐漸增強,說明青貯能促進纖維素無定形組分的降解。
2.4.3 X-衍射分析
如圖8所示,與原料相比,五個青貯組的甜高粱樣品在 2θ=22°左右均出現強吸收峰,在 2θ=9°和 2θ=17°左右出現弱峰,說明加入不同添加劑的強化作用效果和厭氧生化反應能使甜高粱莖稈中纖維素組分發生明顯的結構重定向。結合相對結晶指數(CrI)可知,四個添加劑處理組的 CrI指數與 CK組和原料相比均有明顯下降,其中BT組降幅最大,說明外源添加劑的加入均能有效破壞纖維素和半纖維素與木質素緊密結合的酯鍵與氫鍵;添加劑沼液中存在的高濃度氨氮更能進一步促進纖維素與木質素的剝離[40]。另一方面,90 d時XT、RT和BT組中的纖維結晶指數較45 d有所提高(表6),這可能是因為青貯時間的延長使非纖維素多糖或無定形區被溶解,導致纖維素結晶度增加[8]。此外,整個青貯期間BT組的CrI指數始終處于最低值,除了沼液中豐富的微生物菌群及其木質纖維分解酶系所帶來的生物強化預處理作用外,沼液中豐富的氨氮組分對木質纖維結構的分解也可能是該處理組CrI[10]
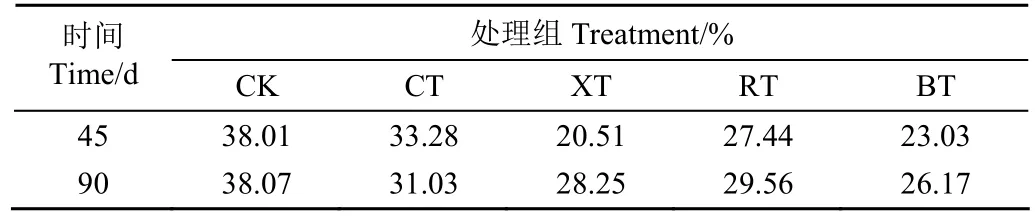
表6 甜高粱青貯前后的相對結晶指數變化Table 6 Changes of crystallization index of sweet sorghum before and after ensiling
2.5 甜高粱青貯前后的酶解糖化效果評價
酶解糖化效果是評價木質纖維生物質降解能力優劣的重要指標[39]。如圖9所示,無論甜高粱青貯與否,6個樣品底物在酶解初期(<12 h)的酶解得率均呈現快速增加、直線上升趨勢,12h后酶解得率增速放緩。這是因為該階段甜高粱中的纖維素和半纖維素組分優先被生物酶作用釋放出較多的可發酵糖。隨著酶解進程的推進,木質纖維底物的降解難度逐漸增加,而且還存在還原糖產物的反饋抑制作用,從而使酶解得率增速放緩[41]。另一方面,青貯甜高粱的酶解得率均顯著高于原料,說明青貯發酵能通過改善甜高粱木質纖維結構、優化化學組分構成、打破碳水化合物-木質素之間聯系等形式促進木質纖維組分被酶解,進而改善其酶解糖化性能。此外,不同種類添加劑的強化作用效果也不盡相同,由于瘤胃液和沼液中含有大量木質纖維分解菌及水解酶類,使BT和RT組的酶解得率明顯高于CT和XT組。尤其青貯90 d時的BT組酶解糖化得率最高達84.69%,長時間的酸性青貯環境、沼液中木質纖維分解菌等微生物菌群、較高濃度的氨氮等因素都可能是有效提升酶解效能的重要原因,這些因素相互疊加起到明顯的生物強化處理作用[10,22]。
3 結 論
1)纖維素酶、木聚糖酶、瘤胃液和沼液4種添加劑均能使青貯體系的pH值顯著下降至4.3以下,除添加瘤胃液青貯90 d外,其他處理組的乳酸占總有機酸比例均高于0.58,乳酸與乙酸比值均高于2.0,說明四種添加劑均能有效促進乳酸發酵,改善發酵質量和青貯品質。通過強化青貯作用,還能有效促進木質纖維組分分解及其優化重組,進而提高酶解得率和生物降解性能,其中甜高粱添加沼液青貯90 d時的酶解糖化得率最高,達到了84.69%。
2)青貯甜高粱的微觀結構發生了有利于生物降解性能提升的積極變化,尤其沼液、瘤胃液富含木質纖維分解菌群,能在青貯發酵的同時有效瓦解甜高粱莖稈的木質纖維抗降解屏障結構,莖稈微觀表面出現明顯孔洞和裂縫,使纖維素相對結晶度指數明顯降低。尤其是沼液處理組的相對結晶指數在整個青貯期間均處于最低值,分別為 23.03和26.17。因此,在研究使用的4種添加劑中,添加沼液對甜高粱青貯過程的生物強化作用效果最佳。
3)綜合考慮4種添加劑的資源量、易得性、成本以及對甜高粱青貯質量和生物降解性能的改善效果等因素,建議選擇來源廣泛、廉價易得、資源豐富的沼液作為甜高粱青貯過程的添加劑。這對于以甜高粱為原料的沼氣工程運行具有重要意義,不僅能實現原料的可持續供給,提高生物降解性,還有助于消納轉化沼氣工程排放的一部分沼液,減少沼液排放量。
