環境污染物干擾魚類體色的研究及其生態毒理學意義
2021-09-23 02:51:56楊莉王力軍金映虹侯宇張春暖張紀亮
生態毒理學報 2021年3期
楊莉,王力軍,金映虹,侯宇,張春暖,張紀亮,,*
1. 河南科技大學動物科技學院,洛陽 471000 2. 海南師范大學熱帶島嶼生態學教育部重點實驗室,海口 571158
魚類是水生態系統中的重要組成部分。水環境污染對魚類生長、發育以及繁殖產生的不利影響,引起了國內外學者的關注。最近,污染物對魚類體色的影響也開始引起關注。魚類體色具有多種功能,如偽裝、威懾、物種識別、溫度調節和求偶等,對其生存與成功繁殖至關重要[1]。在所有脊椎動物中,魚類體色調控最為復雜,涉及的體色相關基因也最為豐富[2]。目前,魚類中至少發現了6種不同的色素細胞,包括黑色素細胞、黃色素細胞、紅色素細胞、虹彩細胞、藍色素細胞和白色素細胞。加上魚類體色還對污染物非常敏感,使其成為了一種具有巨大應用前景的污染監測生物指示物[3-5]。因此,污染物干擾魚類體色的相關研究是一個值得挖掘的新方向。
目前,已經發現多種污染物能夠干擾魚類的體色(表1)。內分泌干擾物(雌激素、有機錫和雙酚A)和對辛基苯酚均能夠引起魚類顏色強度的改變[6-12]。某些農藥,如殺蟲劑二氯二苯三氯乙烷(dichlorodiphenyltrichloroethane, DDT)、殺蟲劑甲基對硫磷、除草劑阿特拉津和久效磷等,都在不同程度上影響魚類體色。研究發現,DDT和阿特拉津能夠減少孔雀魚(Poeciliareticulata)體表橙色斑點的面積,降低體色的亮度;甲基對硫磷能夠降低阿馬里洛魚(Girardinichthysmultiradiatus)鰭和體表顏色強度[13-15]。久效磷對孔雀魚不同色素細胞均產生干擾作用,表現為色素顆粒顯著減少、分散與溶解,黑色素細胞和黃色素細胞退化、數量減少,虹彩細胞胞膜溶解以及反射小板減少、板層變薄且間隙變得不規則甚至閉合[16]。高濃度氯氰菊酯長期暴露會增加鯉魚(Cyprinuscarpio)的色素沉著[17]。硝酸鹽能夠對魚類皮膚造成嚴重的組織學損傷,表皮和真皮會出現不同程度的壞死脫落,且伴隨黑色素細胞和粘液細胞的分散和減少[18]。硫酸鋅暴露導致印度囊鰓鯰(Heteropneustesfossilis)身體出現明顯的色素沉著障礙,并伴隨粘液分泌增多[19]。金屬化合物三氧化二砷會引起斑馬魚(Daniorerio)皮膚粘液滲出、身體褪色[20]。虹鱒(Salmogairdneri)長期暴露于防污劑美托咪啶,顏色變淺,色素細胞發生聚集反應[21];在酸暴露下,虹鱒體色減褪[22]。Kaur和Dua[23]將淡水魚翠鱧(Channapunctatus)暴露于廢水中,發現鱗片破損、松動、脫落和鈣質結構紊亂,色素細胞出現異常分散現象,色素顆粒明顯減少和分散。在暹羅斗魚(Bettasplendens)色素細胞的研究中,發現甲基汞、亞砷酸鹽、威百畝(甲基二硫代氨基甲酸鈉)和百草枯能夠引起色素細胞聚集;殺蟲脒和磷酸鹽導致不同程度的色素細胞分散[24]。
