腐植酸對干旱脅迫下燕麥葉片碳同化酶活性及產量的影響
2021-09-23 08:41:08朱珊珊米俊珍趙寶平孫夢媛李英浩劉景輝
麥類作物學報 2021年6期
關鍵詞:產量
朱珊珊,米俊珍,趙寶平,孫夢媛,李英浩,劉景輝
(內蒙古農業大學農學院,內蒙古呼和浩特 010019)
燕麥是我國特色糧飼兼用型作物,具有營養成分高,抗逆性強等優點,在我國西北、華北、西南等干旱半干旱少雨地區種植面積較大,干旱是限制其生長發育和高產的重要因素[1]。干旱脅迫會抑制作物旗葉光合作用的酶活性[2],影響光合碳同化,導致作物減產[3]。嚴重干旱會導致植物葉片碳同化酶活力降低[4-5]。磷酸烯醇式丙酮酸羧化酶(PEPC)可通過植物內部釋放的二氧化碳和氮來提供碳骨架以進行生物合成,緩解干旱脅迫[6-7]。光合產物的積累量和作物產量與其葉片中甘油醛-3-磷酸脫氫酶(GAPD)和乙醇酸氧化酶(GO)活性有關[4,8]。核酮糖-1,5-二磷酸羧化酶(Rubisco)、GO、PEPC及GAPD是植物碳同化的關鍵酶,提高其效率和活力是促進植物光合作用的重要途徑[4,8-10]。腐植酸是存在于土壤、河泥和煤炭之中的天然有機混合物[11],有促進作物生長、節肥增效、增強作物抗逆性等功效,是一種多功能葉面肥[12]。張志芬[13]研究發現,噴施腐植酸可抑制燕麥葉片的氣孔開度,增加氣孔阻力,降低蒸騰作用,緩解燕麥干旱脅迫。腐植酸能夠緩解干旱對麻黃葉片中葉綠體片層結構的破壞,增加葉片的葉綠素含量和凈光合速率,顯著減輕由干旱脅迫造成的氧化損傷,提高麻黃的耐旱性,從而提高其產量[14]。梁太波[15]研究發現,腐植酸肥料可以提高旱作小麥旗葉實際光化學效率和光合電子傳遞速率,延緩衰老,提高水分利用效率。以上研究表明,干旱脅迫下,腐植酸類物質能通過改變作物葉綠素熒光參數和光合參數、降低電子傳輸通量、提高植物體內氣體交換率,顯著提高作物光合作用、抗旱能力以及產量[16-18]。但有關干旱脅迫下腐植酸對燕麥葉片的碳同化酶活性及產量影響的研究鮮有報道。本研究以燕科2號為材料,分析不同程度干旱脅迫下噴施腐植酸對燕麥不同生育時期葉片碳同化酶活性和產量的影響,以期為燕麥抗旱、增產措施的制定提供理論依據。
1 材料與方法
1.1 試驗材料及設計
