冷季型草坪草耐熱性研究進展
2021-09-22 08:56:26譚真真張夏香楊志民
草業學報 2021年9期
譚真真,張夏香,楊志民
(南京農業大學草業學院,江蘇 南京210095)
草坪草根據其對氣候的適應性,一般分為冷季型草坪草和暖季型草坪草兩種類型[1-2]。常見的冷季型草坪草有:高羊茅(Festuca arundinacea)、草地早熟禾(Poa pratensis)、多年生黑麥草(Lolium perenne)、細羊茅(Festuca rubra,Festuca ovina)和匍匐剪股穎(Agrostis stolonifera)等[3]。
冷季型草坪草主要分布在溫帶冷涼地區,地上莖葉最佳生長溫度為18~24℃,根系最佳生長溫度為10~18℃[2],當地上溫度高于30℃時,會出現枯黃、甚至死亡現象[4]。近年來,隨著草坪育種技術發展,國內外培育了大量耐熱的草坪草品種,其使用范圍也向長江流域過渡帶南部推進[5]。冷季型草坪草耐低溫且綠期長,目前已經成為長江流域主要建植草種[6]。但是冷季型草坪草耐熱性差,即使較耐熱的高羊茅,其品種之間的耐熱性也有較大差異[5]。另一方面,根據聯合國政府間氣候變化專門委員會(Intergovernmental Panel on Climate Change,IPCC)2018年發布的報告顯示,目前全球溫度比工業化前升高了1.5℃[7]。在全球氣候變暖背景下,我國近幾十年來極端高溫氣象事件頻發。如2003年,我國南方地區遭受大范圍熱浪襲擊,特別是江南和華南地區出現持續1個多月的高溫天氣,長江中下游至華南北部地區夏季最高氣溫達38~40℃[8],冷季型草坪草很難在長江流域氣候過渡區越夏,嚴重限制了其推廣應用。為了充分利用優良冷季型草種質資源,國內外學者圍繞冷季型草坪草耐高溫問題進行了大量研究。本研究將圍繞冷季型草坪草響應高溫脅迫的生理和分子機制方面,綜述近年來主要的研究進展,以期為冷季型草坪草耐熱性研究提供理論參考。
1 冷季型草坪草響應高溫脅迫的生理機制
植物受到高溫等非生物脅迫時會發生一系列生理生化反應,植物高溫耐性往往是各種應激反應機制協同作用的結果,以防止細胞受損和重建細胞內穩態[9]。在生理水平上,隨著溫度增高,植物會發生一系列生理代謝變化響應熱脅迫,主要表現為:碳代謝、抗氧化代謝和激素代謝變化。
1.1 碳代謝
高溫影響植物的碳代謝,包括植物通過光合作用固定的碳和通過呼吸作用消耗的碳[3]。高溫會抑制冷季型草坪草的光合作用,同時會加快植株的呼吸作用。長時間的熱脅迫會造成植株碳耗竭或饑餓,從而抑制其生長,這是限制冷季型草坪草生長的主要因素之一[10-13]。因此,了解植物碳代謝和碳利用機制對于提高草坪草耐熱性至關重要。
光合作用是植物最重要的代謝過程之一,作為C3光合途徑的冷季型草坪草對溫度升高尤為敏感。在熱脅迫下,各種光合特性常被作為植物耐熱性的指標,因為任何限制光合作用特性的因素都會限制植物在高溫下的生長[14]。高溫脅迫下,植物氣孔關閉,葉綠素含量減少,參與光合作用的酶活性降低,光過量引起光抑制進而綜合影響植物的光合速率[3]。
