黃河源區(qū)建植19年人工草地生物結(jié)皮CO2通量與葉綠素?zé)晒鈪?shù)的變化
2021-09-17 10:10:38孫華方李希來(lái)李成一林春英金立群楊鑫光
草地學(xué)報(bào) 2021年5期
孫華方,李希來(lái)*,李成一,張 靜, 林春英,2,金立群,3,楊鑫光
(1.青海大學(xué)農(nóng)牧學(xué)院,青海 西寧 810016;2.青海省人工影響天氣辦公室,青海 西寧 810001;3.湟源縣草原站,青海 西寧 812100;4.青海民族大學(xué)生態(tài)環(huán)境與資源學(xué)院,青海 西寧 810007)
生物土壤結(jié)皮(Biological soil crusts,BSCs)又稱(chēng)生物結(jié)皮,是藍(lán)藻、地衣、苔蘚、真菌和細(xì)菌組成的復(fù)雜群落,這些結(jié)殼約占地球陸地面積的40%[1-2],被認(rèn)為是世界上熱、冷、干旱和半干旱地區(qū)的重要地表組成部分[3-5]。近年來(lái),BSCs因具有較高的碳循環(huán)效率受到學(xué)者的廣泛關(guān)注[6-8]。在干旱和半干旱地區(qū),BSCs在生態(tài)系統(tǒng)碳固定和釋放過(guò)程中起著關(guān)鍵作用。研究發(fā)現(xiàn),在半干旱草原生態(tài)系統(tǒng)中,BSCs作為主要的碳源,所釋放的碳占呼吸釋放總量的43%[7],而在沙漠地區(qū),BSCs固碳效果顯著,通常被作為重要的碳匯[9-10]。
BSCs碳固定和釋放受許多因素影響。根據(jù)發(fā)育水平和地殼優(yōu)勢(shì)類(lèi)群,可以將BSCs分為藻、地衣和苔蘚土壤結(jié)皮[11]。不同類(lèi)型的BSCs碳通量具有差異。研究表明,藍(lán)藻的BSCs碳通量為0.06~0.25 μmol·m-2·s-1[12],顯著小于苔蘚的碳通量(2.52 μmol·m-2·s-1)[13],這種差異可能跟BSCs自身組成有關(guān)。此外,BSCs還能夠影響土壤和植被CO2排放。土壤CO2釋放是指土壤碳酸鹽風(fēng)化、植物根系自養(yǎng)呼吸、土壤微生物的異養(yǎng)呼吸、菌根的分解代謝和土壤動(dòng)物的呼吸作用的總和[14]。BSCs能夠通過(guò)增加環(huán)境異質(zhì)性如增加土壤養(yǎng)分含量和微生物數(shù)量對(duì)土壤CO2釋放產(chǎn)生影響[15-16]。BSCs發(fā)育階段與植被群落特征顯著相關(guān)。研究表明,苔蘚能夠顯著降低雜類(lèi)草比例[17]。所以在草地中,不同發(fā)育類(lèi)型的BSCs能夠通過(guò)影響植物群落結(jié)構(gòu)對(duì)草地生態(tài)系統(tǒng)CO2通量產(chǎn)生影響。目前BSCs對(duì)草地生態(tài)系統(tǒng)CO2通量的影響研究較少,不同類(lèi)型的BSCs CO2通量數(shù)量特征和BSCs演替對(duì)土壤、植被以及草地CO2通量的影響還不清楚。
光合作用通常被用作衡量植被固碳能力的一個(gè)重要指標(biāo),在植被碳循環(huán)的研究中得到廣泛關(guān)注。目前,許多學(xué)者通過(guò)測(cè)量植物組織的葉綠素含量來(lái)衡量植被光合潛力,但這種傳統(tǒng)的方法會(huì)破壞植物組織,而葉綠素?zé)晒饧夹g(shù)具有快速測(cè)定和不對(duì)植物造成損傷等優(yōu)點(diǎn),逐漸被應(yīng)用于植物光合生理特性的研究中[18-20]。BSCs葉綠素?zé)晒庾兓哂胁淮_定性,該過(guò)程很大程度上受到近地面小氣候和土壤理化性質(zhì)、土壤基質(zhì)以及微生物群落組成的影響[21]。