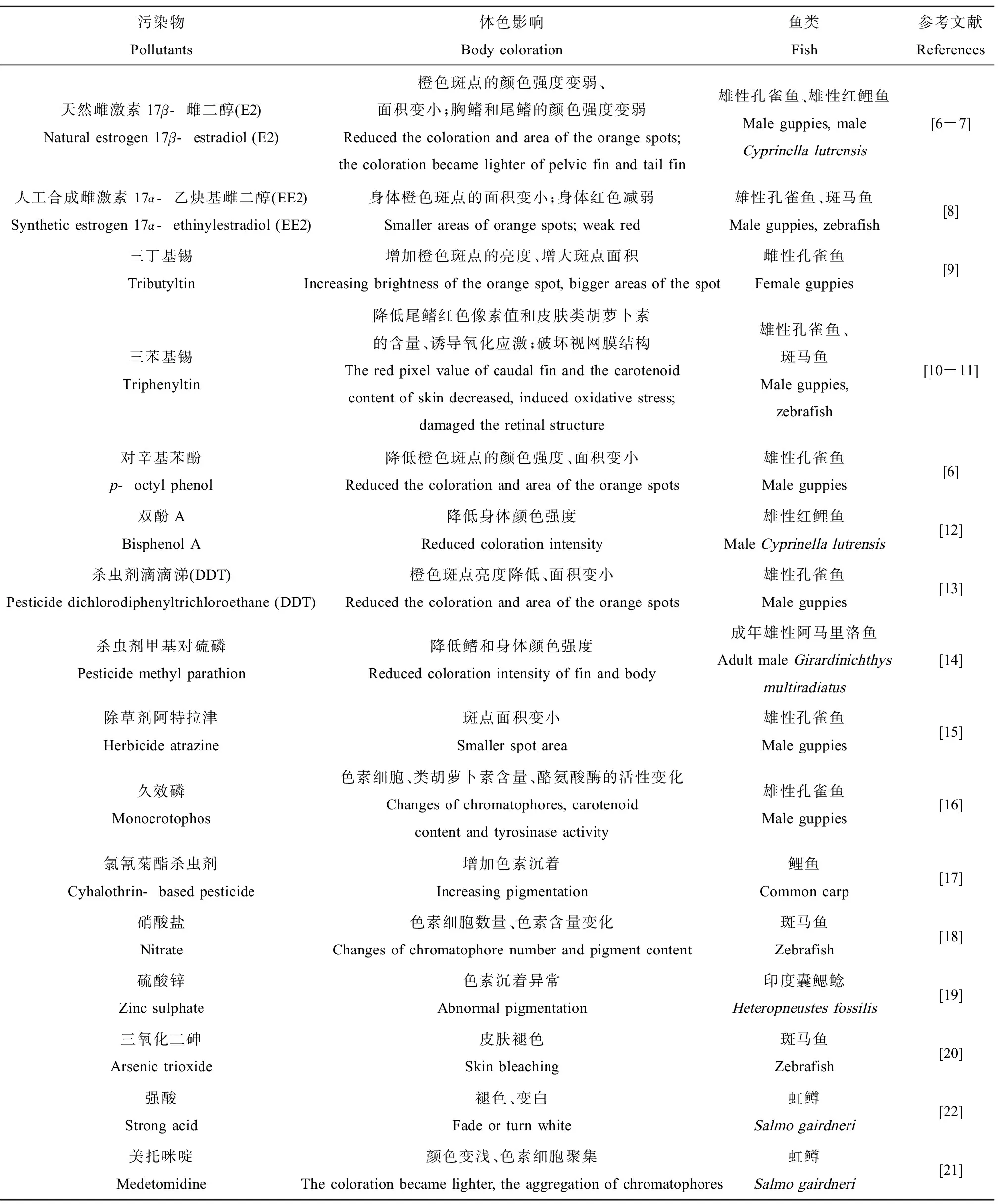
表1 污染物對魚類體色的影響Table 1 The effects of pollutants on body coloration in fish
盡管大量研究已經證實污染物能夠干擾魚類體色,但對其潛在生理與分子機制的研究卻較少。研究表明,有些污染物(雙酚A、雌二醇和類黃酮等)可以調節核受體的轉錄活性,進而可能影響色素細胞分化和發育等過程[25-26]。另外,化學物質(重金屬、芳香烴等)還可以與色素細胞的復雜分子機制相互作用。因此,在環境監測中,色素細胞被用作細胞傳感器[24]。魚類皮膚色素的合成、代謝及轉運和色素細胞活動等過程都可能存在污染物的作用靶點。鑒于魚類體色形成的分子機制較復雜,因此,對魚類體色形成的分子基礎進行系統總結,不僅有助于探究污染物干擾體色的可能靶點和毒理學機制,而且將促進體色相關生物標志物的開發及其在環境監測中的應用。
1 魚類皮膚中主要色素的合成、代謝及轉運(The synthesis, metabolism and transport of main pigments in fish skin)
魚類擁有多種色素,儲存在特定的色素細胞中,是體色呈現多樣性的重要組織學基礎。目前,研究較多的色素細胞主要為黑色素細胞、黃色素細胞、紅色素細胞和虹彩細胞[2]。黑色素儲存于黑色素細胞;呈現紅色/橙色的類胡蘿卜素和呈現黃色的蝶啶儲存于黃色素細胞和紅色素細胞;虹彩細胞不含色素,通過透明嘌呤晶體反射小板形成衍射光柵,呈現不同光澤。
1.1 黑色素
黑色素對魚類體色表型產生重要影響,黑色素的生成被破壞會導致色素沉著度降低,甚至會引起白化病[27]。酪氨酸酶(tyrosinase, Tyr)是黑色素(魚類僅有真黑色素)合成的關鍵限速酶,酪氨酸在Tyr催化下,經由多巴(dihydroxyphenylalanine, Dopa)、多巴醌和吲哚醌等形式,合成真黑色素;該過程需要Tyr、多巴色素異構酶(dopachrome tautomerase, Dct)、酪氨酸酶相關蛋白1(tyrosinase related protein 1, Tyrp1)等酪氨酸酶家族成員以及銀蛋白(Silv,也稱Pmel17)的參與。另外,3種黑素體轉運蛋白(Oca2、Aim1和Slc24a5)對黑色素的合成也至關重要[28],如圖1(a)所示。
1.2 類胡蘿卜素
