意大利蒼耳與烏拉爾甘草種間競爭能力比較
2021-10-09 01:19:14徐懷秀
生態學報 2021年16期
關鍵詞:植物
徐懷秀,馬 淼
新疆植物藥資源利用教育部重點實驗室,石河子大學生命科學學院, 石河子 832003
生物入侵是當前世界性的生態環境問題[1],隨著全球化進程加快,入侵物種嚴重威脅到了各國生態系統的完整性[2]。其中農田生態系統中植物群落結構簡單,穩定性較低,而入侵植物大多具有強大的競爭力,這使得農田生態系統往往最易受到外來植物的成功入侵[3]。一些與作物具有相似生態位的入侵植物,會通過種間競爭來獲得生存優勢,并對作物的生長、繁殖和產量積累產生強烈的負面影響[4- 5]。
意大利蒼耳(XanthiumitalicumMoretti)為菊科(Compositae)蒼耳屬(Xanthium)一年生草本植物,原產于北美洲,現廣泛分布于東、西半球中緯度地區,已成為世界性入侵雜草[6]。1991年在我國北京首次發現,目前已經廣泛分布至遼寧、北京、山東、河北、陜西、新疆等地,并且有擴大蔓延至全國各地的趨勢[7- 8]。其中意大利蒼耳在新疆的危害最為嚴重,目前在新疆已擴散至伊犁、博州、阿勒泰、塔城、石河子、昌吉、烏魯木齊等地區,已占據了農田、草原、林緣、溝渠及水庫淺水區等多種生境[9]。對新疆的農業生產和畜牧業發展都產生了極大危害[10- 11]。
烏拉爾甘草(GlycyrrhizauralensisFisch)為豆科(Leguminosae)甘草屬(Glycyrrhiza)多年生草本植物,其根及根狀莖是西北地區的特色道地性藥材[12],其中含有的甘草酸和甘草苷等黃酮類活性成分,具有抗病毒、抗癌、抗過敏、抗炎等功效[13]。同時,烏拉爾甘草的莖葉部分也是優良的飼草,被廣泛應用于畜牧業[14]。極高的藥用價值及經濟價值使其成為中國西北地區大面積種植的一種經濟作物。本課題組前期在新疆的野外調查中發現,意大利蒼耳已經開始向烏拉爾甘草農田擴張,與烏拉爾甘草形成混生的格局。研究表明,外來入侵植物進入新生境后會與本地植物通過搶占空余生態位或通過種間競爭獲得優勢[15- 16],其中種間競爭是入侵植物排擠本地種的一種重要策略,其主要是基于資源的競爭性利用[5]。影響植物種間競爭結果的因素包括種間競爭能力的遺傳差異,資源的有效性,生態位相似性,共生微生物,空間分布格局,競爭期長短等[17]。