艾美爾球蟲對高原鼠兔個性與生理特征的影響
2021-09-27 01:38:40朱紅娟余義博曲家鵬
草業科學 2021年8期
鐘 亮,朱紅娟,余義博,曲家鵬
(1.中國科學院西北高原生物研究所,青海 西寧 810008;2.中國科學院大學,北京 100049;3.青海省動物生態基因組學重點實驗室,青海西寧 810008)
寄生是自然界中普遍存在的一種行為[1-3]。寄生蟲寄生于宿主體內,依賴宿主進行營養繁殖,會對宿主造成一系列的傷害[4]。例如,肝片吸蟲(Fasciola hepatica)感染會導致牛羊食欲減弱、精神萎靡等[5];大熊貓(Ailuropoda melanoleuca)感染西氏貝蛔蟲(Baylisascaris transfuga)后,會出現膽道蛔蟲和胃腸道梗阻,最終導致膽管、胰管和腸道破裂,引發腹膜炎甚至死亡[6];粗厚鰻居線蟲(Anguillicola crassus)感染日本鰻鱺(Anguilla japonica)會嚴重損害魚鰾的正常功能[7]。寄生蟲感染還能直接或間接影響宿主的行為[8]和生理特征[9]。玉黍螺(Littorina littorea)被吸蟲寄生后,大膽性降低[10];而彎鰭若花鳉(Poeciliopsis retropinna)被吸蟲寄生后,大膽性無顯著變化[11];剛地弓形蟲(Toxoplasma gondii)感染會導致褐家鼠(Rattus norvegicus)探索性增強[12],但小鼠探索性減弱[13]。寄生蟲感染導致馴鹿(Rangifer tarandus)糞便皮質醇濃度下降[14],但紅疣猴(Colobus polykomos)皮質醇濃度升高[15]。因此,寄生蟲對宿主行為和生理特征的影響存在物種間差異。
寄生蟲操縱假說認為,許多寄生蟲進化出了操縱宿主行為的能力,使宿主行為發生改變,以增加自身的存活率[16]或傳播率[17-18]。某些寄生蟲使中間宿主更容易受到捕食者(最終宿主)的攻擊,如被毛蟲寄生的蟋蟀(Gryllus bimaculatus)會主動跳進水里從而更易被捕食[19];剛地弓形蟲感染可增強小鼠的活動性,使它們更容易遭到貓的捕食[13]。個性特征是指動物個體間的行為差異,包括大膽性(boldness)、探索性(exploration)、攻擊性(aggressiveness)、活躍性(activity)和社會性(sociability)等[20]。