高溫脅迫會打破植物水分代謝平衡,導致葉片失水,根系吸水能力下降,從而導致氣孔關閉[15]。高溫脅迫對植物類囊體和光合功能的損害主要表現在葉綠素含量下降[14-15]。葉綠素熒光量子產量反映了光合作用光反應的功能,被廣泛用作高溫抑制光合作用的指標[16]。高溫脅迫下,早熟禾屬(Poaspp.)、黑麥草屬(Loliumspp.)、羊茅屬(Festucaspp.)和剪股穎屬(Agrostisspp.)等C3冷季型草坪草凈光合速率、葉綠素含量和葉綠素熒光量子產量呈下降趨勢[17-19]。高溫脅迫下葉片葉綠素含量下降的主要原因是熱脅迫加速了葉綠素的降解。熱脅迫下,葉綠素降解酶(包括葉綠素酶和葉綠素降解過氧化物酶)的活性顯著升高[20]。
核酮糖-1,5-二磷酸羧化酶/加氧酶(ribulose bisphosphate carboxylase oxygenase,Rubisco)是C3光合途徑和光呼吸中的關鍵酶。熱脅迫抑制C3植物光合作用與Rubisco的失活密切相關[21]。雖然Rubisco的催化活性在一定范圍內隨著溫度的升高而增加,但該酶對CO2的低親和力及其在高溫下作為氧合酶的雙重特性限制了C3植物在高溫下的凈光合速率[22]。溫度持續升高降低了Rubisco對C3草坪草中CO2的親和力,主要由于Rubisco和Rubisco結合蛋白的大小亞基減少,并抑制了Rubisco激活酶活性[22-23]。Yu等[24]研究表明,提高高羊茅生長環境中的CO2濃度可以增強其耐熱性。Liu等[25]研究表明,與光合色素含量或光化學效率的變化相比,Rubisco活性在熱脅迫下受到抑制是導致冷季型草坪草的光合作用下降的主要原因。而Chen等[26]認為高羊茅耐熱性可能與其保持較低的光化學活性有關。
大多數植物的呼吸作用隨溫度的升高而增強。Huang等[27]研究表明,在持續高溫脅迫下,匍匐剪股穎每日碳消耗量與產量比急劇上升,當溫度升高到34~38℃時,其呼吸速率超過光合速率。與耐熱品種相比,熱敏感匍匐剪股穎品種具有更高的呼吸與光合作用比[27]。長時間的熱脅迫,會導致植物碳耗竭或饑餓,從而限制草坪草生長。
1.2 抗氧化代謝
在熱脅迫下,光合作用中的碳同化生化反應速率比光的吸收和電子的傳輸速率要慢,這就會導致氧脅迫。由熱脅迫引起的氧脅迫,其結果就會產生大量的活性氧(reactive oxygen species,ROS),對植物細胞造成損傷。已有研究表明,氧脅迫會導致匍匐剪股穎[28]、草地早熟禾[18]脂質過氧化,膜穩定性降低。同時,植物細胞也有一套清除自由氧的機制,通過抗氧化酶系統或積累抗氧化化合物來保護細胞免受ROS的攻擊[29]。