研究表明環(huán)境因子如污染物、干旱及光輻射是影響B(tài)SCs光合能力的關(guān)鍵因子,缺水與強(qiáng)光輻射均能夠?qū)е翨SCs葉綠素?zé)晒鈪?shù)顯著下降,降低BSCs光合速率[22-23]。此外,BSCs自身發(fā)育階段的不同也會(huì)引起葉綠素?zé)晒鈪?shù)的差異,通常認(rèn)為發(fā)育后期的BSCs具有更高的光合潛力[24]。黃河源退化黑土灘上建植的人工草地內(nèi)分布著大面積的BSCs,但因局部環(huán)境差異如微地形和水熱條件等導(dǎo)致BSCs類(lèi)型和葉綠素?zé)晒膺^(guò)程發(fā)生變化,不同類(lèi)型的生物結(jié)皮葉綠素?zé)晒鈪?shù)變化特征以及關(guān)鍵影響因子還不清楚。
目前草地生態(tài)系統(tǒng)碳循環(huán)的研究多聚焦于高等維管束植物,植被群落之間的BSCs常被忽略,尤其在高寒地區(qū)黑土灘退化草地生態(tài)恢復(fù)中BSCs方面的研究鮮見(jiàn)報(bào)道。由于恢復(fù)治理黑土灘建植人工草地局部環(huán)境的差異性,導(dǎo)致生物結(jié)皮處于不同演替階段,進(jìn)一步增加了人工草地BSCs碳循環(huán)的復(fù)雜性。本研究通過(guò)測(cè)定黃河源區(qū)建植19年人工草地不同類(lèi)型BSCs的光保護(hù)熒光參數(shù)Y(NPQ)(Non photochemical quenching,NPQ)、光損傷熒光參數(shù)Y(NO)(Fluorescence quenching)、葉綠素初始熒光參數(shù)F0(Minimal fluorescence)、最大熒光參數(shù)Fm(Maximal fluorescence)、PS Ⅱ最大光化學(xué)量子產(chǎn)量Fv/Fm(Optimal/Maximal quantum yield of PS Ⅱ)5個(gè)葉綠素?zé)晒鈪?shù)和植被群落以及土壤CO2通量,分析環(huán)境因子對(duì)葉綠素?zé)晒鈪?shù)和CO2通量的影響,以期了解人工草地中不同類(lèi)型生物結(jié)皮的碳釋放和碳固定變化規(guī)律,為后期評(píng)價(jià)BSCs對(duì)草地碳循環(huán)的貢獻(xiàn)以及完善草地生態(tài)系統(tǒng)碳循環(huán)機(jī)制提供理論基礎(chǔ)。
1 材料與方法
1.1 研究區(qū)域概況
研究區(qū)位于青海省東南部、果洛藏族自治州東北部阿尼瑪卿山區(qū)瑪沁縣(98°45′~100°56′ E,33°43′~35°16′ N,海拔4 100~4 500 m)大武灘(圖1)。該區(qū)屬典型的高原山地地貌,氣候類(lèi)型屬青藏高原寒冷氣候,冬季長(zhǎng)而干寒,夏季短而溫濕;年平均氣溫為—3.9~3.8℃,最低氣溫達(dá)—31.4℃;年降水量 492.7 mm,是青海省降雨最多的地區(qū)之一,無(wú)絕對(duì)無(wú)霜期。

圖1 試驗(yàn)樣地地理位置
研究所用人工草地樣地于2000年建成,面積約為1.75 hm2,海拔為3 735 m,由“黑土灘”退化草地機(jī)械翻耕并重新播種建成。農(nóng)藝措施為:深翻—耙平—施肥—撒種—覆土—鎮(zhèn)壓—圍欄封育。所播牧草混播品種有:垂穗披堿草(Elymusnutans)、青海冷地早熟禾(Poacrymophila)和青海中華羊茅(Festucasinensis),混播比例為2∶1∶1。牧草種子由當(dāng)?shù)夭葑逊敝硤?chǎng)生產(chǎn),播量為45 kg·hm-2,施肥量為45 kg·hm-2(磷酸二銨復(fù)合肥),使用C-型肉毒素控制高原鼠兔種群數(shù)量[25-26]。