與其他色素不同,類胡蘿卜素不能在脊椎動物體內合成,使其成為體內有限的資源[29]。脊椎動物主要通過食物攝取類胡蘿卜素,轉運至體表色素細胞。目前,已經發現多個基因參與類胡蘿卜素的攝取(Sr-bi、Cd36)、結合與沉積(Star1、Mln64、Star4、Star5、Apod、Plin、Gsta2)以及降解(Bco2、Bcmo1)[30],如圖1(b)所示。
1.3 蝶啶
蝶啶不僅與黃色體色有關,其合成中間產物四氫生物蝶呤還可作為酪氨酸合成的輔助因子調控黑色素合成[31]。蝶啶合成包括3個分途徑:鳥嘌呤(guanosine triphosphate, GTP)環化水解酶I(GTP cyclohydrolase 1, Gch1)、6-丙酮酰四氫喋呤合成酶(6-pyruvoyltetrahydropterin synthase, Pts)和墨蝶呤還原酶(sepiapterin reductase, Spr)以GTP為底物,從頭合成四氫生物蝶呤;四氫生物蝶呤的再生,涉及蝶呤-4-氨基甲酰胺脫水酶(pterin-4-alpha-carbinolamine dehydratase, Pcbd)和二氫蝶啶還原酶(dihydropteridine reductase, Dhpr);墨喋呤及衍生物的合成,主要涉及Spr和黃嘌呤氧化酶/黃嘌呤脫氫酶(xanthine oxidase, Xod/xanthine dehydrogenase, Xdh)[31],如圖1(c)所示。另外,Gch1還參與黑色素細胞和黃色素細胞分化;Pcbd復合物的轉錄靶標是酪氨酸酶啟動子,影響黑色素細胞分化[31-32]。
1.4 晶體反射小板(GTP與水結晶形成)
GTP除了作為蝶啶合成中的重要底物外,還是虹彩細胞晶體反射小板的主要組成成分,而Xod/Xdh也是嘌呤分解代謝的關鍵限速酶[33],如圖1(d)所示。虹彩細胞通過嘌呤核苷磷酸化酶(purine nucleoside phosphorylase 4a, Pnp4a)把鳥嘌呤核苷轉化為GTP,Slc23l參與GTP的運輸[34-35]。另外,嘌呤合成酶(Gart、Paics、Ppat、Pfas、Atic和Adsl等)[35-36]以及嘌呤代謝相關基因(Ada2、Prtfdc1和Slc2a9)[37]均可能影響GTP的水平。
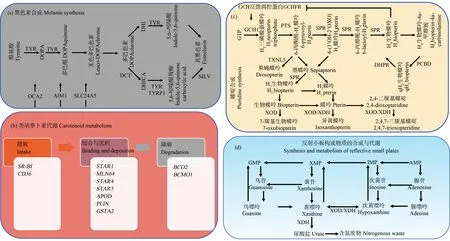
圖1 魚類色素細胞色素或呈色物質合成與代謝過程Fig. 1 The synthesis and metabolism of pigments or colored substances in fish
2 魚類色素細胞的發育與遷移(The development and migration of fish chromatophores)