植物細胞中的抗氧化酶主要有:超氧化物歧化酶(superoxide dismutase,SOD)、抗壞血酸過氧化物酶(ascorbate peroxidase,APX)、過氧化物酶(peroxidase,POD)、過氧化氫酶(catalase,CAT)、脫氫抗壞血酸還原酶(dehydroascorbate reductase,DHAR)、單脫氫抗壞血酸還原酶(monodehydroascorbate reductase,MDAR)、谷胱甘肽過氧化物酶(glutathione peroxidase,GPX)、谷胱甘肽還原酶(glutathione reductase,GR)、氧化型谷胱甘肽(glutathione oxidized,GSSG)和單脫氫抗壞血酸(monodehydroascorbic acid,MDHA)等。但是,細胞中的抗氧化酶活性一般會隨著溫度的進一步升高而降低,從而減弱了抗氧化酶系統的保護作用。Du等[30]研究表明,暖季型草坪草(C4)比冷季型草坪草(C3)具有更強的耐熱性,主要原因是前者具有更強的氧化清除能力,表現在較高的SOD和POD活性上。Jiang等[18]研究表明熱脅迫會使高羊茅和草地早熟禾APX、CAT和GR活性降低,也會降低匍匐剪股穎根和葉中的SOD和CAT活性,加速了細胞脂質過氧化水平和葉片衰老[28]。同一冷季型草坪草的不同品種的耐熱性也可從抗氧化酶活性體現。研究表明,熱脅迫下的草地早熟禾、匍匐剪股穎耐熱品種比不耐熱品種具有更高的SOD活性、較低的脂質過氧化水平和較高的膜穩定性[31-33]。
植物細胞中的抗氧化化合物主要有:谷胱甘肽(glutathione,GSH)、L-抗壞血酸(L-ascorbic acid,Asc)、α-生育酚(維生素E)、類胡蘿卜素、玉米黃素和多胺等。抗氧化物可以保護細胞免受由熱脅迫引起的活性氧對細胞的毒害作用,維持細胞內穩態[34]。內源積累或外源施用抗氧化物可以提高植物抵御非生物脅迫的能力[35]。Zhang等[36]的試驗中,高溫脅迫下外施亞精胺(spermidine)顯著降低了高羊茅電解質滲漏率以及H2O2和O2-含量,并提高了細胞抗氧化酶活性及葉綠素含量。Chen等[37]研究表明,外施Asc可以通過降低ROS含量及抑制光能量的輸入來減輕熱脅迫對高羊茅的損害。
1.3 激素代謝
植物激素是指植物體內產生的一些微量而能調節自身生理過程的有機化合物。植物激素在植物生長發育過程中起著調控樞紐作用,它可以連接、傳遞并重新編譯復雜信號以應對外界脅迫。各類植物激素存在復雜的交叉反應,既相互獨立又相互作用,共同調控植物生長發育及環境適應的各個過程[38]。目前熟知的植物激素主要有5大類:生長素(auxin)、赤霉 素(gibberellins,GAs)、細 胞 分 裂 素(cytokinins,CTK)、脫落酸(abscisic acid,ABA)和乙烯(ethylene,ETH)。除此之外,如油菜素甾醇(brassinosteroids,BRs)、茉莉酸(jasmonic acid,JA)、水楊酸(salicylic acid,SA)、褪黑素(melatonine,MT)和獨腳金內酯(strigolactones,SLs)等植物激素陸續被發現并應用。在冷季型草坪草熱脅迫研究上,以上激素都有相關報道(表1),本研究將著重闡述CTK、GA、ABA和MT這4類在冷季型草坪草耐熱性方面研究相對較多的激素。