人工草地建植之前的草地類(lèi)型屬于典型“黑土灘”退化草地,優(yōu)勢(shì)種莎草科植物被毒雜草取代,原生植被蓋度不足10%,禿斑裸露地大面積存在。該地主要優(yōu)勢(shì)植物種為甘肅馬先蒿(Ligulariavirgaurea)和鐵棒槌(Aconitumpendulum),次要優(yōu)勢(shì)種為鵝絨委陵菜(Potentillaanserina)和細(xì)葉亞菊(Ajaniatenuifolia)。人工草地生長(zhǎng)季內(nèi)圍欄封育不利用,每年12月至次年4月冬季放牧利用,放牧強(qiáng)度是4只羊·hm-2[27]。
1.2 研究方法
1.2.1植被調(diào)查方法 2019年8月在青海省瑪沁縣大武灘選擇建植19年人工草地,依據(jù)外觀形態(tài)和顏色等特征判斷生物結(jié)皮類(lèi)型,具體詳見(jiàn)秦福雯等方法[28]。其中苔蘚結(jié)皮分布最廣泛,在人工草地中占比為40%左右,其次是地衣(占比15%左右),藻結(jié)皮和物理結(jié)皮分布較少(小于10%)。本文選擇物理結(jié)皮(Physical Crust)和不同類(lèi)型生物結(jié)皮如藻結(jié)皮(Algal Crusts)、地衣結(jié)皮(Lichen Crust)、苔蘚結(jié)皮(Moss Crust)等為研究對(duì)象,調(diào)查人工草地中分布著不同生物結(jié)皮的群落分布特征。利用樣方法調(diào)查各生物結(jié)皮演替階段的群落結(jié)構(gòu),調(diào)查樣方面積為1 m2。隨機(jī)選擇調(diào)查樣方位置,目測(cè)估算樣方內(nèi)植被總蓋度、各經(jīng)濟(jì)類(lèi)群蓋度,清點(diǎn)樣方內(nèi)各經(jīng)濟(jì)類(lèi)群密度,鋼卷尺測(cè)定植被高度;將地上生物量刈割并烘干測(cè)定干物質(zhì)量;利用土鉆(直徑為3 cm)鉆取0~10 cm深度土壤,洗去土壤獲得0~10 cm土層中的根,烘干獲得地下生物量干重。地上地下生物量烘干條件為烘箱65℃條件下將樣品烘干48 h至恒重,使用電子天平稱(chēng)量獲得地上、地下生物量干重。不同演替階段的生物結(jié)皮(除物理結(jié)皮)的植被特征重復(fù)調(diào)查6次。

1.2.3BSCs呼吸和葉綠素?zé)晒鉁y(cè)定 利用土壤碳通量測(cè)量系統(tǒng)LI-8100A紅外氣體分析儀(美國(guó),IR-GA)原位測(cè)定不同生物結(jié)皮演替階段的植被+BSCs+土壤、BSCs+土壤、土壤和物理結(jié)皮CO2通量,CO2通量測(cè)定時(shí)間為上午9:00—11:00[29]。為減小誤差,測(cè)量時(shí)需將氣室垂直埋入土壤0.5~1.0 cm,測(cè)定90 s,記錄CO2通量變化。各測(cè)定方法如下:植被+BSCs+土壤CO2通量,直接將氣室垂直地面放置,測(cè)定CO2通量;BSCs+土壤CO2通量,在確保BSCs完整前提下,將氣室內(nèi)所有維管束植物全部去除,再測(cè)定;土壤CO2通量,繼續(xù)將氣室內(nèi)所有BSCs去除,測(cè)定去除維管束植物和BSCs后地表的CO2通量;物理結(jié)皮CO2通量,直接將氣室垂直地面放置,測(cè)定CO2通量。BSCs的CO2通量,利用BSCs+土壤CO2通量與土壤CO2通量的差值估算。利用基礎(chǔ)型調(diào)制熒光儀Junior-PAM葉綠素?zé)晒鈨x(德國(guó),WALZ)測(cè)定3個(gè)演替階段BSCs的葉綠素?zé)晒鈪?shù)。測(cè)量之前將待測(cè)BSCs暗反應(yīng)20 min,用光強(qiáng)約1 500 μmol·m-2·s-1的飽和脈沖激發(fā)葉綠素分子[30],測(cè)定180 s,記錄光保護(hù)Y(NPQ)、光損傷Y(NO)參數(shù)、最小熒光Fo、最大熒光Fm和Fv/Fm,注意測(cè)定時(shí)調(diào)節(jié)光纖末端距離測(cè)定樣本1 mm左右。