試驗材料為燕科2號,由內蒙古農牧業科學院提供。腐植酸水溶肥料采購于內蒙古永業農豐生物技術有限責任公司,其水溶腐植酸含量≥50 g·L-1,N+P+K≥200 g·L-1,微量元素(錳、硼、鉬、鋅等)≥10 g·L-1。于2019年4月至8月在內蒙古農業大學燕麥產業研究中心溫室進行盆栽試驗。試驗選用直徑為23 cm、高18 cm的塑料盆進行,內裝混合土5 kg(河沙∶泥炭土體積比1∶1),泥炭土中N+P+K> 10 g·kg-1,有機質含量> 50 g·kg-1,pH值為5.5~8.5,土壤含水率為9%。播種前每盆施入2 g磷酸二銨(N 18%,P2O546%)做底肥。設置3個水分處理:正常供水(田間最大持水量的75%,M1)、中度干旱脅迫(田間最大持水量的45%,M2)和重度干旱脅迫(田間最大持水量的30%,M3);2個噴施處理:噴施500倍稀釋的腐植酸水溶肥料(HA)和等量清水(CK),共6個處理。采用隨機區組排列,每個處理重復3次,每重復種植10盆,每盆播種30株,三葉期定苗為20株。2019年4月14日播種,8月21日收獲。從拔節期(5月18日)開始控水至成熟期,控水方法采用稱重差值法[19]。分別在拔節期、抽穗期、灌漿期的上午 9:00-11:00進行HA和CK處理,在植株葉片均勻噴施至葉面濕透無滴水,每盆噴施量約14 mL;噴施7 d后取燕麥中上部葉片,投入液氮速凍后,放入-80℃冰箱冷藏,用于碳同化酶活性的測定。
1.2 測定項目與方法
1.2.1 碳同化酶測定
核酮糖-1,5-二磷酸羧化酶(Rubisco)、磷酸烯醇式丙酮酸羧化酶(PEPC)和乙醇酸氧化酶(GO)活性均采用蘇州科銘生物技術有限公司試劑盒測定,3-磷酸甘油醛脫氫酶(GAPD)活性采用上海繼錦化學科技有限公司試劑盒測定。
1.2.2 地上部干物質積累量和產量及其構成因素測定
地上部干物質積累量:分別于拔節、抽穗、灌漿期葉面噴施處理后7 d取樣,每個重復取10株,放入紙袋,烘箱105℃殺青30 min,80℃烘干到恒重。
產量及構成因素:燕麥成熟時收獲、考種,每個處理取5株, 測定主穗長、單株小穗數、穗粒數、穗粒重和千粒重,并測每盆的籽粒產量,重復3次。
1.3 數據統計分析
采用Excel 2010進行數據處理,用SPSS 22.0進行顯著性和相關性分析。
2 結果與分析
2.1 腐植酸對不同水分處理燕麥葉片Rubisco活性的影響