入侵植物意大利蒼耳具有較高的生長速率,易于形成高密度單優勢種群落,而農田生境通常具有較好的水和營養資源,這使得農田中意大利蒼耳的株高和冠幅能夠分別達到2 m和1.6 m[9, 18],高大的個體和龐大的冠幅使意大利蒼耳對農田作物產生嚴重的遮蔽,從而造成農田作物的嚴重減產[4]。在烏拉爾甘草農田中,甘草的株高通常不到1 m,在與意大利蒼耳混生的群落中位于意大利蒼耳的下層,這可能使得意大利蒼耳在對光資源的競爭中占有優勢地位。此外,意大利蒼耳旺盛的生長,勢必會加大對土壤中水分和礦質營養的獲取,而這也可能會加劇其與農田中的甘草之間對于資源的競爭。以往的研究更多關注的是意大利蒼耳對入侵地自然生態系統所造成的影響[6,8,10, 19],而其對農田的入侵風險及后果的評估研究較少,其與甘草屬植物之間的競爭關系也不明確,關于入侵植物意大利蒼耳與烏拉爾甘草的種間競爭會對烏拉爾甘草的生長及品質會產生怎樣的影響尚不清楚。因此,本研究采用取代試驗法,在受控條件下,模擬栽培烏拉爾甘草常規水肥管理條件,開展了意大利蒼耳與烏拉爾甘草種間競爭關系及競爭結果的研究,以期為意大利蒼耳對烏拉爾甘草農田的入侵能力及入侵后果的評估提供試驗依據。
1 材料與方法
1.1 試驗區概況
試驗地點位于新疆石河子市石河子大學(44°31′10″N, 86°06′94″E),海拔約450 m,屬典型的大陸性干旱氣候,光照充足,降水少蒸發大。年均降水量180 mm左右,年均蒸發量1820 mm左右,年均氣溫為7.2℃,年日照時數2721—2818 h,無霜期約147 d。
1.2 試驗材料
試驗所用的意大利蒼耳成熟種子于2018年10月采自新疆石河子市東郊(44°30′01″N, 86°12′35″E);烏拉爾甘草的成熟種子于2018年9月采自新疆尉犁縣(41°38′83″N, 86°28′73″E),種子自然陰干后置于4℃保存。
1.3 試驗方法
1.3.1種子預處理
為保證種子萌發的同步性,選取大小一致、成熟飽滿的意大利蒼耳種子,播種前在室溫下置于蒸餾水中浸泡12 h;選取大小一致、成熟飽滿的烏拉爾甘草種子,用98%的硫酸溶液浸泡約0.5 h,用流水沖洗至種子表面無硫酸殘留,在室溫下用蒸餾水浸泡12 h。
1.3.2試驗設計
于2019年4月15日將經過水浸泡處理的意大利蒼耳和烏拉爾甘草種子播種于育苗盆中,以沙土作為栽培基質,播種深度為1 cm,置于戶外開闊地,每天用稱重法補水以保持土壤相對濕度為70%左右,待幼苗長出4枚真葉時選長勢一致的幼苗并將其移栽于直徑為22 cm,高度為20 cm的塑料花盆中。