1.3 數(shù)據(jù)處理與分析
本文數(shù)據(jù)利用Excel 2016對(duì)數(shù)據(jù)進(jìn)行整理;利用SPSS 19.0對(duì)不同演替階段BSCs CO2通量、葉綠素?zé)晒鈪?shù)、植被和土壤理化性質(zhì)進(jìn)行單因素方差分析、多重比較;利用Sigma Plot 14.0繪制不同演替階段BSCs CO2通量和葉綠素?zé)晒鈪?shù)分布圖;利用R4.0對(duì)不同演替階段BSCs呼吸、葉綠素?zé)晒鈪?shù)與植被特征、土壤理化性質(zhì)進(jìn)行冗余分析,在RDA分析之前,對(duì)環(huán)境因子進(jìn)行Hellinger轉(zhuǎn)換,并對(duì)R2進(jìn)行校正,最后繪制圖。
2 結(jié)果與分析
2.1 不同類(lèi)型BSCs的CO2通量和葉綠素?zé)晒鈪?shù)
對(duì)不同類(lèi)型BSCs和植被CO2通量分析發(fā)現(xiàn)(圖2):藻結(jié)皮和地衣結(jié)皮對(duì)應(yīng)所處的植被和土壤群落(植被+BSCs+土壤)CO2通量分別為19.91 μmol·m-2·s-1和20.13 μmol·m-2·s-1,顯著高于苔蘚結(jié)皮CO2通量(15.87 μmol·m-2·s-1,P<0.05)。不同類(lèi)型生物結(jié)皮的BSCs+土壤CO2通量變化趨勢(shì)與植被+BSCs+土壤CO2通量變化相似,均為藻和地衣結(jié)皮類(lèi)型顯著高于苔蘚(P<0.05)。但對(duì)比不同類(lèi)型的BSCs CO2通量發(fā)現(xiàn),苔蘚有較高的CO2通量(10.58 μmol·m-2·s-1),顯著高于地衣和藻結(jié)皮(P<0.05)。BSCs能夠引起土壤呼吸速率發(fā)生變化,不同類(lèi)型生物結(jié)皮下的土壤CO2通量不同。對(duì)比不同類(lèi)型結(jié)皮下土壤CO2通量發(fā)現(xiàn),所有類(lèi)型生物結(jié)皮均能夠提高土壤呼吸強(qiáng)度,其中藻和地衣結(jié)皮效果更顯著(P<0.05),它們的CO2通量約是物理結(jié)皮土壤CO2通量的2倍,而苔蘚下土壤CO2通量與物理結(jié)皮相近。
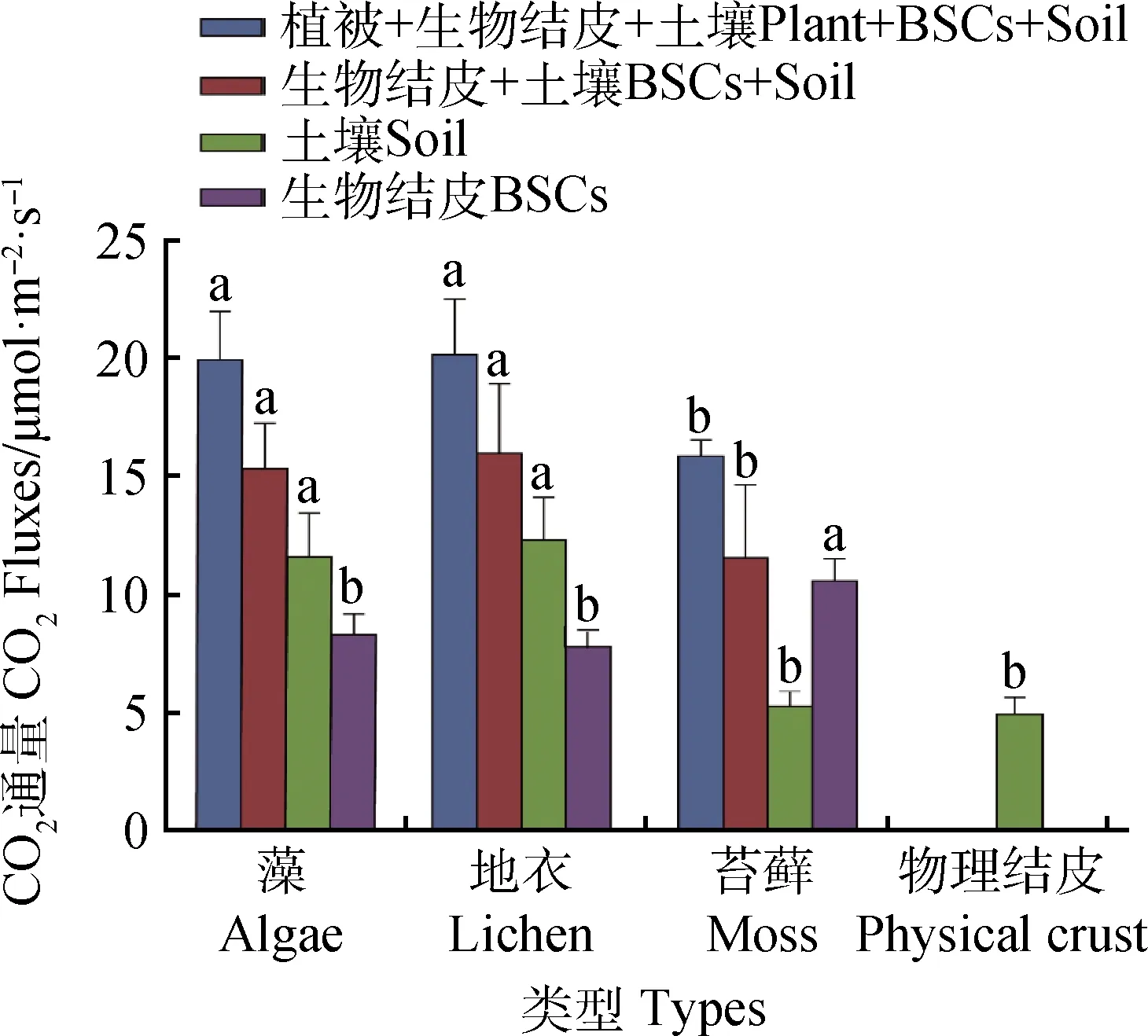
圖2 不同類(lèi)型BSCs CO2通量
圖3為不同類(lèi)型BSCs葉綠素?zé)晒鈪?shù)變化特征。由圖3-I和3-II可知:不同類(lèi)型BSCs的Y(NO)具有顯著差異(P<0.05),其中地衣最高(0.567),苔蘚最低(0.246)。不同類(lèi)型BSCs的Y(NPQ)變化趨勢(shì)不同,苔蘚值最大,為0.605,約是地衣的2.4倍。圖3-III和3-IV分別表示不同類(lèi)型BSCs初始熒光參數(shù)(F0)和最大熒光參數(shù)(Fm)變化特征,從圖中可以看出藻-地衣-苔蘚過(guò)程中,F(xiàn)0和Fm逐漸增加,苔蘚生物結(jié)皮的值最大,分別為135.16(F0)和286.5(Fm),顯著高于藻和地衣結(jié)皮(P<0.05)。圖3-V是不同類(lèi)型BSCs光合潛力(Fv/Fm)大小變化特征,不同類(lèi)型BSCs的Fv/Fm值排列順序?yàn)樘μ\>藻>地衣,彼此之間差異顯著(P<0.05)。

