棉花幼苗內源保護酶對高溫與漬水脅迫的響應
2021-10-11 02:52:26郭佳黃麗楊盼王海苗陸泉泉盧秋美冉莉琴
南方農業·上旬 2021年9期
郭佳 黃麗 楊盼 王海苗 陸泉泉 盧秋美 冉莉琴

摘 要 為量化“中棉所45”棉花幼苗內源保護酶對高溫和漬水雙重脅迫的響應,采用盆栽試驗,以正常生長情況為對照(CK),設置3種脅迫:高溫(HT)、漬水(WL)、高溫+漬水(HT+WL)互作處理;處理前后測定棉花幼苗的形態指標、SPAD值、內源保護酶活性和丙二醛(MDA)含量,分析各處理間差異。結果表明:1)HT、WL和HT+WL脅迫7 d后,棉花株高和真葉相對含水量較對照顯著降低。2)與處理前相比,各處理棉花葉片SPAD值顯著下降;棉花根和子葉中的超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、過氧化物酶(POD)活性和MDA含量增加,其中子葉SOD活性分別增長了15.00%、9.84%和40.93%,子葉CAT活性分別增長了22.6%、76.3%和100.2%,子葉POD活性分別增長了36.23%、56.59%和88.16%。3)高溫與漬水雙重脅迫對棉花幼苗中內源保護酶活性具有顯著疊加效應及互作效應,且根響應脅迫較子葉敏感。
關鍵詞 棉花幼苗;高溫脅迫;漬水脅迫;內源保護酶;丙二醛
中圖分類號:S562 文獻標志碼:A DOI:10.19415/j.cnki.1673-890x.2021.25.003
棉花,在我國農作物經濟發展上具有重要地位。據相關報道,在全球棉花產量中,我國棉花產量占1/4[1]。我國棉區劃分為三大區域,包括黃河流域棉區、西北內陸棉區、長江流域棉區,其中,長江流域棉花種植區氣候屬于亞熱帶季風氣候,常會出現洪澇、梅雨漬害等災害天氣。棉花生長發育需要經歷幼苗期、現蕾期、花鈴期、吐絮期四個階段,每一階段對棉花產量質量都有重要的影響,其中幼苗期是棉花生長發育最基礎的階段。1)棉花喜光熱[2],耐澇漬性差,整個生育期需水量少[3],因此,即使是短期的澇漬也會對棉花的生長產生極大的持久影響[4]。2)過高的溫度會嚴重影響棉花的長勢、葉片含水量、光合能力和內源保護酶活性等正常的生理活動,最終影響棉花產量品質。3)棉花還在較大程度上受到其他非生物因素如光照、澇害等不可控的自然條件的影響。有研究表明,氧化脅迫是植物遭受環境脅迫的主要危害因素,ROS(活性氧)代謝在植物交叉適應各種非生物脅迫中起著重要作用,植物的交叉耐受性與抗氧能力之間存在正相關[5-7]。前人對于植物受到不適溫度或者水分等非生物因素脅迫后,植株生理響應機制的研究頗豐,對于棉花植株在不同時期受到旱澇[2-4,8-9]、高溫[10]、低溫[11-12]、鹽分[13-15]或者金屬離子[16-17]等非生物因子脅迫后的生理機制研究也很多。但針對中國農業科學院棉花研究所育成的早熟高產優質棉花新品種“中棉所45”苗期受到短期的澇漬和高溫雙重脅迫與單一脅迫后,對植株的根和子葉內源保護酶不同影響的量化對比鮮有報道。