根據甘草農田高產管理栽植技術[20],其栽植密度通常為120萬株/ hm2,與盆栽面積進行換算,確定本研究中總種植密度為4株/盆。采用取代試驗法[17, 21],設置單種種植和混種種植兩種模式,共計三種處理,即單種種植意大利蒼耳,單種種植烏拉爾甘草以及意大利蒼耳與烏拉爾甘草混合種植,單種種植指每盆單獨種植4株意大利蒼耳(即意大利蒼耳∶烏拉爾甘草=4∶0)或每盆單獨種植4株烏拉爾甘草(即意大利蒼耳:烏拉爾甘草=0:4),混種種植時每盆種植意大利蒼耳和烏拉爾甘草各2株(即意大利蒼耳:烏拉爾甘草=2:2),四株個體在花盆中呈“十”字形均勻分布,兩種植物相同排列,具體種植方式見圖1。
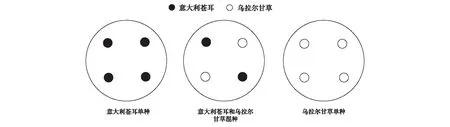
圖1 意大利蒼耳和烏拉爾甘草栽植示意圖Fig.1 The planting diagram of Xanthium italicum and Glycyrrhiza uralensis
栽培基質為沙土(全氮0.268 g/kg、全磷0.0855 g/kg、全鉀5.72 g/kg、速效氮43.59 mg/kg、速效磷4.1 mg/kg、速效鉀119.09 mg/kg、有機質5.81 g/kg)。使用稱重法,每天早晚補水,保持70%的土壤相對濕度;模擬甘草農田土壤肥力水平[22]分2次(2019年5月21日和2019年5月28日)對各個花盆進行施肥(1.4492 g尿素、1.9354 g過磷酸鈣、1.7108 g硫酸鉀)。每種處理重復6次,計18盆。所有花盆都隨機擺放在開闊地,每隔10 d隨機調換一次各花盆的位置,以消除光照和邊際效應的影響。于2019年9月1日意大利蒼耳生育期結束(即果實成熟)后,收獲全部植株。
1.4 研究指標及方法
1.4.1形態學參數的測定
植株收獲后,用水將植株的根部沖洗至無浮土,測量所有植株的株高、冠幅及烏拉爾甘草的根狀莖長度;統計意大利蒼耳的種子數量和烏拉爾甘草的根狀莖條數與根瘤菌數量;用根系掃描儀(WinRHIZO La 2400)測量烏拉爾甘草的根總長度、根總表面積及根平均直徑。
1.4.2生物量參數的測定
將每株植株的根、莖、葉、果實(意大利蒼耳)、根狀莖(烏拉爾甘草)進行分離,置于60℃烘箱中烘干至恒重,稱量各部分生物量并計算總生物量。
1.4.3甘草酸含量的測定