圖3 不同類(lèi)型BSCs葉綠素?zé)晒鈪?shù)
2.2 CO2通量、葉綠素?zé)晒鈪?shù)與環(huán)境因子關(guān)系
表1為不同BSCs階段植被變化特征,從表中能夠看出不同類(lèi)型BSCs對(duì)應(yīng)所處的植被特征有較大差異。就植被總蓋度而言,苔蘚階段植被蓋度最高,為72%,地衣階段植被覆蓋度最低,僅為29.17%,各BSCs階段植被覆蓋度差異顯著(P<0.05)。分析不同類(lèi)型BSCs的不同經(jīng)濟(jì)類(lèi)群(禾本科、莎草科、雜類(lèi)草)蓋度和高度發(fā)現(xiàn),就禾本科植物而言,苔蘚階段禾本科植物蓋度和高度較其他經(jīng)濟(jì)類(lèi)群大,分別為23%和19.05 cm;就莎草科而言,苔蘚階段莎草科蓋度顯著高于藻和地衣階段(P<0.05),但高度沒(méi)有表現(xiàn)出顯著的優(yōu)勢(shì)。雜類(lèi)草在苔蘚階段蓋度最大(48.00%),顯著高于地衣階段。對(duì)比同一類(lèi)型生物結(jié)皮不同經(jīng)濟(jì)類(lèi)群蓋度和高度發(fā)現(xiàn),不同經(jīng)濟(jì)類(lèi)群蓋度高度特征表現(xiàn)出一定差異性。各BSCs演替階段雜草蓋度最高,而莎草科植物蓋度最低。但各BSCs階段禾本科植物高度均占優(yōu)勢(shì),均顯著高于其他兩個(gè)經(jīng)濟(jì)類(lèi)群(P<0.05)。就生物量而言,地衣階段地上和地下生物量最高,分別為293.97 g·m-2和22.8 kg·m-3,其中地上生物量顯著高于藻和苔蘚結(jié)皮(P<0.05)。

表1 不同BSCs類(lèi)型植被分布特征
圖4為不同類(lèi)型BSCs階段不同經(jīng)濟(jì)類(lèi)群植被密度特征圖。分析發(fā)現(xiàn),地衣和苔蘚階段禾本科植物密度顯著高于藻類(lèi)階段(P<0.05)。就莎草科植物而言,藻類(lèi)和地衣階段的莎草科植物密度顯著低于苔蘚時(shí)期(P<0.05);而雜類(lèi)草密度在不同生物結(jié)皮階段差異不顯著。通過(guò)對(duì)比相同階段生物結(jié)皮不同經(jīng)濟(jì)類(lèi)群植物密度發(fā)現(xiàn),各生物結(jié)皮階段的莎草科植物均顯著低于禾本科和雜類(lèi)草植物密度(P<0.05)。

圖4 不同生物結(jié)皮階段各經(jīng)濟(jì)類(lèi)群植被密度
不同類(lèi)型生物結(jié)皮土壤理化性質(zhì)分布特征見(jiàn)表2。由表可得,物理結(jié)皮土壤表層溫度顯著高于生物結(jié)皮覆蓋下的土壤表層溫度(P<0.05),即生物結(jié)皮對(duì)維持地面溫度效果顯著,同時(shí)對(duì)比不同類(lèi)型BSCs發(fā)現(xiàn),苔蘚效果最好。就土壤體積含水率而言,不同類(lèi)型的生物結(jié)皮均顯著高于物理結(jié)皮表層土壤體積含水率(P<0.05)。苔蘚類(lèi)生物結(jié)皮保水能力最強(qiáng),其覆蓋下表層土壤體積含水率最高,為33.53%;地衣次之,藻類(lèi)結(jié)皮效果較差,僅為25.35%。就電導(dǎo)率而言,物理結(jié)皮與地衣結(jié)皮之間差異顯著(P<0.05)。苔蘚結(jié)皮對(duì)改良土壤pH的效果較其他類(lèi)結(jié)皮更好(P<0.05)。各類(lèi)型生物結(jié)皮均能積累土壤養(yǎng)分,但不同類(lèi)型生物結(jié)皮存在一定差異。其中苔蘚和地衣對(duì)土壤全氮、全磷、銨態(tài)氮和速效磷的積累效果較好,而藻類(lèi)對(duì)硝態(tài)氮、速效磷積累效果顯著。