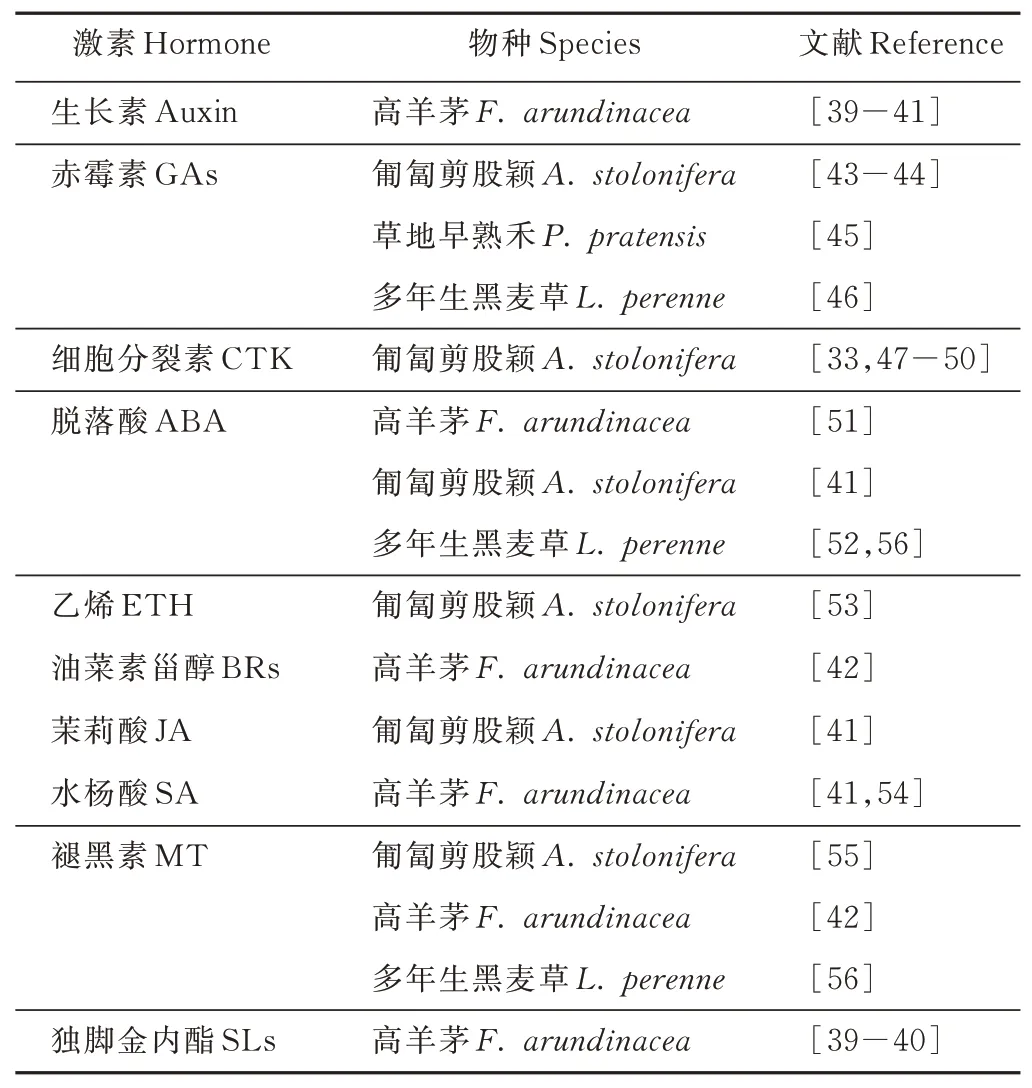
表1 部分冷季型草坪草耐熱性與激素研究Table 1 Partially research on heat-resistance and hormone of cool-season turfgrasses
1.3.1 CTK CTK最先在植物根部合成,調控植物的各種發育過程,包括細胞分裂、葉綠體合成、葉片衰老和抗氧化系統等[3]。熱脅迫會導致匍匐剪股穎異戊烯基腺嘌呤(isopentenyladenine,iPA)、玉米素/玉米素核苷(zeatin/zeatin riboside,Z/ZR)和二羥基玉米素/二羥基玉米素核糖(dihydroxyzeatin/dihydroxyzeatin riboside,DHZ/DHZR)含量下降[47-48]。熱脅迫下CTK含量下降會加速葉片衰老,而耐熱的匍匐剪股穎品種在相同處理條件下,具有較高的CTK含量,葉黃化程度低,草坪質量高[33]。Liu等[48]研究表明,在匍匐剪股穎根區施用ZR可以提高內源CTK的含量,緩解熱脅迫對植株地上和地下部分的傷害,其耐熱性提高的機理可能與提高抗氧化酶(如SOD和CAT)的活性,以緩解細胞膜的脂質過氧化作用相關。Veerasamy等[50]研究表明,葉面噴施ZR能提高匍匐剪股穎可溶性蛋白質含量,并使熱激蛋白(heat shock proteins,HSPs)(HSP32和HSP57)上調表達。
1.3.2 GA GA參與調控植物生長發育,如葉片膨脹、莖伸長和開花等環節[57]。然而,對于追求低矮平整的草坪來說,莖葉伸長并不是一個好的特性。因此,在草坪草中應用最多的是使用GA抑制劑—抗倒酯(trinexapacethyl,TE)來控制草坪高度[44]。相關研究表明,TE能提高草坪草應對各種非生物脅迫,如干旱、寒冷、高溫和遮陰等的耐性,提高草坪質量[45-46,58-60]。研究表明,應用TE提高了匍匐剪股穎[58]、草地早熟禾[45]和多年生黑麥草[46]的葉綠素含量和莖葉密度。McCann等[44]研究表明,熱脅迫下,經葉面噴施TE的匍匐剪股穎具有較高的葉綠素含量、光化學效率、葉片相對含水量,推測可能與TE提高了植物光合能力和水分利用效率有關。
1.3.3 ABA ABA在植物應對干旱和高溫等非生物脅迫中起著重要的調控作用[61-63]。逆境脅迫會促使植物體內ABA含量迅速增加[64],而ABA的積累也會提高植物耐熱性[65],因為ABA參與氣孔調節和抗氧化防御系統的激活。已有研究表明,外源噴施ABA可以提高匍匐剪股穎的耐熱性[53]。Zhang等[52]對ABA提高高羊茅耐熱性及其機理的研究表明,外源噴施ABA顯著提高了高羊茅植株的耐熱性,在熱脅迫下高羊茅葉片相對含水量、光化學效率和膜穩定性增加;ABA在干旱鍛煉誘導的耐熱性可能與脅迫響應及ABA信號通路相關基因(CDPK3、MPK3、DREB2A、AREB3、MYB2、MYC4)、熱激蛋白(HSP18、HSP70)及熱激轉錄因子HsfA2等上調表達相關。