寄生影響動物的個性特征,如與未被寄生的個體相比,被裂頭絳蟲(Schistocephalus solidus)寄生的三刺魚(Gasterosteus aculeatus)活動性更強,被捕食率更高[21]。寄生蟲對宿主行為的改變不一定均對宿主有害[22]。例如被褐飛虱(Apanteles euphydryidis)感染后,格斑堇蛺蝶(Euphydryas phaeton)的配偶定位效率明顯提高,寄生蟲存活率和傳播率增大[23]。因此,寄生蟲對宿主行為影響的差異性與寄生蟲傳播策略有關。
宿主可以通過激活免疫系統來抵抗寄生蟲的攻擊[24],并付出相應的代價[25]。當宿主免疫系統被激活后,其皮質醇水平降低[26]、生長緩慢、體重降低[27]、能量消耗增加[28]。宿主的總體能量代謝是有限的,免疫功能與生理活動(如生長、繁殖)所需能量之間存在著一種權衡[27-29]。目前,關于寄生是否會改變宿主能量平衡的研究甚少,尤其是以小哺乳動物為宿主的研究更加少見。
高原鼠兔(Ochotona curzoniae)隸屬兔形目鼠兔科鼠兔屬,是青藏高原特有的小哺乳動物,也是維持高原草甸生態系統的關鍵物種[30]。高原鼠兔在65日齡后為成體階段,體重生長基本停止,65日齡雌性高原鼠兔平均體重為121.72 g,雄性高原鼠兔平均體重為119.00 g[31],進入冷季后,越冬動物多為成年個體[32]。高原鼠兔為晝行性動物,在7月?10月活動高峰期為08:00?11:00和16:00?18:00[33]。艾美爾球蟲(Eimeria)作為高原鼠兔的主要寄生物,具有較強的專一性,寄生于宿主腸道,其生活史分為裂殖生殖、配子生殖以及孢子生殖3個階段[34]。目前已有學者研究艾美爾球蟲對高原鼠兔生理特征的影響,如有研究發現感染艾美爾球蟲后,雌性高原鼠兔的妊娠率和胚胎重量顯著降低[9,34],但關于艾美爾球蟲對高原鼠兔行為的影響卻鮮有報道。此外,高原鼠兔種群數量暴發是制約青藏高原草牧業健康發展的關鍵問題之一[35]。生物控制作為防治有害生物的方式之一,能夠有效降低有害生物種群的數量。艾美爾球蟲對宿主的種屬特異性、可調節害鼠種群動態以及對害鼠具有致死性等生物學特征為將其開發成為害鼠生物控制劑提供可能[34]。基于此,本研究選擇高原鼠兔為研究對象,研究艾美爾球蟲對高原鼠兔個性與生理特征的影響,旨在闡明寄生蟲對宿主產生的影響,并為進一步開發高原鼠兔的生物防治技術提供科學依據。