表2 不同BSCs類(lèi)型土壤理化性質(zhì)
圖5為植被、土壤理化性質(zhì)和土壤微生物與CO2通量的RDA排序圖,從圖可以看出RDA模型承載的總方差比例(R2)均達(dá)到了70%以上,其中植被—CO2通量RDA模型R2達(dá)到了82.91%。說(shuō)明以上所有模型均能夠較好解釋環(huán)境因子與BSCs CO2通量的關(guān)系。植被分布的差異性會(huì)引起B(yǎng)SCs的CO2通量發(fā)生變化。由RDA排序可知植被中禾本科和雜類(lèi)草的植被高度為主控因子,CO2通量與禾本科、雜草的植被高度呈現(xiàn)出較強(qiáng)的正相關(guān)關(guān)系(圖5Ⅰ),表明禾本科和雜草植物的長(zhǎng)高有助于提高不同BSCs演替階段各類(lèi)群的CO2排放速率。土壤理化性質(zhì)中銨態(tài)氮含量與土壤pH為主控因子,其中銨態(tài)氮含量與CO2通量呈正相關(guān)關(guān)系,而土壤pH與CO2通量表現(xiàn)出較強(qiáng)的負(fù)相關(guān)關(guān)系,表明較多的速效養(yǎng)分和較低的土壤pH有利于促進(jìn)各類(lèi)群的生長(zhǎng)呼吸(圖5Ⅱ)。
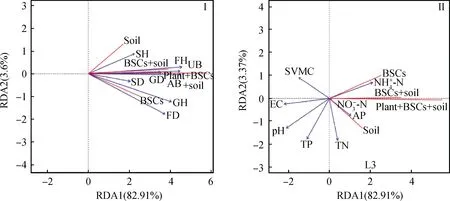
圖5 CO2通量和環(huán)境因子的RDA排序
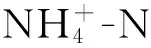
圖6 葉綠素?zé)晒鈪?shù)和環(huán)境因子的RDA排序
3 討論
3.1 不同類(lèi)型生物結(jié)皮CO2通量的變化
不同發(fā)育和演替狀態(tài)以及不同環(huán)境下BSCs的優(yōu)勢(shì)物種、生理、生化等特征各不相同[8],因此不同類(lèi)型BSCs的CO2通量不同。一些研究表明,BSCs演替后期的苔蘚結(jié)皮具有更高的呼吸速率[29,31],本文得出相似結(jié)論(圖2)。研究表明CO2釋放量和釋放速率與有機(jī)體的生物量顯著正相關(guān)[32],演替后期的苔蘚具有較高的生物量,所以苔蘚自身的CO2通量也較大。同時(shí),BSCs也能增加土壤呼吸,但不同類(lèi)型BSCs對(duì)土壤CO2通量的影響具有差異性,這種差異性可能是由于土壤中微生物群落的呼吸活動(dòng)的不同所致[32]。研究發(fā)現(xiàn),不同BSCs對(duì)微生物的積累具有差異性,藻和地衣類(lèi)下微生物數(shù)量更多[33]。土壤微生物是釋放土壤CO2的主要貢獻(xiàn)者,微生物數(shù)量較多導(dǎo)致呼吸加強(qiáng),土壤CO2釋放速率也就較快。因此,藻和地衣結(jié)皮覆蓋下的土壤會(huì)釋放更多的CO2。除微生物數(shù)量外,外界環(huán)境因子也是影響土壤CO2通量的重要因子。與干旱半干旱地區(qū)相比,高寒地區(qū)人工草地相同演替階段的生物結(jié)皮均具有更高的CO2通量[29,34]。與沙漠地區(qū)相比,黃河源區(qū)人工草地土壤具有較高濕度和養(yǎng)分含量,有利于促進(jìn)BSCs物質(zhì)量的積累,提高BSCs生理生化反應(yīng)速率,而沙漠地區(qū)因受到水分及土壤養(yǎng)分條件的限制,該地區(qū)的土壤和BSCs的CO2通量較低。有研究表明水分是影響B(tài)SCs生長(zhǎng)發(fā)育的首要影響因子[35-36],本文通過(guò)冗余分析得出不同結(jié)論。本文研究結(jié)果表明,在土壤理化性質(zhì)中養(yǎng)分和pH為主要影響因子,可能是因?yàn)楸驹囼?yàn)樣區(qū)地處黃河源地區(qū),降水較充足,所以該地區(qū)水分不是首要限制因子,但土壤養(yǎng)分和pH是影響草地BSCs生長(zhǎng)演替的重要驅(qū)動(dòng)因子[37],所以二者可能通過(guò)影響B(tài)SCs演替對(duì)土壤以及BSCs本身CO2釋放速率產(chǎn)生影響。