將烘干的烏拉爾甘草的根及根狀莖混合粉碎成粉末,過80目篩。分別精確稱取單種處理組和混種處理組的粉末各0.50 g,置于10 mL離心管中,加入提取液5 mL(甲醇: 水: 36%冰醋酸= 71: 28: 1),用超聲波提取儀超聲提取30 min,用高速離心機(8000 r/min)離心10 min,吸取上清液,沉淀物再加入5 mL提取液,超聲提取30 min,用高速離心機(8000 r/min)離心10 min,吸取上清液,并與第一次的上清液混勻,上清液經0.45 μm微孔濾膜過濾于安捷倫進樣瓶中。使用高效液相色譜儀(Agilent 1200; Agilent Technologies, CA, USA)測量甘草地下器官(根及根狀莖)的甘草酸含量[23]。
1.4.4競爭關系
分別計算這兩種植物的相對產量(Relative Yield,RY)[21]、相對競爭強度(Relative Competition Intensity, RCI)[24]以及競爭攻擊系數(Aggressivity,A)[25],以確定兩物種間的競爭關系,其公式為:
RYa=Yab/(Zab×Yaa) 或RYb=Yba/(Zba×Ybb)
RCIa=(Yaa-Yab)/Yaa或 RCIb=(Ybb-Yba)/Ybb
Aa=Yab/(Yaa×Zab)-Yba/(Ybb×Zba) 或Ab=Yba/(Ybb×Zba)-Yab/(Yaa×Zab)
式中,a、b分別代表意大利蒼耳和烏拉爾甘草,Yab代表混合種植處理組中意大利蒼耳的單株生物量,Yaa代表單獨種植時意大利蒼耳的生物量,Yba代表混合種植處理組中烏拉爾甘草的單株生物量,Ybb代表單獨種植時烏拉爾甘草的生物量,Zab為混合種植體系中意大利蒼耳所占的比例,Zba為混合種植體系中烏拉爾甘草所占的比例。
當RY<1時,表示種間競爭大于種內競爭;RY> 1時,表示種內競爭大于種間競爭;RY=1時,表示種間競爭等于種內競爭。
當RCI值位于0—1之間時,RCI值越大,說明該物種在混種環境中產量越低,另一植物對它的影響越大,且該物種在混種群落中的競爭能力越差;當RCI=1時說明該物種已被排斥出去;當RCI=0時,說明該物種對混種的其他物種在產量上沒有影響。當RCI<0時,表明一種植物的存在能夠促進該植物的生長,促進其產量。
當Aa> 0時,表明物種a的競爭能力大于物種b;當Aa= 0時,表明物種a與物種b的競爭力相同,當Aa< 0時,表明物種a的競爭能力小于物種b。
1.5 數據處理