1 材料和方法
1.1 試驗動物
2020年9月,在海北高寒草地生態系統研究站附近采用繩套法捕獲30只健康、體重大于130 g 的高原鼠兔(成體)。帶回實驗室后單籠飼養于中國科學院西北高原生物研究所動物飼養房中450 mm×289 mm×180 mm 塑料籠內,籠內鋪有木屑,提供足量的水和標準鼠飼料(來源于北京科澳協力飼料有限公司),定期清理糞尿和墊料,適應7 d 后開始試驗,室溫和光周期均為自然環境溫度和光照。
1.2 感染試驗
已有研究表明,艾美爾球蟲的裂殖生殖主要破壞宿主腸道組織[36],當給高原鼠兔灌喂4×106個·mL?1艾美爾球蟲時,第4天出現高原鼠兔死亡,第8天時高原鼠兔死亡率達到最大值[37]。將試驗動物隨機分為3組,每組10只,分別灌喂2×106個·mL?1艾美爾球蟲卵囊懸濁液(G 組)、生理鹽水(C組)、驅蟲劑(Q組)(主要成份為磺胺氯吡嗪鈉,比例按照0.001 2×高原鼠兔體重,再加生理鹽水至1 mL)各1 mL。根據艾美爾球蟲的致病性周期,將試驗劃分為4個階段,即0、5、8和18 d,在這4個階段分別測定高原鼠兔的個性和生理特征。
試驗期間,為避免試驗個體自身以及感染艾美爾球蟲后排出的艾美爾球蟲卵囊影響試驗結果,每天換洗鼠籠,并用沸水燙洗。于每個試驗階段的前一天07:00,在每個鼠籠內放置50 g 飼料;在試驗測定當天收集剩余飼料,計算食物攝入量,收集每個鼠籠內糞便4 g,用于測定激素含量和艾美爾球蟲卵囊數。隨后測定高原鼠兔的行為和生理參數,包括探索性、體重、糞便皮質醇濃度和靜止代謝率。
1.3 艾美爾球蟲卵囊數量測定
采用麥克馬斯特法(McMaster’s method)計數糞便中艾美爾球蟲卵囊數。取2 g 糞便,加20 mL飽和鹽水混勻,經0.425和0.150 mm 濾篩過濾后,取1 mL 濾液,再加入9 mL 水,將稀釋液充分混勻后,用毛細吸管吸出少量液體滴入麥克馬斯特氏計數板的計數室內,置于顯微鏡臺上,靜置幾分鐘后,用低倍鏡將兩個計數室內的艾美爾球蟲卵囊全部計數完,取平均值后換算成每克糞便中的艾美爾球蟲卵囊數。
1.4 探索性測定