BSCs作為高寒地區(qū)人工草地演替過(guò)程中出現(xiàn)的重要伴生種,其碳循環(huán)功能的發(fā)揮與草地植被群落息息相關(guān)。冗余分析表明,禾本科植物是影響B(tài)SCs CO2通量的主控因子。作為高寒地區(qū)人工草地主要優(yōu)勢(shì)植物物種,禾本科植物的分布對(duì)BSCs和土壤CO2釋放具有重要意義,禾本科植物能夠通過(guò)積累土壤養(yǎng)分以及為BSCs生長(zhǎng)提供合適的生存環(huán)境來(lái)促進(jìn)BSCs生長(zhǎng)發(fā)育[38],例如禾本科植物生長(zhǎng)過(guò)程中能夠?yàn)锽SCs提供散射光以及降低土壤水分蒸發(fā)速率,從而緩解BSCs受到的光和干旱脅迫,使BSCs生理和生化活動(dòng)均處于活躍狀態(tài),促進(jìn)CO2釋放。但禾本科植被的生長(zhǎng)繁殖也會(huì)與BSCs形成競(jìng)爭(zhēng)關(guān)系,如較多禾本科凋落物嚴(yán)重降低了BSCs蓋度和厚度,特別是對(duì)苔蘚影響最大。苔蘚類(lèi)植物體積累生物量的主要途徑是光合作用,而較多凋落物使苔蘚植物長(zhǎng)時(shí)間處于遮光狀態(tài),不能進(jìn)行光合作用,最終導(dǎo)致苔蘚植物體死亡。生態(tài)系統(tǒng)中沒(méi)有獨(dú)立關(guān)系存在,植被影響B(tài)SCs的同時(shí),BSCs也會(huì)影響植被生長(zhǎng)。研究表明,苔蘚階段有較高的植被總蓋度,但該生物結(jié)皮階段植被總CO2通量最低(圖2),所以苔蘚可能能夠影響植被群落結(jié)構(gòu)特征,進(jìn)而減少草地生態(tài)系統(tǒng)CO2的排放。
3.2 不同類(lèi)型生物結(jié)皮葉綠素?zé)晒鈪?shù)的變化
BSCs光合活性或光和潛力受自身組成的影響[39]。不同類(lèi)型的BSCs群落光合優(yōu)勢(shì)種具有差異性,導(dǎo)致生物結(jié)皮光合能力的不同。有研究指出,藻結(jié)皮的Fv/Fm約為0.4,而苔蘚結(jié)皮的Fv/Fm超過(guò)0.5,顯著高于藻結(jié)皮[24],本研究也得出相似結(jié)論(圖3-V)。這可能是因?yàn)樵褰Y(jié)皮階段主要的光合生物是藍(lán)藻(原核生物)和藻類(lèi)植物[40],而苔蘚階段光合生物主要是苔蘚。原核生物藍(lán)藻中,由于缺乏葉綠體結(jié)構(gòu),光合作用和呼吸中的電子轉(zhuǎn)移都發(fā)生在類(lèi)囊體膜內(nèi),兩個(gè)電子轉(zhuǎn)移鏈同時(shí)共享大量的電子轉(zhuǎn)移中間體,在一定程度上降低了光合電子傳遞效率[41],導(dǎo)致光合效率降低。而演替后期的苔蘚具有獨(dú)立的呼吸和光合場(chǎng)所,光合電子傳遞效率較其他類(lèi)型生物結(jié)皮高,所以苔蘚階段光合效率高。