本試驗以中棉所45為材料進行模擬雨熱同期的環境脅迫,以期量化苗期棉花對HT與WL單一脅迫,以及雙重脅迫對植株不同部位的響應變化特征,為大田生產苗期棉花的育苗、保苗、篩選苗株等措施提供有用的參考信息,為棉花高抗性品種選育提供理論基礎。
1? 材料與方法
1.1? 供試材料
2019年在廣西桂林(110°29′E,25°10′N)廣西師范大學伯康樓恒溫光照培養箱進行盆栽試驗,盆高7.5 cm、直徑9 cm,盆底有排水孔。供試材料為中棉所45 (生育期135 d)。栽培土壤取自有機營養土,含有泥炭土、珍珠巖、蛭石等,有機質含量約占65%。
1.2? 試驗處理
本試驗共設置4個處理:HT(高溫)、WL(漬水)、HT+WL(HT+WL)、CK(正常條件下生長)。試驗采用隨機區組設計,每個處理設置3個重復,每個重復15盆,每盆3株棉花苗株。
5月7日將中棉所45(生育期128~135 d)種子播種于小盆中,3粒/盆,距離均相等。將營養土和自來水混勻后往花盆里裝土,裝后壓實,直到與小盆口徑平行,各小盆所裝土壤體積相同。將小盆以3×5(每個中轉箱可裝15個盆)整齊排列盛放于事先準備的中轉箱中,放入溫光培養箱中進行培養。后期選用Hoaglands營養液進行澆灌。
棉花出苗后,生長至高約5 cm,用蒸餾水將營養液稀釋100倍,每隔3天澆灌1次,每次10 mL。在棉花生長到三葉一心期時,開始進行處理。對照為自然溫度25/20 ℃。HT脅迫處理將幼苗置于恒溫溫光培養箱中35/30 ℃條件下處理,光照時間6:00—18:00,相對濕度控制在65%左右。處理時間為7月20—27日,共持續7 d。HT處理結束后,將棉花植株移出恒溫溫光培養箱,將其置于與對照組同一自然環境下并及時通風。WL處理、HT處理、HT+WL互作處理同期進行。WL期將塑料盆放入含水的中轉箱內,以小花盆內土壤相對含水量(75±5)%為對照(CK),設置土壤WL 7 d(7月20 日),WL期間保持中轉箱內有6 cm水層。WL處理結束后排除中轉箱中的水,然后將塑料盆原位放置回試驗區,使土壤相對含水量降低至(75±5)%,并維持在此水平。HT+WL互作處理將盆栽放入含水層為6 cm的中轉箱中,并置于恒溫溫光培養箱中35/30 ℃條件,持續7 d。在HT、WL脅迫條件處理前后進行取樣,測定棉花各生長指標、SPAD值、MDA含量、內源保護酶(SOD、CAT、POD)活性。
1.3? 測定項目及方法
1.3.1? 形態學指標測定
株高(cm):用直尺測量棉花在各處理后的株高,每個重復區域呈對角線定點取3盆作為待測樣品,共9株棉花,每個處理3個重復。