采用曠場試驗測定高原鼠兔的探索性,考慮到高原鼠兔的活動節律,在08:00?11:00測定探索性。曠場由不透明的亞克力板組成,底部面積為50 cm×50 cm,將底部中心40 cm×40 cm 的區域記為中心區,中心區以外的區域記為邊緣區。將待測個體輕輕放入曠場,使用攝像機記錄個體3 min 內的行為,使用EthoVision IX 動物運動軌跡跟蹤系統(Noldus Information Technology Co.Ltd.,Holland)分析高原鼠兔的各項行為,包括在中心區停留時間和穿越頻次、在邊緣區停留時間和穿越頻率等。動物在曠場中心區停留時間和穿越中心次數,可以代表探索性[38]。
每次測定完畢后,使用75%的酒精擦拭曠場,清理殘留的糞尿、毛發等,以免影響后續測定。
1.5 生理特征測定
1.5.1 食物攝入量和體重
食物攝入量的測定是在每個試驗階段的前一天,在清洗好的鼠籠內將50 g 鼠飼料放置于飼料盒中,24 h 后收集剩余飼料,放入烘干箱內去除水分后稱重。使用電子稱(精度0.01 g)稱量高原鼠兔體重。
1.5.2 靜止代謝率的測定
使用功能性運動測試攜式動物呼吸代謝測量系統(Sable Systems International,Las Vegas, NV, USA)測量高原鼠兔的靜止代謝率(resting metabolic rate,RMR)。試驗開始時,將動物放入透明呼吸室中,呼吸室位于恒溫箱內,根據高原鼠兔的熱中性區在25~30℃范圍內[39],可以維持最低基礎或靜止代謝率水平[40],將恒溫箱溫度設為27℃。每個通道的取樣間隔設置為3 min,設置氣泵進入呼吸室的空氣流速為600 mL·min?1。1號呼吸室作為空白對照,另外3個呼吸室中放入高原鼠兔。適應30 min 后,開始測定代謝率。重復測量10次,共計2 h。根據公式[41]計算代謝率(metabolic rate,MR):
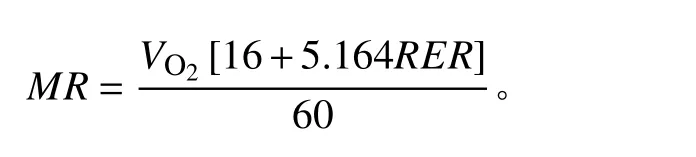
式中:MR為代謝率;RER(respiratory exchange ratio)為呼吸交換率,即CO2產生率(VCO2,mL·min?1)除以O2消耗率(VO2,mL·min?1)。選擇每個個體10次測量結果中MR的最低值,作為其靜止代謝率。
1.5.3 皮質醇激素
采用ELISA 酶聯免疫法測定高原鼠兔糞便皮質醇含量,使用玻璃棒將糞便樣品混勻,稱取1 g糞便,加入9 mL 生理鹽水,制備成濃度為10%的糞便勻漿,經離心機離心15 min,轉速3000 r·min?1;離心結束取上清液1 mL,使用大鼠ELISA 檢測試劑盒(廣州徠智生物科技有限公司生產)測定樣品中的皮質醇含量(同一份樣品重復測量2次,取平均值)。檢測靈敏度為1.0 ng·mL?1。板間和板內差異均為15%。