1.3.4 MT MT被認為是一種積極的植物信號分子和強有力的抗氧化劑,它能減輕各種非生物脅迫對植物造成的損害,在冷季型草坪草耐熱性研究方面,已經有相關報道。Ihtisham等[54]研究表明,高溫脅迫下,用MT處理高羊茅幼苗,其活性氧(ROS)、電解質滲漏率和丙二醛含量均降低,而葉綠素含量、總蛋白含量和抗氧化酶活性升高。Merewitz等[55]研究表明,經MT處理過的匍匐剪股穎植株葉片在熱脅迫下具有更高的SOD和POD活性,并增加了不飽和脂肪酸、亞油酸的含量。Zhang等[56]研究表明,葉面噴施MT明顯減緩了由熱脅迫引起的多年生黑麥草葉片衰老。經MT處理的多年生黑麥草在熱脅迫下植株分蘗數、地上干重、株高、草坪質量、葉綠素含量、光化學效率、凈光合速率和細胞膜穩定性等生理指標數值明顯高于對照組;另外,在轉錄水平上,經MT處理后的多年生黑麥草葉片衰老相關基因(LpSAG12.1和Lph36)下調表達,內源MT和CTK含量增加,但ABA含量減少,推測MT可能參與激活CTK合成和信號并抑制ABA合成進而調控黑麥草對高溫的耐性。
總體來講,冷季型草坪草耐熱性相關激素研究在近十幾年中已取得一定進展。在激素類型上,各類激素幾乎都有涉及,包括一些新型植物激素;在研究深度上,也由簡單的激素和熱脅迫處理后生理指標測定向轉錄組、蛋白組及代謝組方向挖掘;在研究對象上,主要集中在匍匐剪股穎、高羊茅和多年生黑麥草這3種冷季型草坪草上。但相比模式植物和主要農作物,在研究深度和研究手段上的差距還非常明顯。因此,可以借鑒模式植物及主要農作物研究思路、研究方法和成果應用于草坪草耐熱性研究中。
2 冷季型草坪草響應高溫脅迫的分子機制
在分子水平上,當植物受到熱脅迫時,會激活特定的基因,促使一些特定的蛋白質被翻譯或促進某些代謝物合成,最終減輕或緩解熱脅迫對植物造成的傷害。
2.1 相關耐熱基因
目前已經報道的與冷季型草坪草耐熱相關基因主要有2類:熱激蛋白(HSPs)和熱激轉錄因子(heat shock factors,HSFs)。HSPs在高溫等非生物脅迫下能被大量誘導表達,而HSPs的轉錄受HSFs的調控[66],HSFs和HSPs在植物的熱應激和獲得性耐熱中起著重要調控作用[67]。
2.1.1熱激蛋白 熱脅迫下,大多數蛋白質降解的速率會超過其合成速率[68],但研究表明,HSPs在高溫、干旱、過氧化和重金屬等脅迫下能被大量誘導表達[69],主要通過參與蛋白合成、折疊、細胞定位及蛋白降解等多種生物功能來維持植物內穩態[70]。因此,HSPs在植物抵御逆境方面發揮著重要作用[71]。Mian等[72]將HSPs分為3大類:LMW-HSP(15-30 kDa)、HSP70(69-71 kDa)和HMW-HSP(80-114 kDa),Kotak等[73]將HSPs家 族 分 為HSP100、HSP90、HSP70、HSP60和小HSP(sHSPs)5大類。