真葉相對含水量(%):于各處理結束后第二天早上10:00,每個重復區域定點取9株棉花,稱棉花真葉鮮重(FW)。然后將葉片浸入裝滿水的培養皿中浸泡10 h至葉片吸水飽和,輕輕擦干棉花葉片表面水分,稱葉片飽和重(TW)。隨后將樣品放入牛皮紙袋中,貼好標簽,105 ℃殺青30 min,80 ℃烘干至恒重后稱干重(DW),按照公式(1)計算真葉的相對含水量(RWC,%):
RWC=[(FW-DW)/(TW-DW)]×100% (1)
1.3.2? SPAD值、丙二醛含量與內源保護酶活性測定
HT、WL 、HT+WL和CK處理前一天與處理結束后第2天早上9:00,從各處理的每個重復中選取3盆長勢一致的棉花,分別取其根、葉(真葉和子葉),測試分析上述指標。
真葉葉綠素含量,采用SPAD儀測定;不同部位的丙二醛(MDA)含量,硫代巴比妥酸比色法;不同部位的超氧化物歧化酶(SOD)活性,氮藍四唑法;過氧化氫酶(CAT)活性,分光光度計法;過氧化物酶(POD)活性,愈創木酚法[18]。
1.4? 數據統計及分析
利用 Microsoft Excel 2007軟件進行試驗數據的整理和圖表制作,應用 SPSS 20.0對各處理間和處理前后數據進行方差分析。
2? 結果與分析
2.1? 苗期棉花特征
處理前植株生長狀態一致。表1列出了對照(CK)、高溫(HT)、漬水(WL)及高溫和漬水(HT+WL)互作處理7 d后,棉花的株高和真葉相對含水量。從表1可知,HT處理、WL處理、HT+WL互作處理對棉花的地上部生長產生抑制效應導致植株生長緩慢且真葉相對含水量下降。具體表現為與CK相比,三種處理后株高分別降低0.91%、9.36%、11.36%,棉花真葉含水量分別降低20.46%、19.81%、24.95%。其中,HT+WL脅迫處理下株高降幅較HT處理高8.45%、較WL處理高2.0%。由此可知HT和WL對株高有顯著交互作用,而影響棉花株高的首要因子是WL。
同列不同小寫字母表示在0.05水平上差異顯著
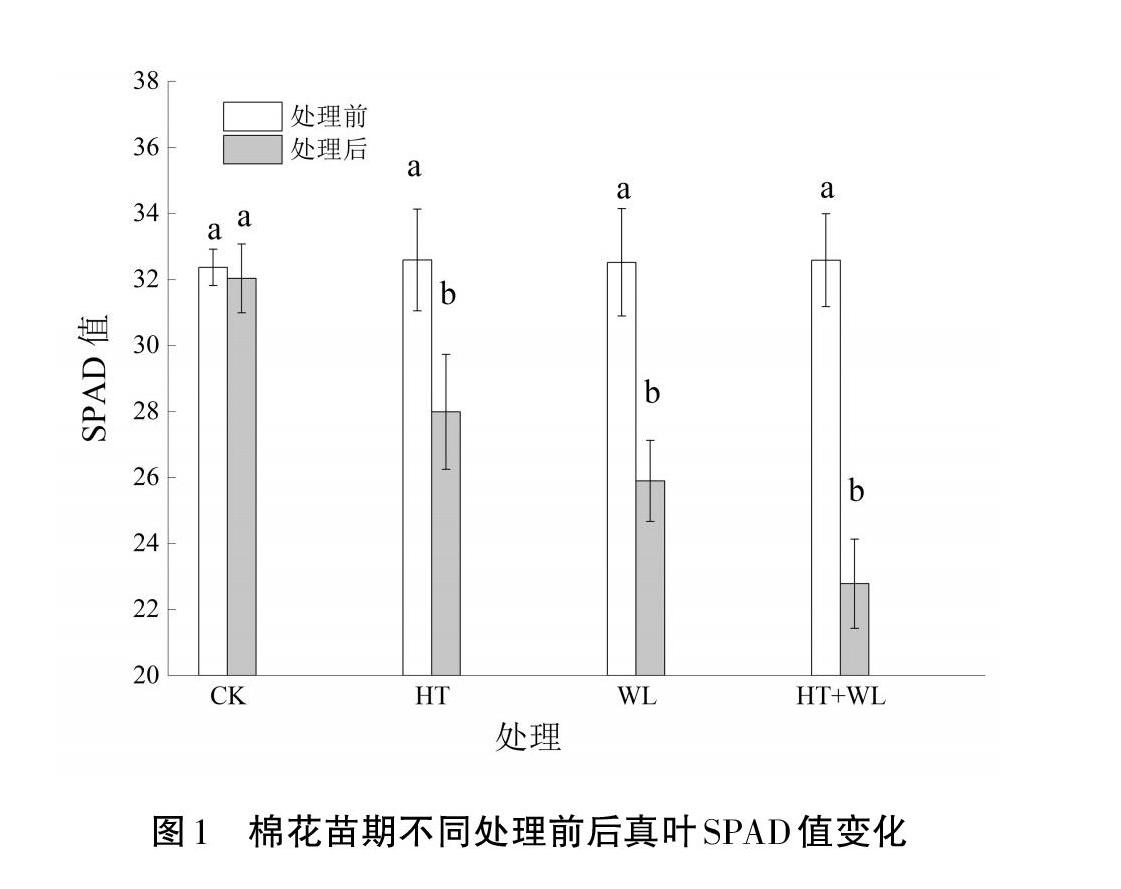
2.2? SPAD值(葉綠素含量)
從圖1可以看出,與處理前相比,HT、WL和HT+WL脅迫處理7 d棉花幼苗后,SPAD值分別下降了14.38%、20.69%、30.63%,HT條件處理后的降幅最小,WL居中,HT+WL條件處理后的降幅最大。其中HT+WL互作處理降幅較HT處理高6.35%、較WL處理高9.94%,WL處理較HT處理降幅高6.31%。說明三種脅迫處理均會造成SPAD值下降,引起棉花真葉光合速率下降。而HT+WL雙重脅迫加劇了SPAD值下降,單一HT或WL脅迫時,WL脅迫對SPAD值影響更大。
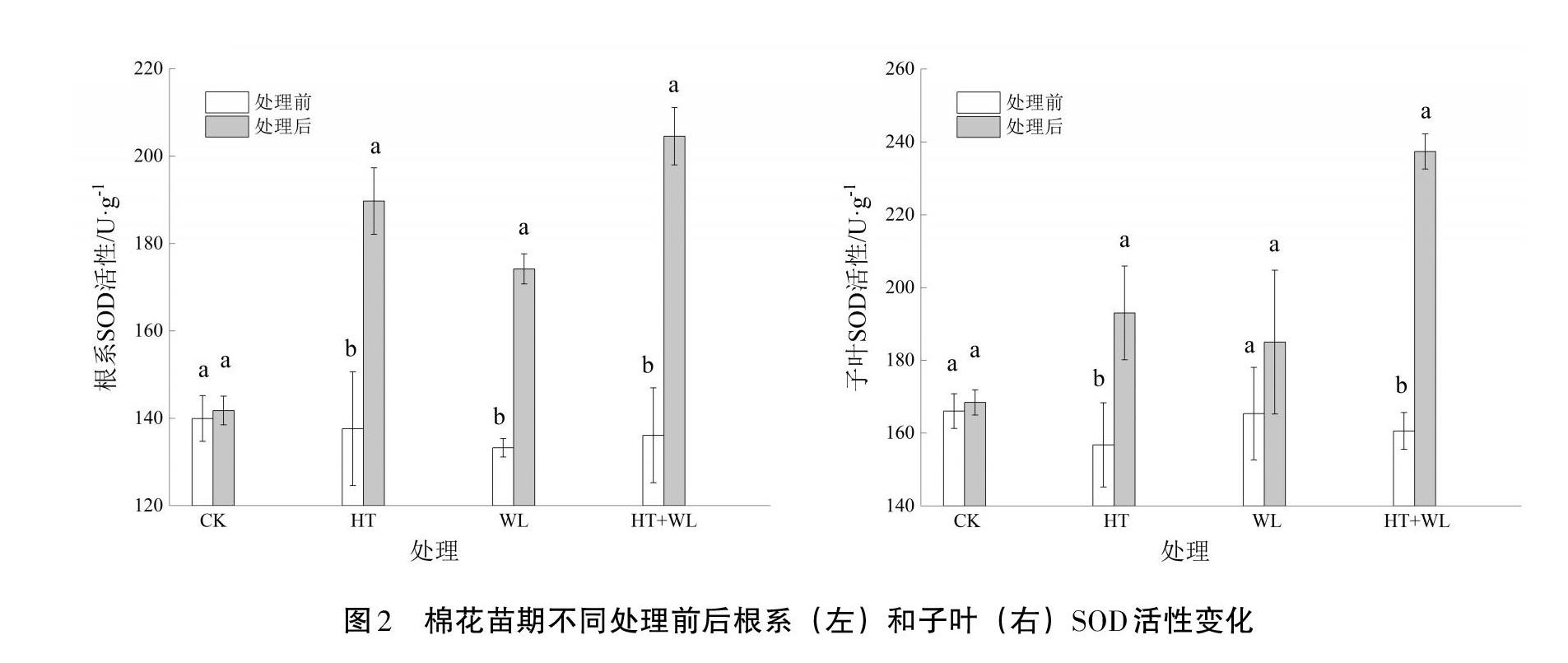
2.3? 超氧化物歧化酶(SOD)活性