除視網膜上皮,色素細胞均源于與外周神經系統背根神經節相關的神經嵴細胞[38]。色素細胞模式形成包括細胞模式的發育(如增殖、分化、遷移以及死亡)以及色素的合成與表達[39]。深刻認識色素細胞模式形成有關基因,對研究魚類體色形成的分子機制,特別是體表斑紋圖案形成,具有重要意義[40-42]。神經嵴細胞向色素細胞的分化由一系列細胞內和細胞外信號分子共同調控[43]。
目前,魚類色素細胞遷移與分化的研究主要以斑馬魚為研究對象。斑馬魚神經嵴細胞依靠Erbb3b增殖,有一部分胚后祖細胞衍生物在Tuba813a作用下遷移到皮下組織,產生成魚模式的色素細胞(圖2)。該祖細胞具有分化為3種色素細胞的潛能,且大部分色素細胞都來源于胚后祖細胞;集落刺激因子1受體a (colony stimulating factor 1 receptor a, Csf1ra)決定黃色素細胞的命運;小眼畸形相關轉錄因子a (microphthalmia-associated transcription factor a, Mitfa)決定黑色素細胞的命運,干細胞因子受體a (stem cell factor receptor a, Kita)促進成魚黑色素細胞前體的分化、存活;白細胞受體酪氨酸激酶(leukocyte receptor tyrosine kinase, Ltk)促進虹彩細胞的分化和存活[44]。此外,部分色素細胞直接來源于遷移的神經嵴細胞,如胚胎或者早期幼魚黑色素細胞會持續到成魚階段,且顏色與新分化的黑色素細胞相比偏棕色;幼魚黃色素細胞命運的決定依靠Csf1r,同時Pax7促進黃色素細胞的發育,前期會失去色素,進入隱蔽期,而在成魚期,依靠Csf1ra重新獲得色素(圖2)。色素細胞沿著背根神經節、橫隔膜、縱隔膜遷移到真皮,并按照一定的順序排列[44-45]。另外,Pcdh10、Igsf11和Erbb等能促進色素細胞或前體細胞的遷移[46-48]。色素細胞前體和色素細胞會因各種因子(Bcl、Gu和Ednrb等)而增殖,并可能依賴這些因子而存活[48]。隨著分子生物學技術以及組學技術的發展,已經篩選出大量與色素細胞發育與遷移相關的基因(表2),有待于進一步的研究。
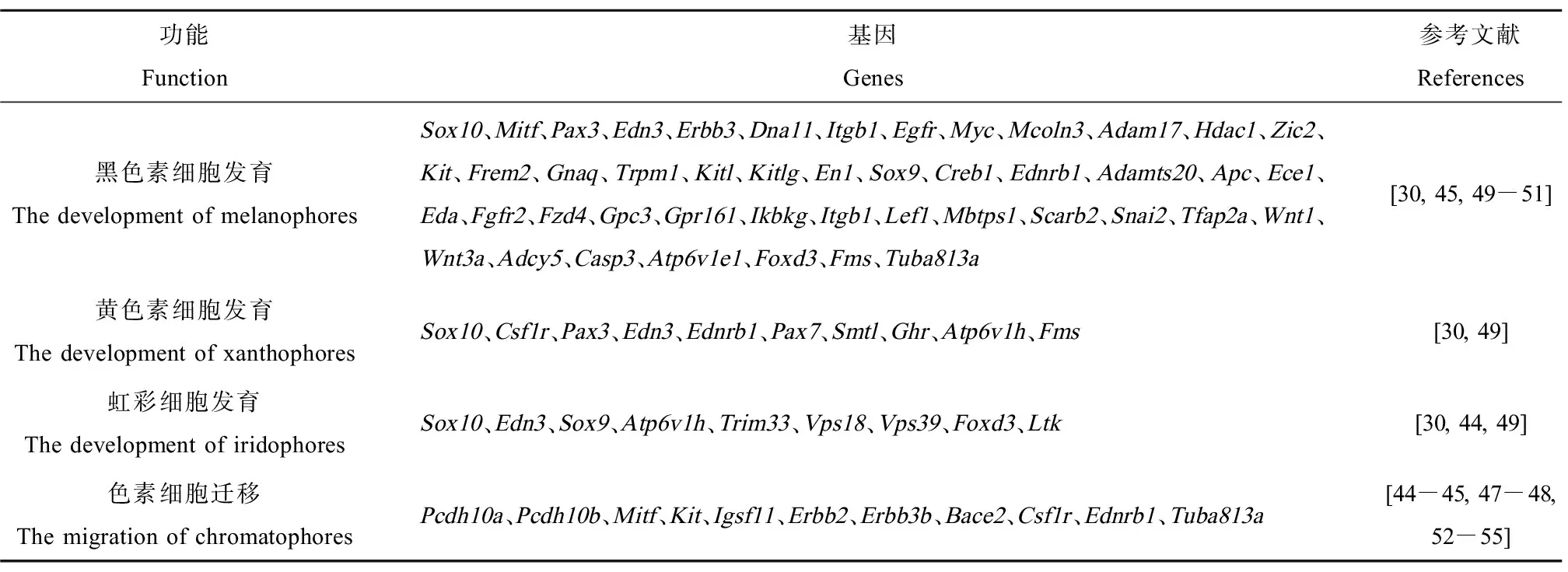
表2 色素細胞發育和遷移相關基因Table 2 The genes related to the development and migration of chromatophores
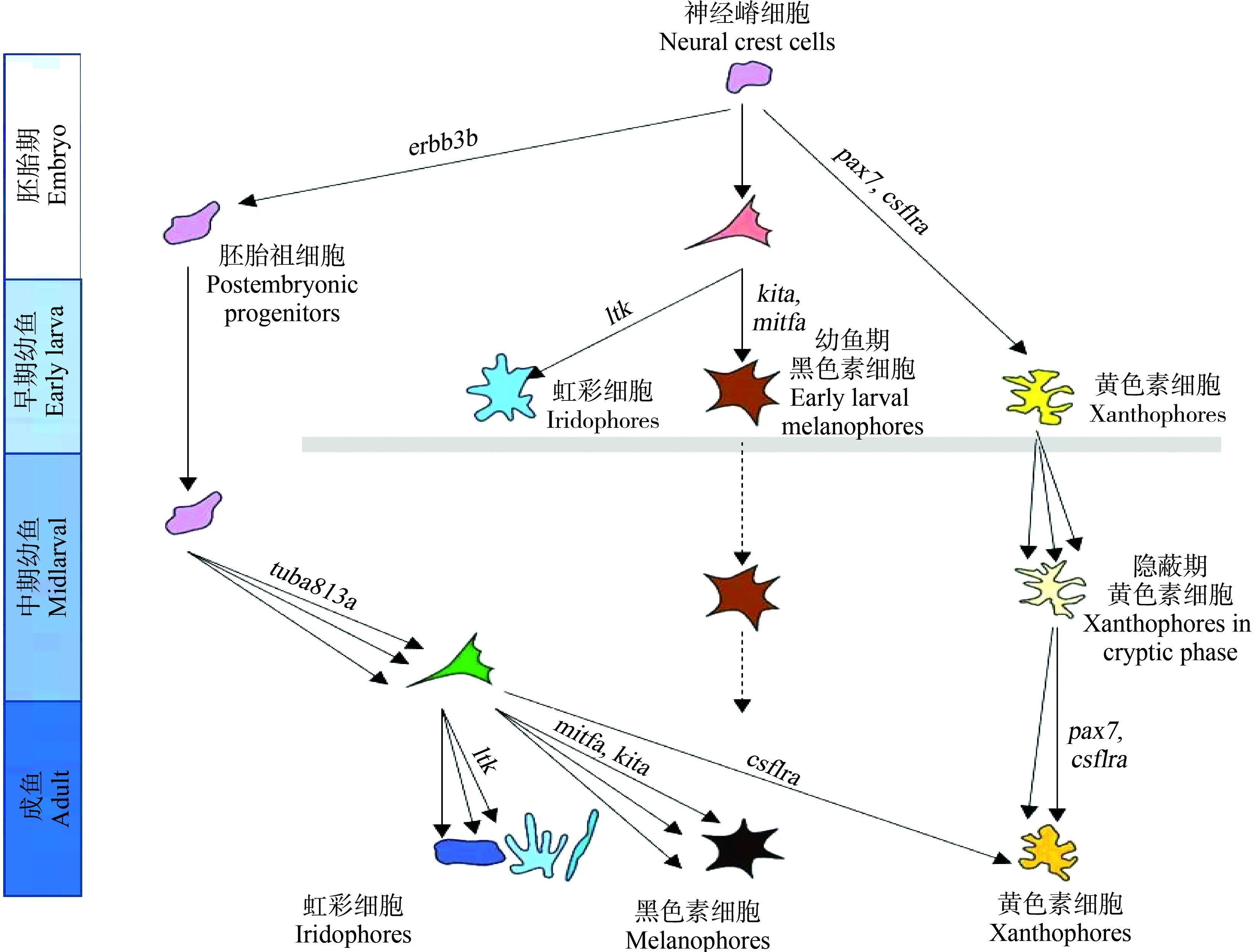
圖2 斑馬魚色素細胞的遷移與分化(仿Patterson和Parichy[44])Fig. 2 The migration and differentiation of chromatophores in zebrafish (Imitating Patterson and Parichy[44])