由圖1可見,從拔節期到灌漿期,不同水分處理燕麥葉片的Rubisco活性均呈先升高后降低趨勢。干旱脅迫導致葉片Rubisco活性顯著降低。M1水分條件下,HA處理拔節期、抽穗期和灌漿期燕麥葉片Rubisco活性較CK分別增加 5.62%、2.79%、6.88%,僅在灌漿期差異達到顯著水平。M2水分條件下,HA處理各時期葉片Rubisco活性較CK均顯著提高,拔節期、抽穗期和灌漿期分別增加了22.38%、25.83%、 19.17%。水分條件為M3下,拔節期、抽穗期和灌漿期HA較CK處理葉片Rubisco活性分別增加了28.45%、34.41%、32.22%,差異均達到顯著水平。以上結果說明,干旱脅迫顯著降低了燕麥葉片Rubisco活性;噴施腐植酸可有效緩解干旱脅迫對燕麥葉片Rubisco活性的影響,尤其在M3水分條件下效果更佳。
2.2 腐植酸對不同水分處理下燕麥葉片GAPD活性的影響
隨著干旱程度的加深,各時期葉片中的GAPD活性均呈降低趨勢(圖2)。與CK相比,M1水分條件下噴施腐植酸可以提高GAPD活性,但只在抽穗期效果顯著;在M2和M3水分條件下,噴施腐植酸均能使各時期GAPD活性顯著增加,其中在M3水條件下增加的幅度最大,拔節期、抽穗期和灌漿期分別增加36.55%、31.80%、39.60%。這說明噴施腐植酸可不同程度地減小干旱脅迫對燕麥葉片GAPD活性的影響,在M3處理下效果較好。
2.3 腐植酸對不同水分處理下燕麥葉片PEPC活性的影響
由圖3可知,燕麥葉片的PEPC活性在抽穗期達到最高。在M1水分條件下,噴施腐植酸后燕麥葉片PEPC活性與CK相比僅在拔節期差異顯著;當水分條件為M2和M3時,噴施腐植酸后葉片中的PEPC活性較CK在各時期均顯著增加,其中在M2水分條件下,拔節期、抽穗期和灌漿期噴施腐植酸較CK分別增加了21.43%、 24.04%、25.42%,M3水分條件下,拔節期、抽穗期和灌漿期噴施腐植酸較CK分別增加了 38.24%、32.96%、33.81%。這說明噴施腐植酸提高了干旱脅迫下燕麥葉片的PEPC活性,且M3水分條件下提升幅度最大。
2.4 腐植酸對不同水分處理下燕麥葉片GO活性的影響
從拔節期到灌漿期,燕麥葉片GO活性先降低后升高(圖4)。干旱脅迫導致燕麥葉片GO活性升高,且隨著干旱程度的加劇而增加。在M1水分條件下,噴施腐植酸后燕麥葉片GO活性與CK相比差異均未達到顯著水平;當水分條件為M2和M3時,噴施腐植酸后葉片GO活性在各時期與CK相比均顯著降低,其中M2條件下,拔節期、抽穗期和灌漿期較CK分別降低16.18%、13.64%、14.65%,M3條件下,拔節期、抽穗期和灌漿期分別降低16.85%、16.75%、18.39%。這表明干旱脅迫顯著提高了燕麥葉片 GO的活性,噴施腐植酸能有效緩解這種增長趨勢。
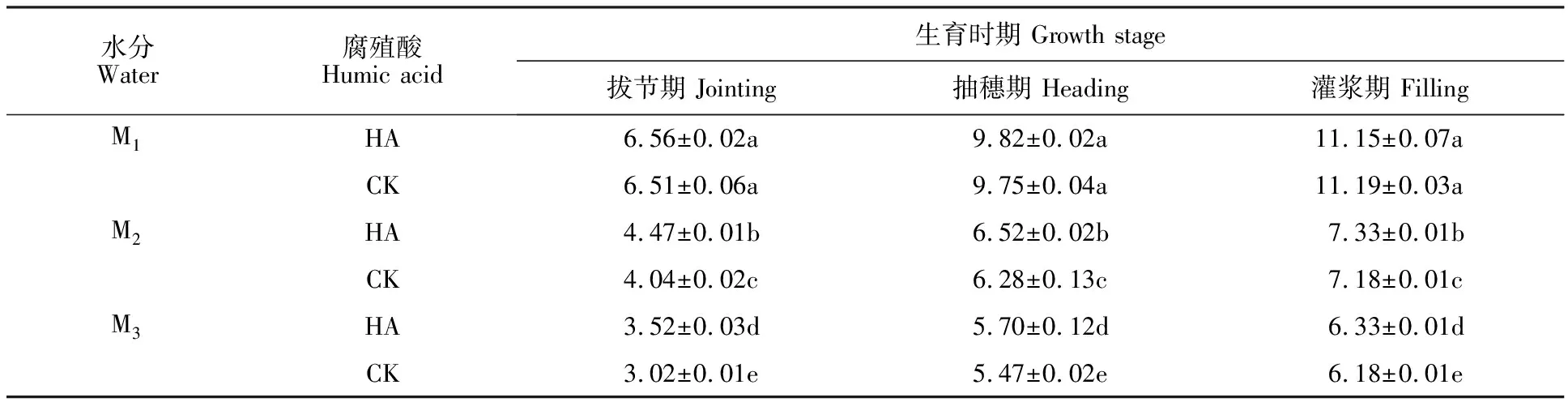