從圖2可以看出,CK處理前后根和子葉SOD活性無明顯波動,而在HT、WL及HT+WL的脅迫下,棉花各個部位的SOD活性呈顯著增加,但在不同部位其活性增加的幅度有所不同。根SOD活性在HT、WL和HT+WL脅迫處理后較處理前分別增加了36.78%、28.89%和48.32%,HT+WL脅迫處理下根SOD活性增幅最大,HT脅迫處理居中,WL脅迫處理下根SOD增幅最小。單一脅迫7 d,HT處理比WL處理增幅高7.89%,可見HT脅迫比WL脅迫對根SOD活性的影響更大。棉花在遭受三種脅迫處理后與處理前相比,子葉SOD活性增加,但子葉SOD活性在HT、WL處理下變化較小;而HT+WL處理下子葉SOD活性大幅度增加,增幅高達45.71%。表明HT+WL互作對根SOD及子葉SOD活性的影響大于單一HT或WL處理。
2.4? 過氧化氫酶(CAT)活性
從圖3可以看出,四種處理的根CAT和子葉CAT活性在處理前基本一致,而經過處理后,CK較處理前的根CAT和子葉CAT活性增長不顯著,而HT、WL和HT+WL處理的根和子葉CAT活性較處理前出現了明顯增長。從圖3左圖看,HT、WL和HT+WL處理后的根CAT活性較處理前分別增長了14.8%、39.1%和63.3%;其中,HT+WL脅迫處理后的CAT活性增幅最高,較單一HT處理高48.5%,較單一WL處理高24.2%,其次是WL處理。從圖3右圖看,HT、WL、HT+WL處理后的子葉CAT活性分別較處理前增長了22.6%、76.3%、100.2%;其中,HT+WL條件下CAT活性增長了一倍,其次增長較快的是WL處理。在HT、WL和HT+WL脅迫下,CAT酶活性在不同部位增加幅度不同,子葉CAT活性增幅大于根CAT活性增幅,且單一WL處理影響高于單一HT處理,HT+WL雙重脅迫對CAT活性提升具有疊加效應。
2.5? 過氧化物酶(POD)活性
從圖4左圖可知,四種處理處置前的根POD活性基本一致,經過處理后,CK條件下處理后的棉花,根POD活性稍下降,而HT、WL、HT+WL處理的POD活性較處理前出現了一定程度的增長,增幅分別為13.47%、31.69%、56.67%;HT+WL處理后POD活性增長最高,增幅較HT處理高43.20%,較WL處理高14.98%。由圖4右圖可知,在CK、HT、WL和HT+WL條件下處理后的子葉POD活性較處理前分別增長了40.98%、36.23%、56.59%、88.16%,說明HT、WL和HT+WL這三種處理均會影響苗期棉花子葉的POD活性,而HT+WL脅迫的影響最明顯。由此說明在HT、WL和HT+WL的脅迫下,棉花各個部位的POD酶活性都呈顯著上升,在不同部位的POD酶活性增加幅度存在差異。而單一WL處理對POD活性的影響高于單一HT處理,HT+WL雙重脅迫對POD活性增長具有疊加效應。
2.6? 丙二醛(MDA)含量