3 魚類體色調控機制(The regulatory mechanisms of body coloration in fish)
3.1 神經調控-黑素皮質素系統
魚類可通過生理體色變化和形態體色變化2種方式對體色進行調控。生理體色變化基于色素體在色素細胞內的聚集或分散,一般由短期的環境因素刺激(包括光照、溫度等)引起,通過交感神經系統和內分泌系統共同調控[56]。已經證實,在光照刺激下,去甲腎上腺素和黑色素聚集素(melanin-concentrating hormone, Mch)可誘導色素體聚集[57-59]。Mch不僅聚集色素體,而且抑制α-促黑細胞刺激素(α-melanocyte stimulating hormone,α-Msh)的分散活性,甚至抑制α-Msh的釋放,阻止黑色素體的分散。形態體色變化是由色素和色素細胞的組成、數量和形態等共同決定的,由長期刺激誘導,如長期的光照等[33]。研究表明,魚類在長期適應淺色或深色背景的過程中,機體可以通過減少或增加色素細胞的數量等改變體色[60-61]。在長期刺激過程中,黑色素細胞對Mch的敏感性降低;α-Msh的調節作用似乎占主導地位,α-Msh不僅參與黑色素體的擴散,而且參與黑色素細胞的發育[59]。
魚類黑素皮質素系統不僅調控色素的分散或聚集;還通過調控色素合成,參與形態學體色變化[2](圖3)。黑素皮質素1受體(melanocortin 1 receptor, Mc1r)以及其激動劑α-Msh和內源拮抗劑刺鼠信號蛋白(agouti signaling protein, Asip)是體色調控的主要因子[62]。α-Msh在垂體中合成,通過與皮膚黑色素細胞、黃色素細胞以及虹彩細胞等色素細胞膜上Mc1r結合而激活色素細胞擴散以及色素的合成[63];而Asip則主要參與背腹體色模式的調控[64]。在黑色素細胞中,激活的Mc1r不僅可以通過環磷酸腺苷(cAMP)通路介導色素的擴散,而且可以影響黑色素合成相關基因(Tyr、Tyrp1)的表達,產生更多的黑色素。黃色素細胞中色素擴散和黑色素相似。在虹彩細胞中,目前確定了cAMP對色素分散的促進作用[63]。
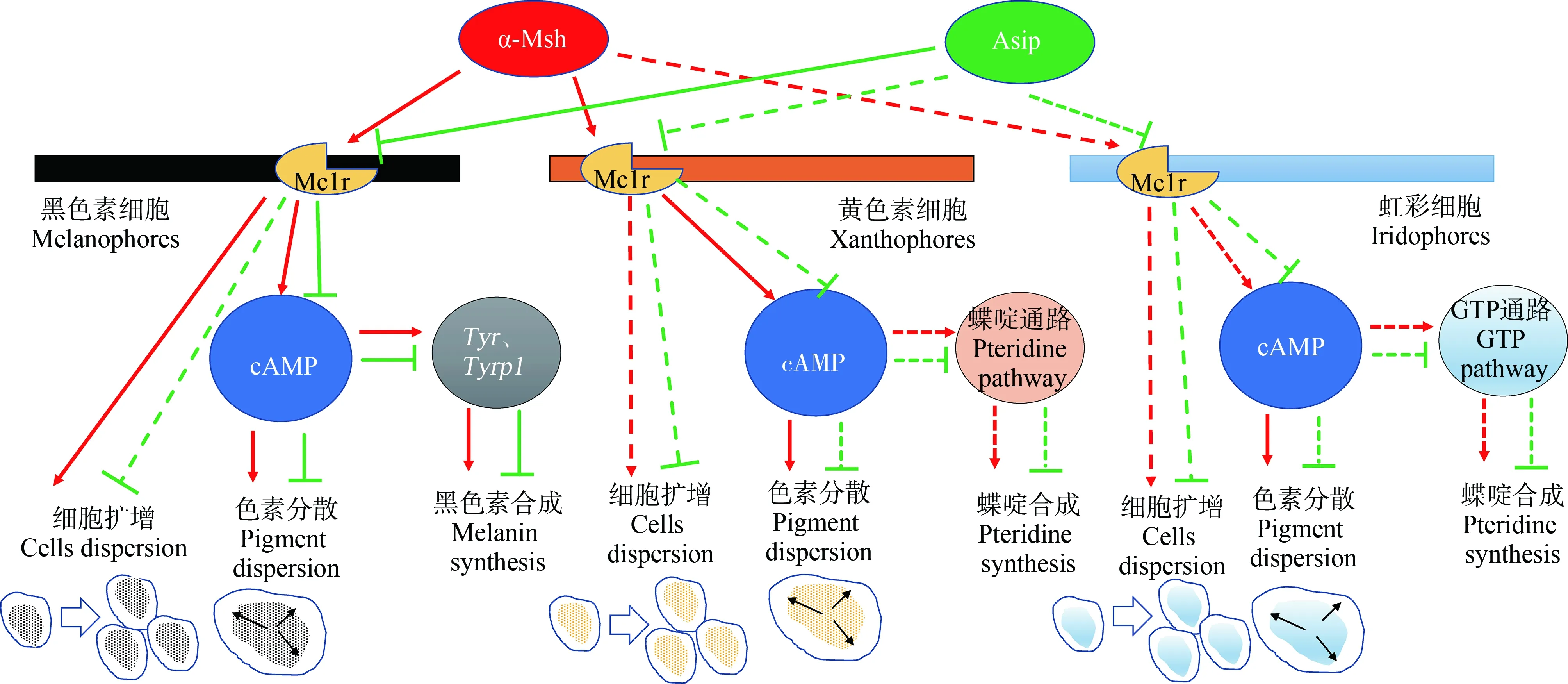
圖3 魚類黑素皮質素系統對體色的調控機制注:↑代表誘導或刺激;┬代表抑制;虛線代表未確定。Fig. 3 The regulation mechanism of melanocortin system on body coloration in fishNote: ↑ stands for inducing or stimulating; ┬ stands for inhibition; the dotted line represents uncertainty.