表1 不同處理下燕麥地上部干物質積累量的影響Table 1 Effect of different treatments on above-ground dry matter mass of oat g·10-1 plants
2.5 腐植酸對不同水分處理下燕麥地上部干物質積累量的影響
由表1可知,隨著干旱程度的加深,燕麥地上部干物質積累量顯著減少。與CK相比,在M1水分條件下,HA處理對燕麥各時期地上部干物質積累量沒有顯著影響;在M2和M3水分條件下,拔節期、抽穗期、灌漿期HA處理燕麥地上部干物質積累量均較CK顯著增加,增幅分別為10.64%、 3.82%、2.09%和16.56%、4.20%、 2.43%。
2.6 腐植酸對不同水分處理下燕麥產量及其構成因素的影響
表2顯示,干旱脅迫導致燕麥產量及其構成因素均顯著下降,且均表現為M1>M2>M3。與CK相比,噴施腐植酸能使燕麥產量增加,M1、M2、M3水分條件下分別增加2.63%、8.46%和10.26%,其中在M2和M3水分條件下差異達到顯著水平。從產量構成因素來看,噴施腐植酸后,穗長、穗粒重和千粒重較CK均增加,增加幅度均表現為M3>M2>M1,其中穗粒重和千粒重在M2和M3水分條件下增加顯著,分別增加 15.38%、16.07%和10.74%、15.67%;與CK相比,噴施腐植酸可以提高燕麥單株小穗數、穗粒數、穗數,但差異均不顯著。說明噴施腐植酸主要通過降低干旱對千粒重的影響而增加產量。
2.7 碳同化酶活性與產量及其構成因素的相關性分析
Rubisco活性與穗粒重和產量的相關性均達到極顯著水平,與千粒重呈顯著正相關,與穗粒重的相關性最大。PEPC和GAPD活性與穗粒重、千粒重和產量均呈極顯著正相關,與穗粒重的相關性最大。GO活性與穗粒重、千粒重和產量均極顯著負相關,與產量的相關性最大。穗粒重與千粒重與產量呈極顯著正相關(表3)。
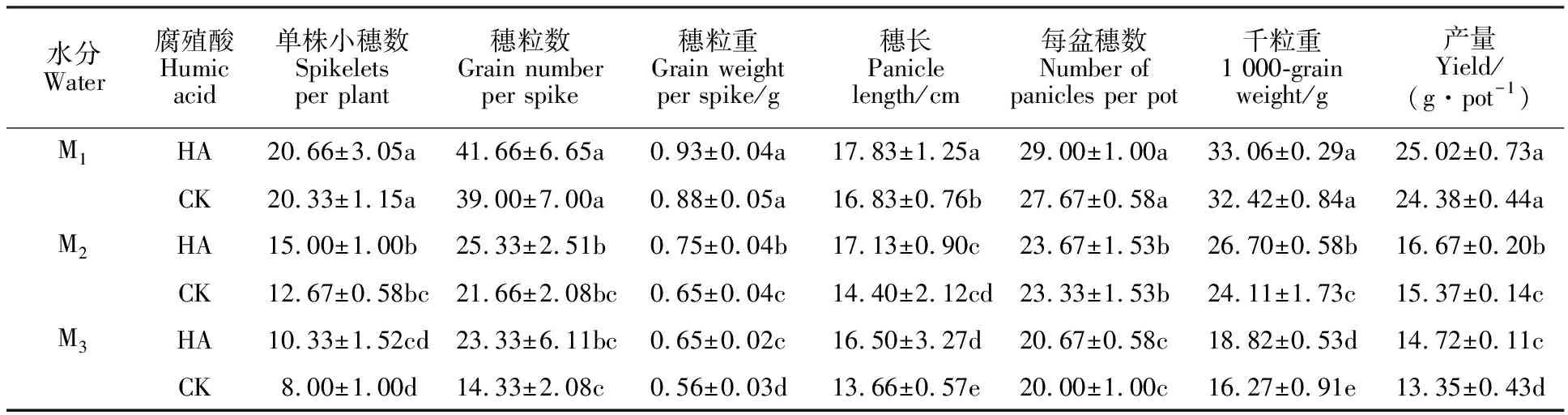
表2 不同處理對燕麥產量及其構成因素的影響Table 2 Effect of different treatments on oat yield and its components
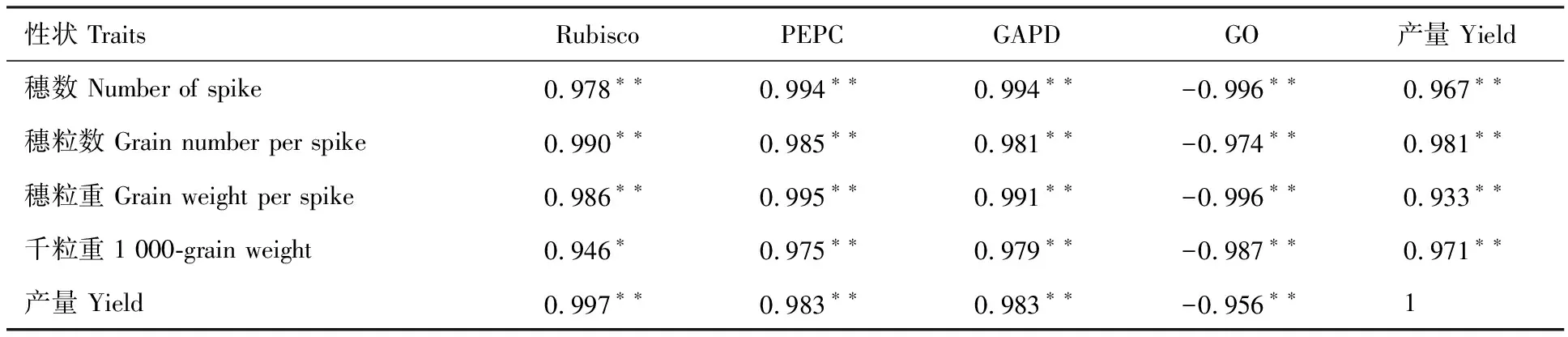
表3 燕麥葉片碳同化酶活性與產量及其構成因素的相關性分析Table 3 Correlation analysis of carbon assimilating enzyme activity in oat leaves with yield and its components