目 前 已 報 道 的 與 冷 季 型 草 坪 草 耐 熱 性 相 關 的 熱 激 蛋 白 有HSP90[74-75]、HSP70[74,76]、sHSPs(HSP17[77]、HSP17.8[76-78]、HSP17.9[78]、HSP18[74]、HSP26[79]、HSP26.2[80]和HSP26.7[76,80]、HSP26.8[81])等,其中大部分HSPs在草坪草耐高溫過程中起到積極作用,而部分如HSP17和HSP26.8負調控植物的耐熱性。Wang等[74]研究認為,葉片噴施磷脂酸(phosphatidic acid,PA)和CaCl2處理的高羊茅表現出較好的耐熱性可能與PA和Ca2+參與介導HSFs和HSPs表達有關,短期熱脅迫會誘導激活FaHsfA2c和FaHSP18,而長期熱脅迫會誘導FaHSP70和FaHSP90表達。Kim等[79]從水稻中克隆了OsHSP26基因,經農桿菌轉化高羊茅,與野生型相比,轉基因高羊茅植株耐熱性顯著提高,推測可能是OsHSP26在熱脅迫和氧化脅迫過程中對PSII起重要保護作用。畢傲月[78]從高羊茅中克隆的FaHSP17.8和FaHSP17.9基因在34℃(4 h)和22℃(20 h)高溫循環脅迫模式下,表現出表達量顯著高于第一次高溫脅迫的現象,認為FaHSP17.8和FaHSP17.9具有耐熱記憶效應。Wang等[80]研究表明,相比熱敏感型匍匐剪股穎,LMW-HSP基因如ApHSP26.2和ApHSP26.7在耐熱的匍匐剪股穎中高度積累。Hu等[66]對耐熱基因型PI574522和熱敏感基因型PI512315高羊茅進行高溫預馴化處理,結果表明:LMW-HSP和HMW-HSP在隨后的熱脅迫中表現出轉錄記憶。此轉錄記憶可以持續4 d,并可誘導高羊茅產生較高的耐熱性,具體表現在較高的草坪質量、較低的電解質滲透率和較高的葉片代謝物質含量上。Sun等[77]從匍匐剪股穎中分別克隆了AsHSP17和AsHSP26.8a[81]基因,并在擬南芥(Arabidopsis thaliana)中過表達,結果表明轉基因擬南芥對熱脅迫的耐受性降低,并對ABA和鹽脅迫處理更敏感,推測AsHSP17和AsHSP26.8a可能通過調控ABA和其他信號通路而負調控植物應對各種非生物脅迫。
2.1.2 熱激轉錄因子 HSFs是一類熱激轉錄因子,熱脅迫下通過與熱激元件結合激活下游相關基因表達而進一步影響植物代謝[82]。HSF家族可以分為3大類:HsfA、HsfB和HsfC。低溫、高溫、澇漬、鹽、滲透和氧化等非生物脅迫均會誘導HSF家族成員表達[83-85],但其表達模式隨脅迫種類、脅迫強度和脅迫時間的不同而不同。在冷季型草坪草耐熱性方面,研究較多的是HsfA2家族。如Wang等[86]對高溫和低溫處理10 h后的高羊茅和多年生黑麥草葉片進行轉錄組分析,結果表明:HSFs對溫度脅迫響應強烈,HsfAs在熱脅迫下上調表達,HsfBs在高溫和低溫脅迫下均上調表達,而HsfCs主要在低溫脅迫下上調表達;在3類HSFs家族中,HsfA2的表達量最高。