從圖5左圖可知,根MDA含量在HT、WL和HT+WL處理后分別增加了56.83%、47.52%和85.75%,其中,根MDA的含量在HT+WL脅迫處理下增加幅度最大,HT脅迫處理居中,而WL脅迫處理下根MDA含量增加幅度最小,由此可知HT+WL脅迫處理對根MDA含量影響大于單一的HT或WL處理。由圖5右圖可知,四種處理前的子葉MDA含量基本一致,而經過處理后,MDA含量均較處理前呈現一定程度的增長,CK處理后變化不明顯,HT、WL、HT+WL處理后MDA含量分別較處理前增長了153.74%、150.41%、165.71%。在HT、WL和HT+WL的脅迫條件下,棉花幼苗各個部位的MDA含量都呈顯著增加,但在不同部位的MDA含量增加幅度不同,MDA的增幅在子葉中大于根。
3? 討論
3.1? 高溫與漬水對苗期棉花形態及光合特性的影響
當遭遇外界環境變化,從形態指標看,苗期棉花表現出較差的抗逆性。本研究發現對棉花的株高、根系和子葉中MDA含量及相對含水量,在HT、WL及HT+WL互作處理下均產生不同程度的抑制作用。WL抑制根的呼吸,以至于根部吸收運輸養分的效率下降,植株生長受到抑制,而HT脅迫使葉片的氣孔關閉,從而對光合作用的暗反應造成影響,有機物含量減少。此外,HT脅迫使植株水分吸收能力下降并因此受到一定程度的水分脅迫,此條件下葉片膜結構嚴重受損導致光合能力下降[19],同時植株含水量下降。當HT與WL同時發生時,株高和含水量的變化量不是單一HT或WL變化量之和,說明HT與WL處理存在互作效應。
葉片SPAD 值與葉綠素含量存在正比關系,是植物應對逆境脅迫的重要生理特征指標之一[20]。研究發現,HT、WL、HT+WL脅迫棉花幼苗7 d,SPAD值下降,且HT+WL雙重脅迫加劇了SPAD值減少,單一HT或WL脅迫時,WL脅迫對SPAD值影響要大一些,說明三種脅迫下棉花幼苗葉片葉綠素含量減少。表明HT條件下,增溫抑制了植株葉片的光合能力[21],葉綠素被氧化,抑制棉花葉片中的葉綠素合成,加速葉綠素分解[22-24],在WL脅迫下植物葉片氣孔導度降低,CO2通路擴散受到阻礙[25],從而光合色素的量降低,導致葉綠素的含量直線下降。這與吳啟俠等[26]的研究結果一致。因此,發生澇害影響后,可及時根據植株參數變化,做好相應補救措施,比如噴灑適量調節劑,施鉀肥促進光合作用[27]。
3.2? 高溫與漬水對棉株抗氧化保護的影響
棉花在HT等逆境條件生長過程中,植株體內會堆積形成大量的活性氧(ROS),特別是超氧化物自由基(O2?)和過氧化氫(H2O2),使膜脂過氧化增加[28],并積累有害物質膜脂過氧化物丙二醛(MDA)[29],傷害膜體系,引起細胞功能失常[30]。植株的內源保護酶(SOD、CAT、POD)活性為應對積累的ROS而有所提高,從而減輕其對體內的毒害[31-32]。本研究結果表明,無論是HT、WL還是HT+WL互作處理,都會使棉花子葉和根部的MDA含量升高,這與前人的研究結果一致[33-34]。