1.6 統計分析
對所有數據進行正態分布和方差齊性檢驗。采用Kruskal-Wallis秩和檢驗比較4個時間段、3個處理組之間艾美爾球蟲卵囊數的差異。采用主成分分析法對探索性數據進行降維[42],得到與探索性相關的維度。采用lmerTest 程序包,通過混合線性效應模型分析艾美爾球蟲與高原鼠兔個性、生理特征間的關系,分別將探索性、食物攝入量、體重、皮質醇濃度作為響應變量,將試驗時間與不同處理作為固定效應,將個體差異作為隨機效應;若處理效應顯著,采用emmeans包進行簡單效應分析,獲得各組間差異性結果。試驗結果以平均值 ± 標準誤表示,P<0.05為差異顯著,P<0.001為差異極顯著。所有數據統計分析使用R 軟件3.6.2 進行。
2 結果與分析
2.1 艾美爾球蟲卵囊數量
艾美爾球蟲卵囊數量變化結果表明,隨著時間的變化,艾美爾球蟲卵囊數量間有顯著差異(F=3.337,P=0.006)(圖1)。從第0天到第5天,艾美爾球蟲卵囊數呈增長趨勢,第5天后又逐漸下降。第0天、第8天和第18天,不同處理組間艾美爾球蟲卵囊數量無顯著差異(P>0.05);第5天,G 組艾美爾球蟲卵囊數量顯著高于C組和Q組(P< 0.05)。
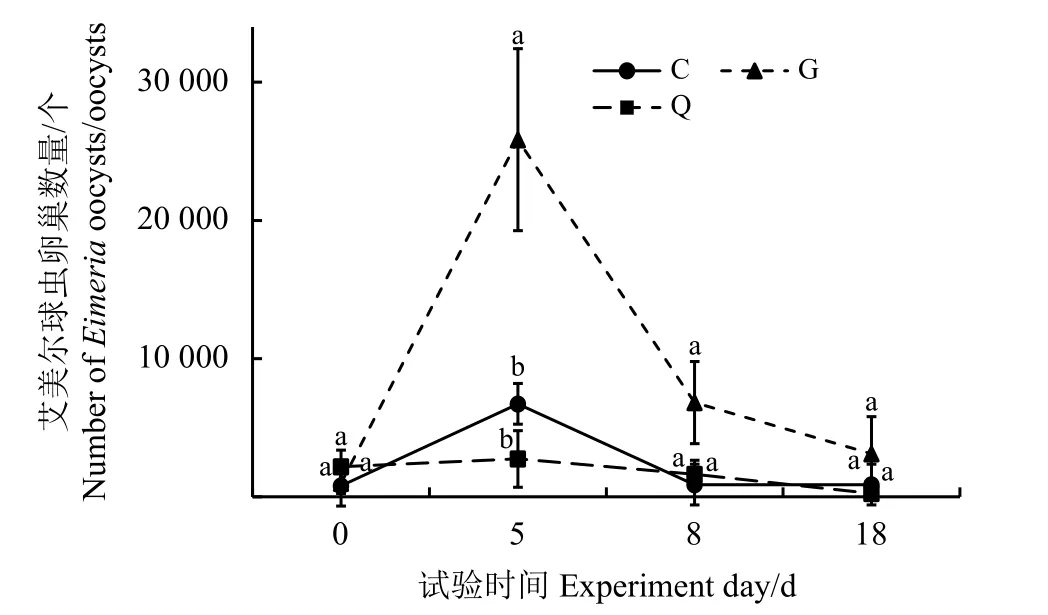
圖1 艾美爾球蟲卵囊數量比較Figure 1 Comparison of the number of Eimeria oocysts
2.2 生理特征
艾美爾球蟲對高原鼠兔生理特征影響的結果表明,隨著時間的變化,艾美爾球蟲對高原鼠兔食物攝入量有顯著影響(F=2.478,P= 0.0476)(圖2)。第18天,G 組與C組高原鼠兔食物攝入量之間無顯著差異(P>0.05),但均顯著低于Q組(P<0.05)。艾美爾球蟲對高原鼠兔的體重無顯著影響(F=1.132,P=0.345),但對其糞便皮質醇濃度有顯著影響(F=2.417,P= 0.0472)。第0天和第5天,3組高原鼠兔糞便皮質醇濃度間無顯著差異(P>0.05);第8天,Q組和G 組高原鼠兔糞便皮質醇濃度間無顯著差異(P>0.05),但均顯著高于C組(P<0.05);第18天,C 組和G組高原鼠兔糞便皮質醇濃度間無顯著差異(P>0.05),但均顯著高于Q組(P<0.05)。艾美爾球蟲對高原鼠兔的靜止代謝率有顯著影響(F=2.951,P=0.010 3)。從第0天到第8 天,3個處理組靜止代謝率間無顯著差異(P>0.05);第18天,G 組高原鼠兔靜止代謝率顯著低于C 組(P< 0.05)。