Wang等[65]研究表明高羊茅FaHsfA2c在ABA介導的植物耐熱性中起重要作用,在外源ABA處理的植物中,FaHsfA2c、FaHSPs和ABA應答轉錄因子的表達水平上調,ABA應答轉錄因子可能參與調節FaHsfA2c和下游FaHSP的轉錄活性,從而提高高羊茅耐熱性。但Zhuang等[87]從高羊茅中克隆HsfC基因(FaHsfC1b)并在擬南芥中過表達,結果表明,在48 h處理過程中,熱、滲透、鹽和冷脅迫均會影響FaHsfC1b表達。高溫脅迫下,在擬南芥中過表達FaHsfC1b,植株存活率、葉綠素含量和光化學效率提高,而電解質滲透率、H2O2和O2·-含量下降,并誘導內源AtHsfC1基因表達,熱保護蛋白編碼基因(AtHSPs、AtGalSyn1、AtRof1、AtHSA32)和ABA合成酶編碼基因(AtNCED3)的表達水平也顯著升高。
2.2 轉錄分析
近年來,隨著轉錄組測序技術高速發展,轉錄組學技術在草坪草研究中逐漸增多。利用此技術可以實現從轉錄水平揭示逆境脅迫下草坪草整個基因組水平的表達特征,并針對不同種類草坪草篩選不同逆境、組織、器官和生長發育階段的差異表達基因。另一方面,挖掘和發現草坪草抵御逆境脅迫的關鍵功能基因對于探究草坪草響應逆境脅迫的分子機制具有重要意義。
2007年,Sathish等[88]就 利 用 基 于Sanger測 序 法 的 基 因 表 達 系 列 分 析(serial analysis of gene expression method,SAGE)對多年生黑麥草轉錄組進行了測序,后面陸續有Studer等[89]、Farrell等[90]利用不同測序方法對其進行了測序。最新結果表明,多年生黑麥草在熱脅迫下鑒定出的差異表達序列與相對常見的熱脅迫響應基因一致,主要包括HSFs、HSPs和抗氧化相關基因等[91]。
Hu等[92]對冷季型草坪草高羊茅高溫脅迫下轉錄組分析,共獲得2.62億個高質量轉錄本,并成功組裝成31803個平均長度為1840 bp的非冗余序列,其中有12974個序列對熱脅迫表現出不同的表達模式。FaHSP20在耐熱和熱敏感兩種基因型的高羊茅中均會激活,而FaHSP90只在耐熱基因型的高羊茅中激活。Alam等[42]用MT和表油菜素內酯(24-epibrassinolide,EBL)處理的高羊茅幼苗轉錄組學分析表明,熱處理2和12 h后,分別有4311和8395個轉錄序列發生了顯著變化。其中,熱處理后涉及熱應激反應的DNA、RNA和蛋白質降解、氧化還原、能量代謝和激素代謝途徑均被高度誘導。MT和EBL處理使包括FaHSFA3、FaAWPM和FaCYTC2等基因顯著上調,表明這些基因可能作為MT和EBL的靶基因參與高溫脅迫響應。Xu等[93]對熱敏感剪股穎(A.stolonifera)和耐熱剪股穎(A.scabra)熱脅迫下RNA-seq轉錄譜分析表明:分別有1393個(675個上調和718個下調)和1508個(777個上調和731個下調)差異表達的基因。