在此基礎上,研究發現HT+WL互作處理對根部MDA具有明顯的疊加效應,同時,子葉MDA含量增幅大于根,但雙重脅迫與單一脅迫對MDA含量的影響程度相差無幾。這些脅迫因子均會導致棉花幼苗細胞發生膜脂氧化損傷,且子葉細胞膜傷害程度比根嚴重,雙重脅迫對根的影響更為直接。在本研究中,HT或WL處理棉花幼苗7 d后,棉花各部位SOD、CAT、POD活性較處理前升高,這是對棉花幼苗中H2O2和O2?積累的響應[11]。說明中棉所45苗期植株為適應逆境脅迫,能夠進行生理適應性滲透調節——細胞主動積累調節物質[35],提高SOD、CAT、POD活性,以削減植物體內因HT或WL脅迫產生的自由基和活性氧,保護植物免受氧化損傷,同時利于棉花幼苗適應漬水、高溫等非生物脅迫。
迄今為止,無論是傳統育種還是分子育種方法,對棉花耐澇耐高溫品種的選育仍在不斷探索。有研究針對澇害脅迫對棉花植株帶來的傷害常采用生長調節劑對作物進行營養調控和化學調控,使之對棉花產量的影響降至最低[27]。本試驗可以進一步改善的是,應該在量化了HT、WL單一脅迫及雙重脅迫對內源保護酶的影響后,進一步培養植株,對部分脅迫處理后的植株進行不同生長調節劑處理,觀察后續產量是否與正常生長植株相去甚遠,添加多少劑量提高最多。今后的研究應傾向于利用已知生理機制解決一些不可避免的多重脅迫帶來的傷害,以及深入研究分子調控水平問題,以期為實際農業生產發展提供有益指導。
4? 結論
本試驗結果表明:1)HT、WL或HT+WL處理下苗期棉花株高較CK增速慢,真葉含水量較CK含量減少,葉片SPAD值較處理前顯著降低。與CK相比,HT、WL和HT+WL處理下棉花株高分別低0.91%、9.36%、11.36%;棉花真葉含水量降低20.46%、19.81%、24.95%;葉片SPAD值較處理前分別下降14.38%、20.69%、30.63%。2)根和子葉的SOD、POD、CAT活性和MDA含量有所增加。在HT、WL和HT+WL脅迫處理后,根SOD活性較處理前分別增加了36.78%、28.89%和48.32%,而子葉SOD則分別增長了15.00%、9.84%和40.93%;根CAT活性較處理前分別增長了14.8%、39.1%和63.3%,子葉CAT活性分別增長了22.6%、76.3%、100.2%;根POD 較處理前分別增長了 13.47%、31.69%、56.67%,子葉POD活性分別增長了36.23%、56.59%、88.16%;根MDA含量較處理前分別增加了56.83%、47.52%、85.75%,子葉MDA含量較處理前分別增長了153.74%、150.41%、165.71%。3)綜上,HT和WL雙重脅迫對中棉所45棉花幼苗的內源保護酶活性變化有疊加效應。該結果可為棉花抗逆品種選育和大田雨熱同期災后幼苗栽培管理提供理論指導。
參考文獻:
[1] 張杰,王力,趙新民.我國棉花產業的困境與出路[J].農業經濟問題,2014,35(9):28-34,110.