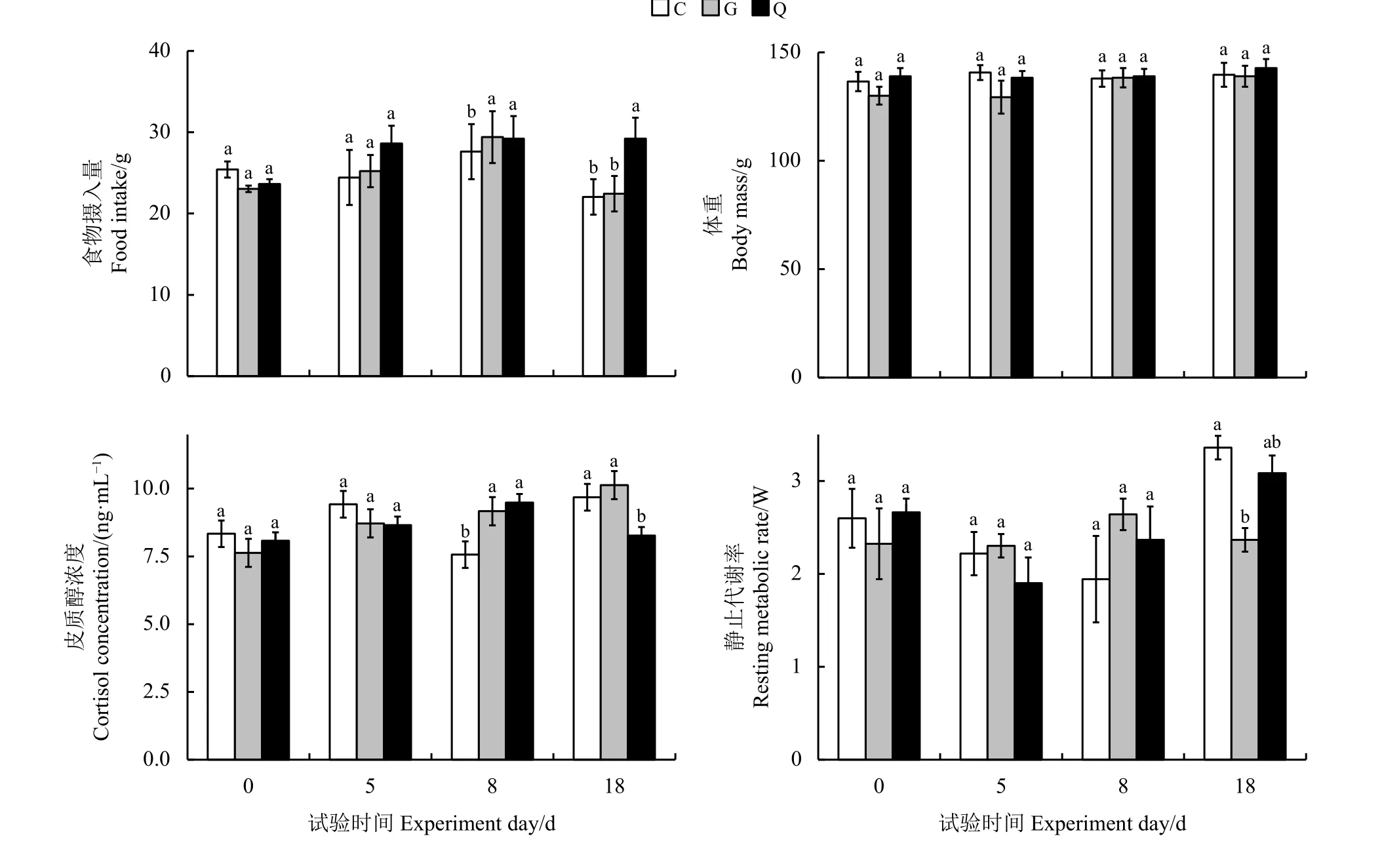
圖2 不同處理組間高原鼠兔的各項生理指標比較Figure 2 Comparison of physiological traits of plateau pika in different treatment groups
2.3 個性特征
對高原鼠兔的探索性數據進行主成分分析,前兩個主成分解釋了80%的方差(表1),將第一主成分命名為探索性用于后續分析。
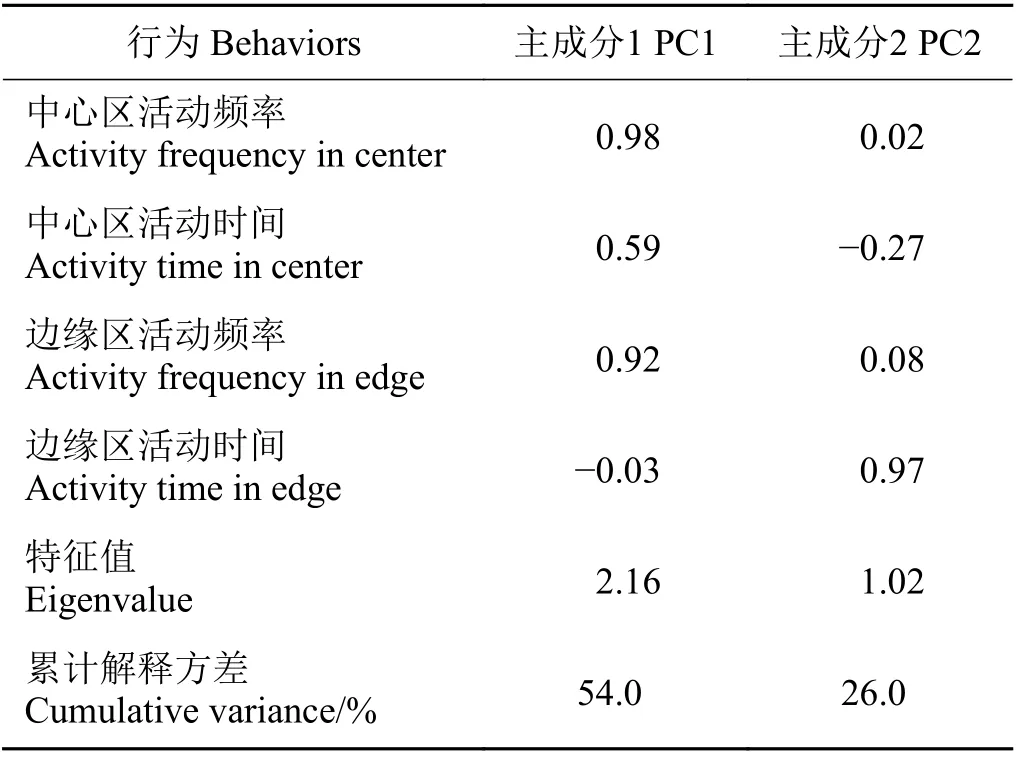
表1 4個高原鼠兔行為變量的主成分分析Table 1 Principal component analysis of measures of the four behaviors of plateau pika
艾美爾球蟲對高原鼠兔探索行為影響的結果表明,隨著時間的變化,艾美爾球蟲對高原鼠兔的探索性有顯著影響(F=2.251,P=0.044)(圖3)。總體上看,從第0天到第5天,Q組和C組高原鼠兔探索性上升,之后逐漸下降;從第0天到第5天,G 組高原鼠兔探索性顯著下降(P<0.05),并維持在相對較低的水平。第5天,G 組高原鼠兔探索性顯著低于C組(P<0.05),而Q組和C組高原鼠兔探索性間無顯著差異(P>0.05)。
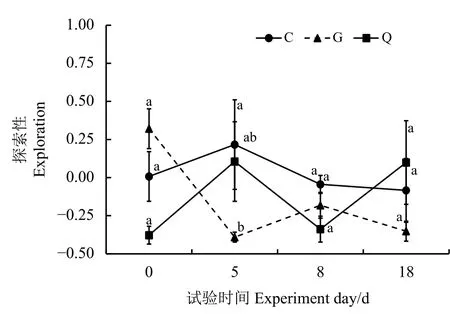
圖3 不同處理組間高原鼠兔探索性比較Figure 3 Comparison of plateau pika exploration in different groups
3 討論與結論
本研究結果表明,3組艾美爾球蟲卵囊數量間有顯著差異,在第5天均達到最大值,灌喂艾美爾球蟲組高原鼠兔排出的艾美爾球蟲卵囊數顯著高于對照組和驅蟲組,這與邊疆暉等[43]研究結果略有不同,其結果表明,灌喂艾美爾球蟲卵囊后,第5天時,高原鼠兔成體才開始大量排艾美爾球蟲卵囊。鄭若愚等[44]認為感染艾美爾球蟲后,動物排艾美爾球蟲卵囊數與其年齡呈正相關關系,如與低齡個體相比,感染相同劑量艾美爾球蟲的大齡兔可排出大量艾美爾球蟲卵囊。本試驗所用高原鼠兔均為成年個體,年齡較大,可能導致高原鼠兔排艾美爾球蟲卵囊數峰值提前。