3.2 “類胡蘿卜素權衡”假說
除在體色中具有重要作用外,類胡蘿卜素在抗氧化系統中也起重要作用[65]。然而,類胡蘿卜素不能在脊椎動物體內合成,只能通過攝食獲得,使其在體內成為一種有限的資源。研究發現,類胡蘿卜素優先參與抗氧化作用中,只有多余的類胡蘿卜素才用于裝飾體色[66]。因此,在脊椎動物機體內,存在通過競爭類胡蘿卜素來實現抗氧化系統和裝飾體色之間的分配權衡[65]。“類胡蘿卜素權衡”假說得到了2類實驗的支持:一類是,類胡蘿卜素補充劑能增加著色、免疫以及抗氧化能力;另一類是,通過脅迫誘導的氧化應激,導致類胡蘿卜素的重新分配和減少類胡蘿卜素色素的著色。
3.3 光傳導和視覺系統
在視網膜光傳導中,視蛋白作為光感受器發揮重要作用,是光傳導的基礎。視蛋白的激活分別導致下游轉導蛋白(G-protein transducin)和磷酸二酯酶(phosphodiesterase)的活化,隨后伴隨著胞質信使環磷酸鳥苷(cGMP)的減少和感光細胞的激活,從而對光刺激作出反應,如視網膜接受光刺激后調節α-Msh的分泌[67]。此外,除視網膜外,某些脊椎動物皮膚可以直接轉換光信號,作為體色變化反應的一部分[68-71]。目前,在多種體色改變的魚類皮膚中,已經發現了視蛋白轉錄產物[72-73]。研究表明,魚類的體表色素細胞可以感光,這也提示皮膚光感受器在體色變化的調控中具有一定的作用[70]。目前,研究認為維生素A衍生的發色團與視蛋白結合,可能是魚類皮膚光傳導的基礎[67]。
視覺缺陷會干擾黑色素細胞分化所需的內分泌信號通路[2]。營養不足和環境刺激都會導致眼睛視網膜缺陷。如,飼料中脂肪酸二十二碳六烯酸、二十碳五烯酸和維生素A的比例不足會干擾魚類視網膜中視紫紅質的合成[74-75],這種缺陷能夠影響從視網膜到中樞神經系統的信號傳輸,而這種信號是誘導垂體產生Msh所必需的[76]。另外,視覺缺陷的魚類無法適應背景。在背景適應過程中,魚類根據環境光線水平調整皮膚中色素的分布,而視網膜到下丘腦的直接投射是魚類背景適應中信號輸入的基礎[77]。所以,視覺系統與神經系統之間的聯系是魚類體色變化調控的重要機制之一(圖4)。
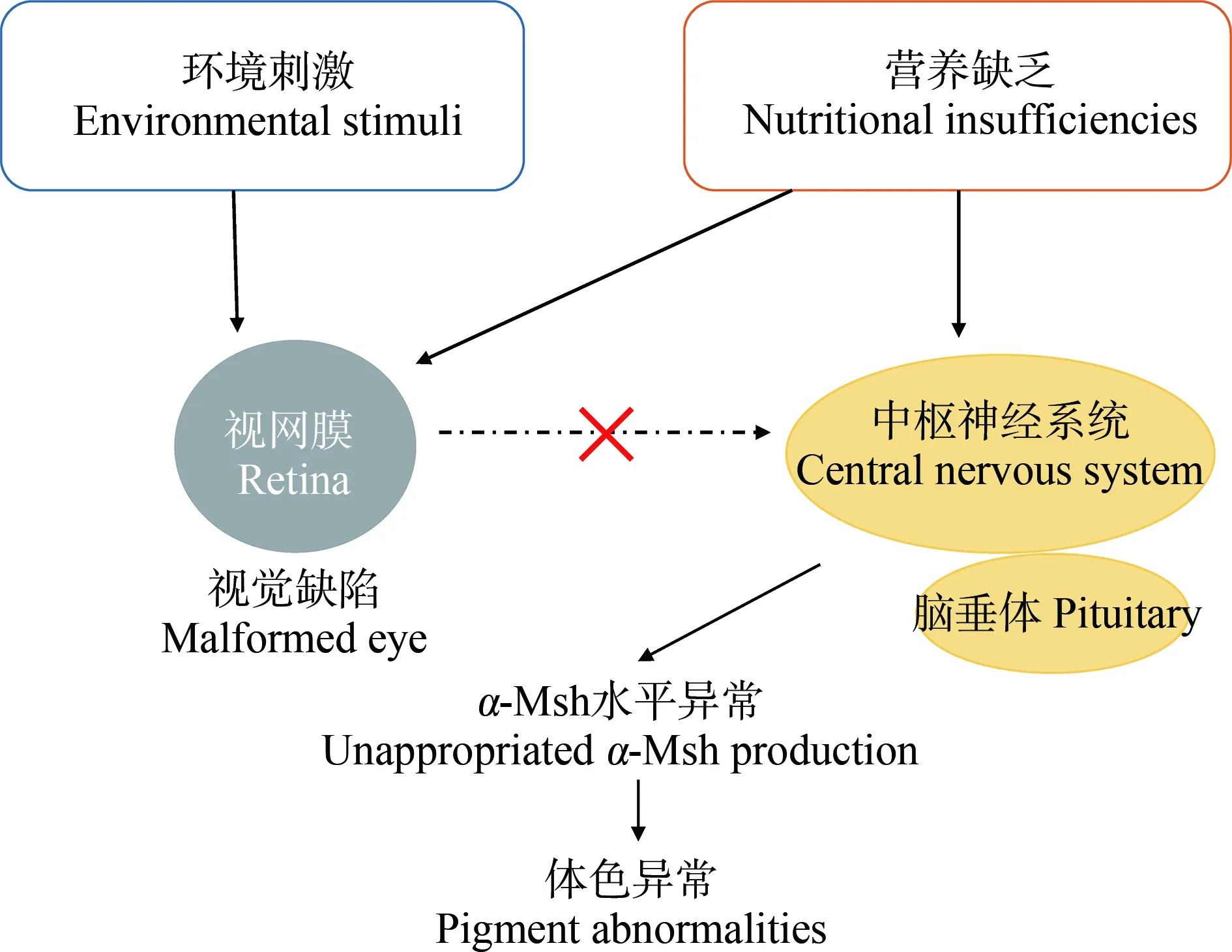
圖4 視覺缺陷誘導魚類體色異常機制圖Fig. 4 Schematic of the malpigmentation based on visual defects
魚類體色調控機制是維持魚類正常體色的前提,同時也是研究污染物對魚類體色毒理學機制的重要切入點。已經有研究表明,有機污染物(多氯聯苯、全氟烷氧烷和雙酚A等)能影響機體神經和內分泌系統、氧化應激和視網膜結構,這些變化均可能會干擾體色的調控[78]。相關研究亟待進一步的開展與完善。