[2] 郭進宇,臧學斌,周仙梅.雨澇災害對棉花生長發育的影響及減災措施[J].江西棉花,2011,33(5):48-49.
[3] 錢龍,王修貴,羅文兵,等.澇漬脅迫對棉花形態與產量的影響[J].農業機械學報,2015,46(10):136-143,166.
[4] Kuai Jie,Zhou Zhiguo,Wang Youhua. The effects of short-term waterlogging on the lint yield and yield componentsof cotton with respect to boll position[J]. European Journal of Agronomy,2015,67(3):29-36.
[5] Li T,Suk-Yoon K,Sun-Hyung K,et al. Enhanced tolerance of transgenic potato plants expressing both superoxide dismutase and ascorbate peroxidase in chloroplasts against oxidative stress and high temperature[J]. Plant Cell Reports,2007,19(3):227-239.
[6] Suzuki N,Koussevitzky S,Mittle R,et al. ROS and redox signalling in the response of plants to abiotic stress[J]. Plant,Cell and Environment,2012,35(2) :259-270.
[7] Wang Haimiao,Chen Yinglong,Hu Wei,et al. Short-term soil-waterlogging contributes to cotton cross tolerance to chronic elevated temperature by regulating ROS metabolism in the subtending leaf[J]. Plant Physiology and Biochemistry,2019,139:333-341.
[8] 劉凱文,蘇榮瑞,朱建強,等.棉花苗期葉片關鍵生理指標對澇漬脅迫的響應[J].中國農業氣象,2012,33(3):442-447.
[9] 張寄陽,劉祖貴,段愛旺,等.棉花對水分脅迫及復水的生理生態響應[J].棉花報,2006(6):398-399.
[10] 王曉曼,王瓊珊,夏松波,等.高溫脅迫下棉花苗期的生理響應和轉錄組分析[J].湖北農業科學,2019,58(S2):423-428.