3 討 論
植物的光合作用是干物質積累和產量形成的基礎,較高的光合碳同化能力是獲得高產的前提。卡爾文循環中,Rubisco 是影響光合碳同化的關鍵酶,能使逆境脅迫中的植物維持光合作用[20]。干旱脅迫會導致 Rubisco 活性降低,光合作用產生限制[16,21-22],本研究結果與此相似,隨著干旱脅迫程度的增加,Rubisco活性逐漸下降,可能干旱脅迫使相應酶量降低,進而導致 Rubiso 活性下降,影響了光合碳同化[23]。在重度干旱脅迫中,作物葉片光合能力的降低主要受非氣孔因素限制,葉片光合放氧能力降低,光合磷酸化解偶聯,阻礙CO2擴散,RuBP羧化酶活性下降,光合器官的生理功能遭到破壞,影響作物光合作用[24-25],噴施腐植酸使植物在干旱脅迫下維持光合系統的穩定和相對較高的光合作用,對非氣孔因素引起的光合限制具有緩解作用[26-27]。有研究表明,在氮脅迫下添加腐植酸比未添加處理RuBPCase活性顯著提高[28]。Jannin等[29]提出,腐殖質處理后的植物Rubisco活性增加可能是由于細胞內葉綠體數量增加所致。本研究表明,重度干旱脅迫下,噴施腐植酸后燕麥葉片中Rubisco酶活性與CK相比提高幅度最大,中度脅迫條件下其活性雖有提高,但幅度較小。這可能是由于腐植酸在重度干旱脅迫下更能緩解干旱對葉綠體的傷害[13],增加RuBP羧化酶固定CO2的量,進而提高Rubisco的羧化效率,提高作物的光合能力。
GAPD是糖異生和糖酵解過程中的關鍵酶,催化3-磷酸甘油酸(3-PGA)還原成3-磷酸甘油醛,影響光合環的運轉效率及光合產物的積累[30]。Ziaf等[31]發現,干旱脅迫下GAPD基因的表達減輕了植物膜脂過氧化的程度。有研究表明,隨著水分脅迫時間的延長和脅迫強度的加劇,小麥旗葉的GAPD活性逐漸降低[23],與本研究結果一致。在逆境條件下,生物體中GAPD基因的轉錄及表達水平不穩定,大多數生物體都增加表達GAPD[32]。郭子平[33]研究表明,干旱脅迫下,小麥葉片總GAPD活性隨著時間的延長而增加。在本研究中,隨著干旱脅迫程度的加劇,葉片中的GAPD活性逐漸降低,與上述研究結果不同,可能是因為不同抗旱類型作物受到干旱脅迫時GAPD基因的表達調控因子和信號傳遞途徑存在差異,也可能是不同作物、不同品種的GAPD蛋白在植物細胞的失水耐受過程中作用不同[34]。