寄生蟲會影響宿主的食物攝入量[45]。一般而言,感染寄生蟲后,宿主的食物攝入量減少,反之亦然[46]。Arneberg 等[47]研究發現,試驗去除寄生蟲后,馴鹿食物攝入量顯著增加,這與本研究中驅蟲組高原鼠兔食物攝入量顯著高于對照組的結果一致。動物感染寄生蟲后其食物攝入量與現存寄生蟲數量有關[46]。在自然環境中,艾美爾球蟲作為高原鼠兔的主要腸道寄生物[43],高原鼠兔對其感染可能產生了一定的免疫力,灌喂較低劑量的寄生蟲,使得灌喂艾美爾球蟲組與對照組高原鼠兔食物攝入量間無顯著差異。
本研究中,灌喂艾美爾球蟲對高原鼠兔體重無顯著影響,這可能是由于高原鼠兔長期進化,適應了艾美爾球蟲的寄生。當寄生蟲侵入宿主體內時,宿主可以通過激活免疫系統來抵御寄生蟲的攻擊[25]。艾美爾球蟲屬自限性感染[48],宿主的最優策略是將能量主要用于免疫防御[49]。在長期的寄生共存關系下,宿主體內已產生相應的抗體[50],當再次灌喂艾美爾球蟲后,體內已有的抗體使得高原鼠兔不需要將過多能量用于免疫,因此高原鼠兔體重沒有顯著降低。Silva 等[51]研究表明,與未驅蟲的母馬相比,驅蟲后的母馬體重顯著增加,而本研究中驅蟲組高原鼠兔體重與對照組間無顯著差異。驅蟲對動物體重的影響與飼料中粗蛋白含量有關,當飼料中粗蛋白質含量<11%時,驅蟲對動物體重沒有影響;當飼料中粗蛋白質含量>19%時,驅蟲后的動物體重顯著增加[52]。本研究飼料中粗蛋白含量為14.45%,可能導致驅蟲后的高原鼠兔體重沒有顯著增加。
寄生蟲感染與宿主皮質醇含量具有一定的關聯性[53]。如感染寄生蟲后,沙錐齒蜥(Zootoca vivipara)、靈敏負鼠(Gracilinanus agilis)的皮質醇濃度更高[54-55]。糖皮質激素適應假說認為,皮質醇濃度增加會導致動物適應能力下降、存活率和繁殖成功率降低,更容易被寄生蟲感染[56]。本研究結果表明,第8天時,與對照組相比,灌喂艾美爾球蟲組高原鼠兔糞便皮質醇濃度顯著高于對照組。寄生蟲感染通過消耗宿主營養、增加與免疫反應相關的成本,導致皮質醇濃度上升[53]。在第5天時,灌喂艾美爾球蟲組高原鼠兔排艾美爾球蟲卵囊數顯著高于對照組,而在第8天時,其皮質醇濃度顯著高于對照組,這可能是由于動物對寄生蟲產生的應激反應具有時滯性[57]。