4 污染物干擾魚類體色的機制研究(Research on the mechanisms of pollutants interfering with fish body coloration)
根據目前的研究,污染物干擾魚類體色的機制主要包括干擾神經和內分泌系統、影響色素合成或代謝、誘導氧化應激、影響視網膜結構與功能以及干擾色素細胞的分化、遷移與發育等。
環境內分泌干擾物可能主要通過干擾性激素水平影響體色,如17β-雌二醇(17β-estradiol, E2)、17α-乙炔基雌二醇(17α-ethinylestradiol, EE2)均能抑制孔雀魚與斑馬魚第二性特征,即交配體色,且這種影響似乎是不可逆的[6-8,79]。另外,金屬離子鎘污染也能夠誘導睪酮水平增加,增強體色,促進魚類的成功繁殖[80-81]。DDT同系物對非洲蟾蜍(Xenopustropicalis)和蘆葦蛙(Hyperoliusargus)內分泌系統產生負面影響,導致個體皮膚顏色的變化[82]。砷暴露會干擾魚類神經系統的分泌活動,影響色素細胞的結構和數量[83]。
污染物能夠干擾脊椎動物色素的合成或代謝。目前,在這方面的研究主要集中在黑色素和類胡蘿卜素。污染物能夠直接影響黑色素合成途徑,既可以促進黑色素的合成[84-85],也可以抑制黑色素的合成[86]。甲基汞能夠通過競爭性阻止銅輔助因子的結合來抑制酪氨酸酶的活性,從而導致黑色素的合成減少[87]。除草劑誘導機體內谷胱甘肽減少,而循環中的谷胱甘肽能夠抑制真黑色素的合成[4]。酸脅迫下,虹鱒體色較淺甚至變白、透明,主要通過抑制多巴色素氧化脫羧作用,抑制5,6-二羥基吲哚的合成,從而合成更少的黑色素,同時還能夠誘導黑色素合成相關酶的失活[22]。金屬污染(鉛和鎘等)能夠促進黑色素和類胡蘿卜素體色相關基因的表達[81,88]。基于類胡蘿卜素的體色不僅受重金屬的影響,而且受有機污染物的影響。有機磷殺蟲劑甲基對硫磷可以影響類胡蘿卜素的沉積,對胚胎期的阿馬里洛魚產生長期影響,甚至持續到成魚期[14]。
在我們的研究中,環境水平的三苯基錫(triphenyltin, TPT)抑制雄性孔雀魚的交配體色,并因此降低雄性求偶過程中的性吸引力[10]。一般認為睪酮水平的增加與雄魚交配體色的形成有關。有意思的是,TPT暴露后,雖然睪酮水平增加,雄性孔雀魚的體色卻表現出褪色,這提示TPT對體色的影響可能涉及其他機制。我們還發現TPT誘導了雄性孔雀魚產生氧化應激。根據“類胡蘿卜素權衡”假說,可以推斷,相對于裝飾體色,TPT暴露的雄性孔雀魚可能優先考慮氧化應激補償,進而導致皮膚中類胡蘿卜素含量的降低,體色減褪(圖5)。