使用SPSS 19.0軟件進行數據的統計分析,用單因素方差分析(one-way ANOVA)和非參數檢驗(nonparametric tests)分析每種植物在單種處理和混種處理下的形態學性狀、生物量及甘草的地下器官甘草酸含量的差異顯著性,使用T檢驗(T-Test)比較RY與1,以及RCI、A與0之間的差異顯著性,使用origin8. 5軟件進行作圖。
2 結果
2.1 種植模式對意大利蒼耳和烏拉爾甘草生長的影響
種植模式對意大利蒼耳的株高、冠幅及種子數量均具有極顯著影響(P<0.01)。混種處理下意大利蒼耳的株高、冠幅及種子數量相比單種處理分別增加了13%、27%和56%(圖2)。
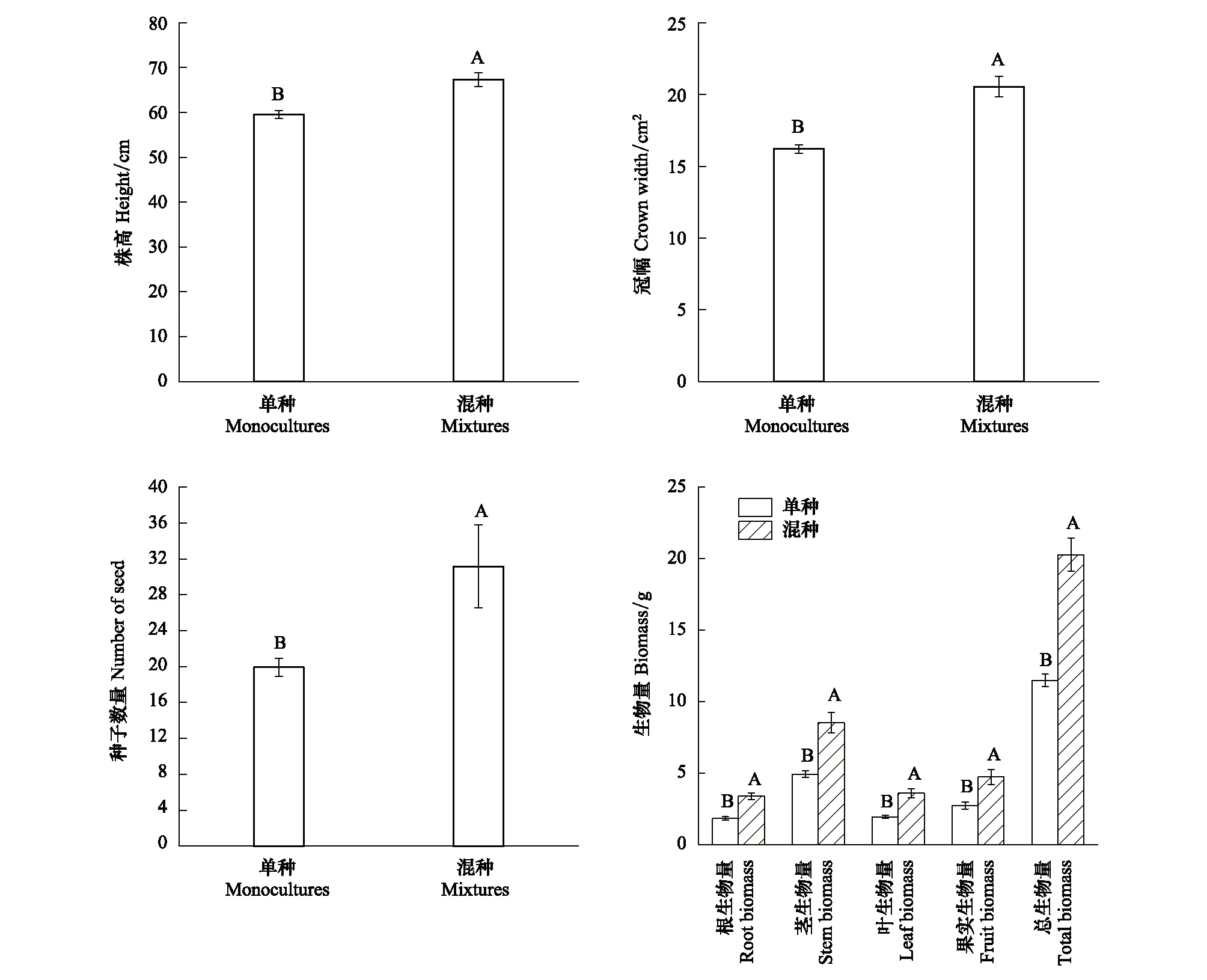
圖2 種植模式對意大利蒼耳生長和生物量積累的影響(均值±標準誤) Fig.2 Effects of planting pattern on growth and biomass accumulation of Xanthium italicum (mean±SE)不同大寫字母表示單種與混種處理在1%水平上差異顯著
種植模式也極顯著地影響了烏拉爾甘草的株高、冠幅、根總長度、根總表面積、根平均直徑、根狀莖條數、根狀莖長度和根瘤數(P<0.01)(表1)。混種處理組中烏拉爾甘草的株高和冠幅相比單種處理組分別下降了35%和45%。單種處理下單株烏拉爾甘草的平均根狀莖條數為3條,根狀莖總長度平均為1.6 cm,而混種處理組中的烏拉爾甘草均未生長出根狀莖。混種處理組中烏拉爾甘草的根總長度、根總表面積、根平均直徑及根瘤數相比單種處理分別下降了55%、63%、19%和76%。
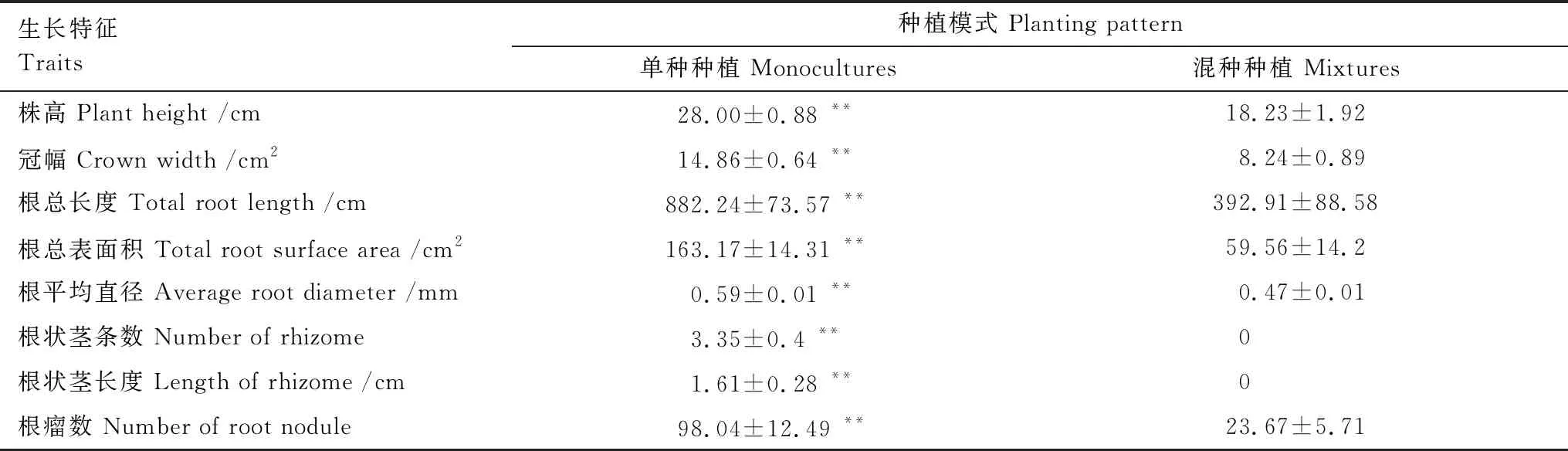
表1 種植模式對烏拉爾甘草的生長的影響(均值±標準誤)
2.2 種植模式對意大利蒼耳和烏拉爾甘草生物量分配的影響
種植模式對意大利蒼耳的根、莖、葉、果實和總生物量均具有極顯著影響(P<0.01)。意大利蒼耳的根、莖、葉、果實及總生物量在混種處理下相比單種處理,分別增長了84%、73%、84%、73%和77%(圖2)。
種植模式也顯著影響了烏拉爾甘草的根狀莖生物量(P=0.014),極顯著地影響其根、莖、葉及總生物量的積累(P<0.01)。單種條件下烏拉爾甘草的根狀莖生物量為0.016 g,而混種處理下烏拉爾甘草未生長出根狀莖,其根、莖、葉及總生物量相比單種處理分別下降了72%、80%、65%和71%(圖3)。
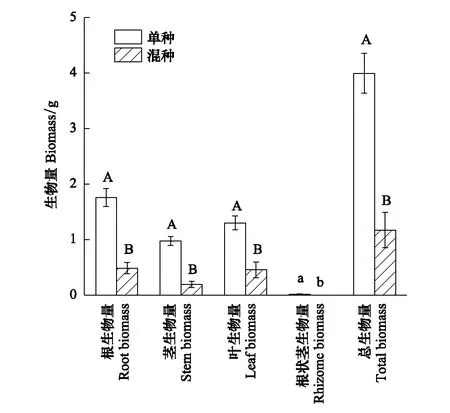