[11] 武輝,張巨松,石俊毅,等.棉花幼苗對不同程度低溫逆境的生理響應[J].西北植物學報,2013,33(1):74-82.
[12] 武輝.低溫脅迫下棉花幼苗生理生化響應機制的研究[D].烏魯木齊:新疆農業大學,2012.
[13] 馮棣,張俊鵬,孫池濤,等.不同生育階段鹽分脅迫對棉花生長和水分生理指標的影響[J].生態學雜志,2014,33(5):1195-1199.
[14] 張雷,張國偉,孟亞利,等.鹽分條件下棉花相關生理特性的變化及水分脅迫指數模型的構建[J].中國農業科學,2013,46(18):3768-3775.
[15] 吳曉東,王巍,金路路,等.鹽脅迫對棉花光合作用和生理指標的影響[J].中國棉花,2013,40(6):24-26.
[16] 蘇仙絨,馬沛勤,陳莉.銅、鎘脅迫對棉花幼苗抗氧化生理指標的影響[J].運城學院學報,2014,32(2):66-69.
[17] 張曉紅,胡根海,任曉亞,等.汞脅迫對棉花幼苗生理特性的影響[J].揚州大學學報(農業與生命科學版),2020,41(5):24-28.
[18] 李合生.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000.
[19] 孫嘯震,張黎妮,戴艷嬌,等.花鈴期增溫對棉花干物重累積的影響及其生理機制[J].作物學報,2012,38(4):683-690.
[20] 楊威,朱建強,吳啟俠,等.花鈴期短期漬水和高溫對棉花葉片光合特性、膜脂過氧化代謝及產量的影響[J].棉花學報,2016,28(5):504-512.
[21] 賀新穎,周治國,戴艷嬌,等.鈴期增溫對棉花產量、品質的影響及其生理機制[J].應用生態學報,2013,24(12):3501-3507.
[22] 云建英,楊甲定,趙哈林.干旱和高溫對植物光合作用的影響機制研究進展[J].西北植物學報,2006(3):641-648.
[23] 田禮欣.澇漬脅迫對玉米農藝性狀、生理特性及產量的影響[D].哈爾濱:東北農業大學,2019.
[24] 陳少裕.膜脂過氧化對植物細胞的傷害[J].植物生理學通訊,1991(2):84-90.
[25] 艾天成,李方敏,周治安,等.作物葉片葉綠素含量與SPAD值相關性研究[J].湖北農學院學報,2000,20(1):6-8.
[26] 吳啟俠,朱建強,楊威,等.花鈴期高溫受澇對棉花的交互效應及排水指標確定[J].農業工程學報,2015,31(13):98-104.
[27] 王曾楨.植物生長調節劑與營養調控減緩棉花澇后危害研究[D].荊州:長江大學,2014.
[28] Toai V,T. T.,Bolles,C. S. Postanoxic injury in soybean (Glycine max) seedlings[J]. Plant Physiology,1991,7:588-592.
[29] 李建英,周長軍,楊柳,等.水分脅迫對大豆苗期葉片內源激素含量與保護酶活性的影響[J].大豆科學,2010,29(6):959-963..
[30] 王小菁.植物生理學(第8版)[M].北京:高等教育出版社,2019.
[31] 何學利.植物體內的保護酶系統[J].現代農業科技,2010(10):37-38.
[32] 莫紅,翟興禮.干旱脅迫對大豆苗期生理生化特性的影響[J].湖北農業科學,2007,46(1):45-48.
[33] 鄧茳明,熊格生,袁小玲,等.棉花不同耐高溫品系的SOD、POD、CAT活性和MDA含量差異及其對盛花期高溫脅迫的響應[J].棉花學報,2010,22(3):242-247.
[34] 陳娟,梁明霞,潘開文.澇漬脅迫下生姜幼苗生長及體內保護酶活性變化[J].江蘇農業科學,2015,43(5):152-155.
[35] 徐道青,鄭曙峰,王維,等.棉花澇害脅迫研究綜述[J].中國農學通報,2014,30(27):1-4.
(責任編輯:丁志祥)