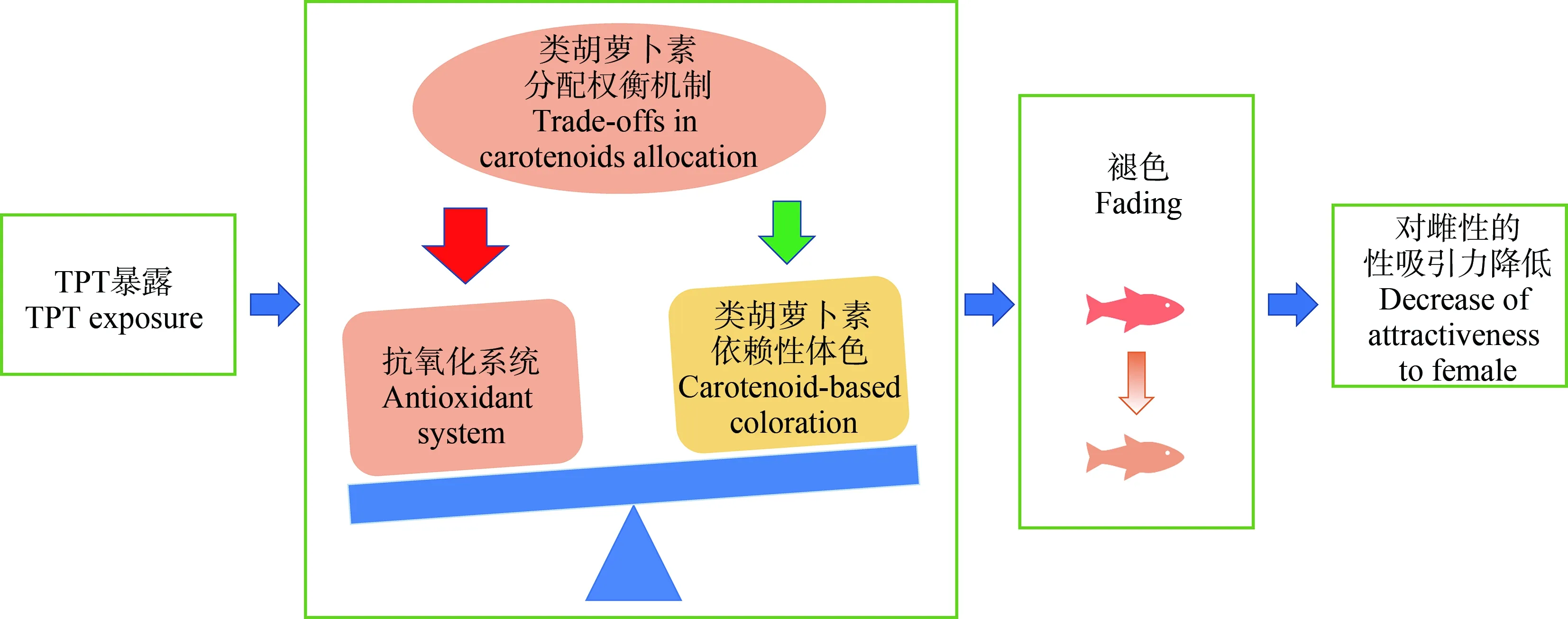
圖5 三苯基錫(TPT)干擾魚類體色的“類胡蘿卜素權衡”機制(仿Zhang等[10])Fig. 5 Fish exposed to triphenyltin (TPT) change their body coloration through the “carotenoid trade-off” hypothesis (Imitating Zhang et al[10])
另外,研究表明,TPT和雙酚S能夠破壞魚類視網膜結構,可能影響視網膜的光傳導,進而導致體色改變[11,89]。在胚胎發育期,TPT能夠引起非洲爪蟾胚胎黑色素的部分或完全喪失,降低黑色素細胞的正常樹突延伸,并破壞黑色素細胞的正常遷移[90];TPT能夠干擾斑馬魚幼魚黑色素分布,正常幼魚黑色素聚集成點狀斑,覆蓋背部小面積區域,而TPT暴露的幼魚黑色素則廣泛分布于背部[11]。
5 小結與展望(Summary and prospects)
污染物可通過影響發育、生理、行為、繁殖成功和生存,對魚類產生有害的影響,并可能最終影響魚類種群數量。通過物種間相互作用(獵物-捕食等),污染物可能會造成嚴重的生態危害[91]。目前,污染物對魚類體色分子機制的研究亟需深入開展,促進魚類體色基礎生物學的發展以及相關生物標志物的開發。在魚類體色形成的分子基礎研究上,利用基因組學和蛋白組學技術,我們可以借助表型/化學反應數據在一定程度上推斷污染物作用于魚類體色形成的靶點和毒理學機制。
污染物對魚類的影響均能夠通過體色的變化進行直觀地觀察和預測,這依賴于魚類體色較高的可塑性以及對外界環境的高敏感性。目前,魚類抗氧化酶、乙酰膽堿和卵黃蛋白原等作為水環境污染的早期預警生物標志物被廣泛關注。然而對魚類體色相關分子標志物的研究尚處于起步階段。有研究者曾提出魚類黑色素細胞可作為快速、可靠的水中重金屬污染物的生物標志物,鱗片作為水污染的指示物[23,83]。魚類包含脊椎動物中最多的色素細胞,體色涉及的基因數量在脊椎動物中也最為豐富,然而我們對魚類體色涉及的分子機制認識還遠遠不夠。因此,亟需深入研究魚類體色的分子基礎,有助于從龐大的體色相關基因庫中篩選具有應用前景的生物標志物,促進其在環境監測中的應用。
在將來的研究中,還需研究和建立代表性魚類的顏色基準,發展魚類體色檢測技術。另外,體色相關生物標志物的開發,不僅要以生理、發育等指標的變化為依據,而且還要從體色形成的分子層面探究相關基因或通路表達的變化,同時還需考慮多物種性和生物指標的多樣性,這樣才能使結果更具全面性、有效性和綜合性。隨著分子生物學與圖像視頻技術等不斷發展,將促進魚類體色及相關分子標志物在水環境污染監測中的應用。