圖3 種植模式對烏拉爾甘草生物量的影響(均值±標準誤) Fig.3 Effects of planting pattern on the biomass of Glycyrrhiza uralensis (mean±SE)不同大寫字母表示單種與混種處理在1%水平上差異顯著,不同小寫字母表示單種與混種處理在5%水平上差異顯著
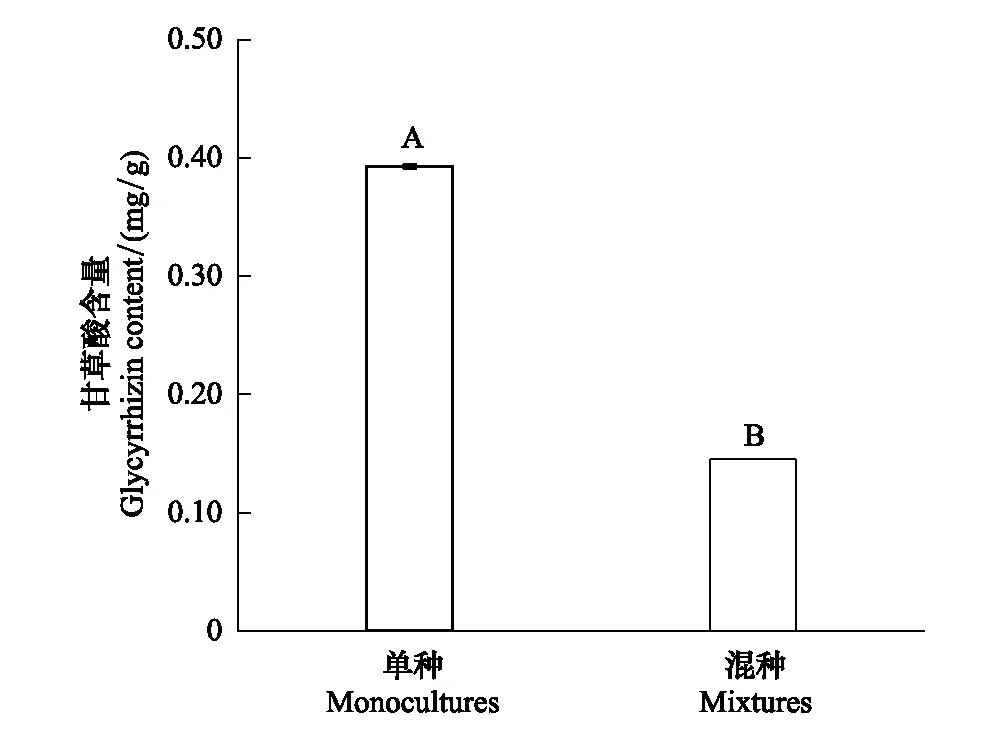
圖4 種植模式對烏拉爾甘草地下器官甘草酸含量的影響(均值±標準誤)Fig.4 Effects of planting pattern on glycyrrhizic acid content in underground organ of Glycyrrhiza uralensis (mean±SE)
2.3 種植模式對烏拉爾甘草甘草酸含量的影響
種植模式極顯著地影響了烏拉爾甘草地下器官甘草酸含量的積累(P<0.01)。混種處理組中烏拉爾甘草的甘草酸含量比單種處理組下降了63%(圖4)。
2.4 意大利蒼耳和烏拉爾甘草的競爭關系
意大利蒼耳的相對產量(RYa)大于1,表明意大利蒼耳的種內競爭大于來自烏拉爾甘草的種間競爭;其相對競爭強度(RCIa)小于0,表明與烏拉爾甘草混種會有利于意大利蒼耳生物量的積累;其競爭攻擊系數(Aa)大于0,表明混種的意大利蒼耳的競爭能力大于烏拉爾甘草(圖5)。
烏拉爾甘草的相對產量(RYb)小于1,表明烏拉爾甘草的種內競爭小于來自意大利蒼耳的種間競爭;其相對競爭強度(RCIb)在0—1之間,表明意大利蒼耳對烏拉爾甘草的影響較大且烏拉爾甘草在混種群落中競爭能力較弱;其競爭攻擊系數(Ab)小于0,表明混種的烏拉爾甘草競爭能力小于意大利蒼耳(圖5)。
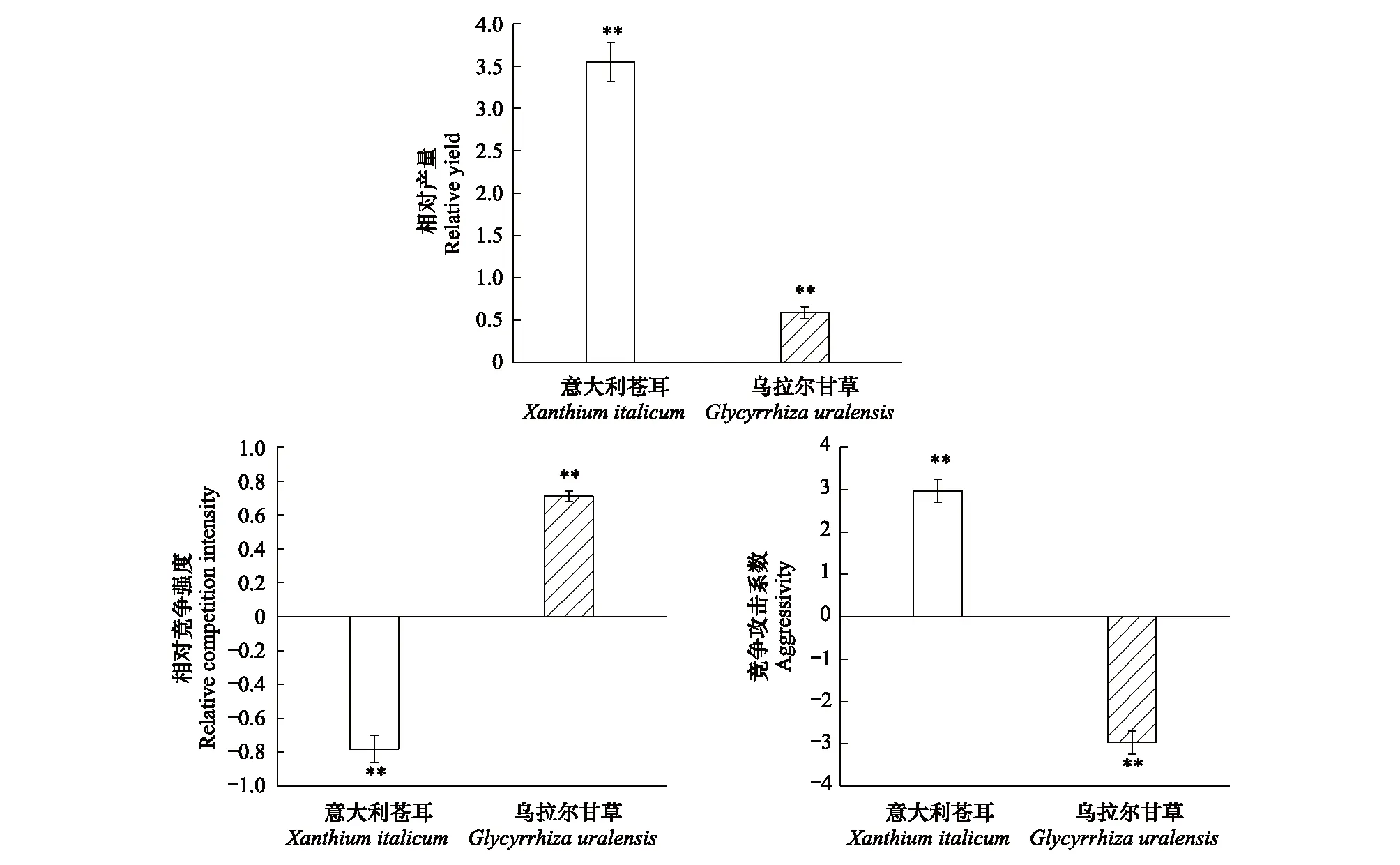
圖5 不同種植模式下該兩種植物的競爭關系(均值±標準誤)Fig.5 Competitive relationship of the two species based on different planting pattern (means±SE) **表示競爭指標與假設檢驗值之間(即RY與1比較,RCI和A分別與0比較) 存在極顯著差異,(單一樣本T檢驗:P<0.01, n=12)
3 討論