第8天,驅蟲組高原鼠兔皮質醇濃度顯著高于對照組。在寄生蟲與宿主的協同進化過程中,二者相互適應、共存[58],某些寄生蟲感染甚至還會降低宿主對疾病的易感性,增加宿主的生存率和寄生蟲的傳播率[59]。如馬來布魯絲蟲(Brugia malayi)分泌的免疫調節分子能限制人肺部的嗜酸性粒細胞反應,減緩過敏性炎癥[60]。艾美爾球蟲作為高原鼠兔的主要寄生物,可能在某些方面對宿主有利,高原鼠兔艾美爾球蟲卵囊數減少了39.33%,這可能會對高原鼠兔產生刺激,導致其皮質醇濃度增加,具體機制還需進一步研究。第18天時,驅蟲組高原鼠兔皮質醇濃度顯著低于對照組,這可能是由于驅蟲后高原鼠兔體內的艾美爾球蟲卵囊數減少,對宿主的刺激減弱,皮質醇濃度降低,符合糖皮質激素適應假說。
寄生蟲感染后,宿主激活免疫系統抵抗寄生蟲攻擊的過程會消耗大量能量,導致體內能量平衡改變[61-63]。例如,感染跳蚤(Xenopsylla ramesis)的沙鼠(Gerbillus dasyurus)能量消耗增加[64];感染蠕形住腸線蟲(Enterobius vermicularis)的地松鼠(Xerus inauris)靜止代謝率降低[65]。能量分配模型認為動物具有穩定的能量收支平衡,總能量保持不變,某一方面能量消耗增加,其他方面能量消耗就會減少[66-67]。灌喂艾美爾球蟲后,高原鼠兔用于免疫防御的能量增加,導致其靜止代謝率顯著低于對照組。
寄生蟲操縱假說認為寄生蟲感染可以影響宿主行為,例如囊尾蚴(Cysticercus cellulosae)聚集在雌海螺(Busycon canaliculatum)的足部會使宿主失去挖掘洞穴的能力,被捕食率是未被寄生海螺的5~7倍[68],有利于寄生蟲傳播[69]。動物的探索性與被捕食風險有關,探索性弱的動物被捕食風險較低[70]。灌喂艾美爾球蟲后,高原鼠兔探索性顯著下降,降低了被捕食的風險,提高了宿主的存活率,有利于艾美爾球蟲的傳播。艾美爾球蟲在高原鼠兔種群內的傳播率增加,能更有效地降低高原鼠兔種群的繁殖力,進而控制高原鼠兔種群數量。
綜上所述,高原鼠兔感染艾美爾球蟲后,其皮質醇濃度升高、靜止代謝率下降、探索性減弱,降低了被捕食風險,從而增加了艾美爾球蟲在高原鼠兔種群內的傳播率,為應用艾美爾球蟲作為潛在的高原鼠兔生物防治制劑以及控制高原鼠兔種群數量提供了新的依據。