有研究表明,腐植酸可以誘發含有NADP依賴性甘油醛-3-磷酸脫氫酶(GAPN)蛋白產生[35]。在本研究中,兩種干旱脅迫條件下噴施腐植酸,燕麥葉片的GAPD活性均能提高,但在重度脅迫下變化幅度較大。可能是因為腐植酸在植物細胞過度失水時誘導了GAPDH蛋白的大量表達,導致酶活性提高,促進光合產物的積累,最終增加燕麥的穗粒重,使產量增加。
PEPC在CAM和C4植物中的主要生理作用是初級CO2的固定還原[36]。有研究表明,C3植物中也存在PEPC[37],在大部分C3植物體內中,PEPC具有回補作用,補充三羧酸循環,其中間產物能被用于多種生物合成及氮同化消耗[36]。關于干旱脅迫對PEPC活性的影響,各試驗結果有所不同。有研究表明,在水分虧缺下,小麥PEPC活性明顯增強[18];而張 旭等[38]認為,干旱導致小麥旗葉PEPC活性下降。玉米葉片中PEPC活性在水分脅迫下先升后降[39]。在本研究中,隨著干旱脅迫程度的加劇,PEPC活性均逐漸降低,有可能是不同種類的植物應答干旱的生態適應性機制不同[40]。噴施腐植酸后,PEPC活性均有所提高,且土壤水分含量低時提升效果更佳,可能是在過度失水下,腐植酸在一定程度上更能誘導PEPC酶蛋白的表達,具體原因還需進一步探討。
GO在植物光呼吸代謝中具有關鍵作用,能提高植物光合效率[41]。隨著干旱脅迫的加劇,檸條中GO活性逐漸增強[42],與本研究研究結果相似。這說明隨著干旱程度加劇,乙醇酸氧化酶活性也隨之升高。乙醇酸氧化酶催化乙醇酸的氧化過程中伴隨著 H2O2的生成,是H2O2總數的70%[43]。水分脅迫能夠增加GO活性,誘導大量H2O2的累積[44]。Garcar等[45]研究發現,腐植酸可通過降低水稻H2O2的含量,提高膜的通透性,對干旱引起的氧化應激反應起到保護作用。本研究結果表明,腐植酸能使干旱條件下燕麥葉片GO活性降低,其中在重度干旱脅迫時,噴施腐植酸較CK處理相比降低幅度更大。這說明腐植酸可能是通過減少過氧化氫的生成來抑制乙醇酸氧化酶的活性,降低燕麥的光呼吸,增加作物的光合作用。
綜上,干旱脅迫通過改變碳同化酶活性,降低光合碳同化能力,從而影響燕麥的干物質積累和產量形成,噴施腐植酸能顯著提高燕麥葉片Rubisco、GAPD和PEPC活性,提高碳的同化能力,降低GO活性,促進光合作用,增加產量,當田間最大持水量為30%提升幅度更大。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