外來植物與本地植物之間的競爭關系是入侵生態學的重要研究內容之一,外來植物能否成功入侵,主要取決于該植物自身的入侵力與生境的可入侵性[26]。而該植物自身入侵力主要取決于物種本身的生物學特性,其中就包括該植物的種間競爭能力[27- 28]。不同的物種具有不同的競爭策略,而植株的大小特征是預測植物相對競爭力的最佳指標,生物量是其中最重要的參數[29]。在本研究中,與單種相比,混種能夠顯著提高意大利蒼耳根、莖、葉及果實的生物量,這意味著混種處理會有利于該入侵植物的生物量積累,從而提高其相對競爭力[30- 31]。我們發現與單種模式相比,在混種處理中意大利蒼耳根和葉的生物量增加幅度最大,這意味著存在競爭作用時,意大利蒼耳的生物量分配會趨向于增加根和葉器官的生長,發達的根系使其能更高效地吸收土壤中的水分和無機營養,而發達的葉片則意味著更大的光合面積,這有助于其制造出更多的光合產物,使其具有更大的營養器官,從而提高其自身的入侵力[32]。混種處理中意大利蒼耳的莖生物量也顯著增加,在與烏拉爾甘草混生時,意大利蒼耳能夠生長出更高的個體和更大的冠幅,使其在地上空間占領方面具有更明顯的優勢,造成物種間競爭的不平衡[33],進而有利于該入侵植物對甘草農田的成功入侵。研究表明,意大利蒼耳的種子具有良好的越冬性能,其在各種極端條件下越冬依舊能夠保持較高的萌發率[11]。本研究中,混種使意大利蒼耳的種子數量顯著增加,這意味著入侵甘草地后,意大利蒼耳能夠產生更多的種子,良好的越冬性能及數量巨大的種子使其有性繁殖能力得到顯著提高,于是在下一個生長季節,被意大利蒼耳入侵的烏拉爾甘草農田中將會出現更多的意大利蒼耳個體,而這也將加劇烏拉爾甘草遭受到的意大利蒼耳的競爭抑制威脅。
生境的可入侵性主要取決于非生物環境和生物環境對入侵植物的抵抗力[34],農田生境作為一種高強度的人為擾動環境,其資源可用量會受人為擾動發生改變[35],而這種高強度的擾動通常都利于外來物種的成功入侵[36]。因此,農田生境的可入侵性,則主要取決于生物環境對入侵植物的抵抗力,其中作物與入侵植物的種間競爭就是一種重要的抵抗策略[37]。在本研究中,混種處理中的烏拉爾甘草受到意大利蒼耳強烈的競爭抑制,各部分生物量的積累均顯著降低。相對于單種處理,混種處理中烏拉爾甘草的株高、冠幅及根系的生長參數均呈現大幅的下降,這些營養器官的生長受到顯著抑制,必然會對烏拉爾甘草葉片的光捕獲能力和根系對土壤水分及營養資源的獲取能力產生極其不利的影響。這意味著在該兩種植物對主要資源的競爭利用中,烏拉爾甘草處于不利的地位,對意大利蒼耳的入侵表現出較弱的抵抗力。同時意大利蒼耳還強烈地抑制了甘草根狀莖的發育,這使得烏拉爾甘草的克隆繁殖潛力被顯著降低。而甘草酸是中藥材甘草的主要活性成分[38],藥用植物有效成分的含量會受伴生植物種類的強烈影響[39]。本研究結果顯示,混種處理組中烏拉爾甘草地下器官甘草酸含量極顯著地低于單種處理組的烏拉爾甘草,這表明意大利蒼耳能夠顯著降低中藥材甘草的品質。綜合來看,與意大利蒼耳混生情況下,烏拉爾甘草的個體生長、克隆繁殖能力以及藥材甘草的產量與品質均受到強烈的抑制。