與熱敏感剪股穎相比,耐熱剪股穎具有較高的耐熱性與氧化保護、脯氨酸生物合成、脂類水解、半纖維素和木質素生物合成等基因的上調有關。Qian等[94]采用單分子實時測序和Illumina測序相結合的方法,獲得了高羊茅熱處理1和72 h的轉錄組。短期和長期熱脅迫分別誘導了4076和6917個差異表達基因(differentially expressed genes,DEGs)。對其DEGs進行生物信息學分析表明,短期熱脅迫能強烈激活HSPs和HSFs表達,并特異性激活編碼谷胱甘肽S-轉移酶、鈣信號傳遞相關基因、光合作用相關基因和植物激素信號通路相關基因表達。而長期熱脅迫使葉綠素分解代謝基因、植物激素合成/降解基因、脅迫相關基因和植物特異性轉錄因子(NACs)上調表達,光合作用相關基因和過氧化氫酶編碼基因下調表達。在煙草(Nicotiana tabacum)中的瞬時過表達FaHsfA2a,表明其可以緩解熱脅迫引起的細胞膜損傷,但瞬時表達FaNAC029和FaNAM-B1加重了細胞膜損傷。
隨著生物技術進一步發展,冷季型草坪草耐熱性相關基因研究取得長足發展,一些響應熱脅迫的熱激轉錄因子及相關蛋白基因不斷被克隆和發掘,基因功能也逐漸明晰,但目前對于熱脅迫誘導的一系列基因表達的調控機制尚不清楚,特別是植物耐熱性受多基因控制時,單個基因的克隆或轉化并不能說明其整個調控分子機制,因此還需要進一步研究。
3 問題及展望
在全球氣候變暖的大背景下,研究冷季型草坪草抗熱性具有重要意義。近年來,隨著對冷季型草坪草抗熱性研究不斷深入,其抗熱生理和分子機制也進一步明晰,特別是植物激素代謝機制、耐熱基因挖掘及蛋白功能等研究,對于選育耐熱草坪草品種、提高草坪養護管理水平提供了重要的理論支持。
但冷季型草坪草耐熱性研究也存在諸多問題,主要表現在:1)草坪草耐熱生理機理仍然不十分清楚。目前雖明晰了一些離子轉運代謝途徑、抗氧化防御系統及轉錄因子等作用機制,發掘了一些耐熱相關的基因與蛋白,但研究并不十分深入和系統。基于系統的蛋白組、代謝組和轉錄組方面的冷季型草坪草抗熱性研究遠遠滯后于其他模式植物和主要作物研究。2)缺乏廣泛的田間試驗研究。目前對冷季型草坪草耐熱性研究試驗基本都是在人工氣候室或培養箱進行熱脅迫處理后,測量相關生理代謝組、蛋白組或轉錄組數據后分析其是否抗熱,很少有田間試驗數據補充佐證。
因此,結合最新的植物代謝組學技術,通過改變傳導信號或代謝等途徑提高冷季型草坪草耐熱性,并借鑒模式植物和作物研究中最新成果,如綜合利用高通量測序、蛋白質組學、基因編輯等方法和技術定向挖掘與耐熱性相關的基因,提高冷季型草坪草耐熱性,對于冷季型草坪草理論研究和產業發展具有重要意義。另外,補充田間試驗研究,能真正評估目的基因、蛋白或轉基因植物是否具有耐熱性。因為在實際生產中,草坪草耐熱性還受環境和管理技術等多方面因素的影響,因此,在已有研究基礎上補充田間試驗這對于將來應用推廣具有重要意義。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