共生微生物是影響植物競爭的一種重要因素之一[17]。根瘤菌可以與豆科植物的根形成共生體系,這種共生體系是生物固氮中最重要的一種類型[40]。本研究中發現,與單種相比,混種處理組中烏拉爾甘草根系的根瘤數量明顯降低,其數量僅是單種處理組的四分之一左右,這表明與意大利蒼耳的混種處理會強烈抑制根瘤菌與烏拉爾甘草根共生體系的形成。研究表明,混生情況下根瘤菌的缺失對豆科植物的影響明顯大于其他植物[41],但本研究中甘草根瘤數量減少的內在機制尚不明確,需進行更深入的研究。但從長遠來看,這種根瘤發育的受阻和根瘤數量的減少,勢必會對農田土壤的氮素水平產生負面的影響。
植物的相對產量、相對競爭強度以及競爭攻擊系數是解釋物種間競爭關系的重要指標[42]。在本研究中,相對產量的結果表明,意大利蒼耳受到鄰株同種其他個體的種內競爭壓力大于來自鄰株烏拉爾甘草的種間競爭壓力,而烏拉爾甘草受到鄰株同種其他個體的種內競爭壓力則小于來自鄰株意大利蒼耳的種間競爭壓力。相對競爭強度結果顯示,混種模式對意大利蒼耳的生長有顯著的促進作用,這可能是由于鄰株的烏拉爾甘草具有良好的固氮作用[40],因此與意大利蒼耳單種模式相比,混種模式可能具有更高的土壤氮素水平,而入侵植物通常能夠比本地植物具有更高的氮資源利用能力[43],從而可能影響到兩種植物間的競爭關系。競爭攻擊力系數的數據表明,意大利蒼耳的競爭能力大于烏拉爾甘草,在二者混生的群落中,意大利蒼耳將會在競爭中占據優勢地位。
綜上所述,在與烏拉爾甘草混合種植時,意大利蒼耳的營養生長和生殖生長均得到了極顯著的促進,使其個體比單種處理組長得更高大,并能產生更多的種子。這不僅顯著地抑制了烏拉爾甘草的生長,而且強烈削弱了烏拉爾甘草憑借根狀莖的克隆繁殖能力,并使烏拉爾甘草的藥材產量顯著降低,同時還強烈抑制了甘草酸的形成和積累,從而消弱了甘草藥材的品質。該藥材產量和品質的大幅下降必然會給農戶造成巨大的經濟損失。甘草莖和葉生物量的顯著降低也勢必造成家畜飼草產量的下降,從而危及當地畜牧業的健康發展。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