藻類(lèi)、地衣和苔蘚都屬于光合生物,這些光合生物主要通過(guò)光合色素(如葉綠素和藻藍(lán)蛋白)吸收和轉(zhuǎn)化光能,所以光合色素含量也是影響植被光合作用的關(guān)鍵因子。不同類(lèi)型的BSCs群落的色素分子不同,故其光能轉(zhuǎn)化效率具有一定差異[42]。研究發(fā)現(xiàn),與演替后期的苔蘚類(lèi)生物結(jié)皮相比,藻類(lèi)結(jié)皮中的光合生物量較低,各種光合色素如葉綠素A和B含量也較低,故表現(xiàn)出較低的F0,F(xiàn)m和Fv/Fm,該結(jié)果與Pushkareva等研究結(jié)果相似[43]。而在苔蘚階段,苔蘚占據(jù)光合生物量的90%以上,該階段的光合生物量和光合體中的光合色素含量顯著高于藻類(lèi)[44],光合能力增強(qiáng),葉綠素?zé)晒鈪?shù)如F0,F(xiàn)m和Fv/Fm增加,顯著高于藻結(jié)皮(約2倍)。同時(shí),BSCs抗逆性(抗光損害)的強(qiáng)弱也對(duì)BSCs光合作用有顯著影響。不同類(lèi)型的BSCs的Y(NO),Y(NPQ)不同。Y(NO)、Y(NPQ)分別是光損傷和光保護(hù)重要指標(biāo)。本文研究表明地衣類(lèi)結(jié)皮具有高的Y(NO)和低的Y(NPQ)值,說(shuō)明地衣對(duì)光調(diào)節(jié)和抗光損害和忍受強(qiáng)光的能力最差,可能是因?yàn)榈匾骂?lèi)結(jié)皮色素含量最低,而植物的光保護(hù)色素如類(lèi)胡蘿卜素含量缺失導(dǎo)致地衣結(jié)皮在受到較強(qiáng)的光脅迫時(shí),有機(jī)體內(nèi)沒(méi)有足夠的色素來(lái)消除組織內(nèi)過(guò)多的活性氧化物(ROS)[45],地衣結(jié)皮受到光損傷,最終引起有機(jī)體光合能力下降。苔蘚的Y(NPQ)指數(shù)顯著大于地衣和藻結(jié)皮(圖3-I),可能是因?yàn)樘μ\的絲核菌素合成能力的提升,其基內(nèi)菌絲產(chǎn)生各種水溶性或脂溶性的色素從而增強(qiáng)BSCs抵抗強(qiáng)光脅迫能力[46]。青藏高原地區(qū)海拔高,光輻射強(qiáng)度大,BSCs容易受到光脅迫,生理生化活動(dòng)降低,所以在該地區(qū)散射光的形成和利用對(duì)BSCs生長(zhǎng)演替以及光合等生理作用十分重要。冗余分析發(fā)現(xiàn),在黃河源區(qū)人工草地中優(yōu)勢(shì)種禾本科植物(表1)的高度是影響B(tài)SCs葉綠素?zé)晒鈪?shù)的主控因子。較高的植被高度能夠減小BSCs被太陽(yáng)輻射直射的機(jī)會(huì),有利于BSCs在避免強(qiáng)光脅迫的同時(shí)高效的進(jìn)行光合作用。同時(shí),青藏高原上BSCs對(duì)非降水形式的水資源的利用也十分重要,較高的植被有利于凝結(jié)水的形成,在非降水時(shí)期為BSCs提供充足水資源進(jìn)行光合作用。研究發(fā)現(xiàn),苔蘚本身也能夠顯著提升土壤濕度(表2),促進(jìn)自身和禾本科植物生長(zhǎng)(表1),能夠進(jìn)一步優(yōu)化BSCs生長(zhǎng)和光合的條件。因此,高寒地區(qū)人工草地中禾本科植物與BSCs互利生長(zhǎng)。土壤理化性質(zhì)中,銨態(tài)氮以及pH為主控因子(圖6Ⅱ)。養(yǎng)分和土壤pH顯著影響B(tài)SCs生長(zhǎng)和演替[47-48],如養(yǎng)分充足有利于苔蘚的建植,藻類(lèi)BSCs更趨向于土壤pH低的地方生長(zhǎng)[49],故pH低的地方多分布藻結(jié)皮,光合效率較低。
4 結(jié)論
高寒地區(qū)人工草地中苔蘚結(jié)皮CO2通量最大,但苔蘚階段植被群落和土壤CO2通量最低,故苔蘚有利于減少草地系統(tǒng)碳的外排。苔蘚能促進(jìn)禾本科和土壤養(yǎng)分恢復(fù),地衣有效積累了人工草地地下生物量,而藻類(lèi)與莎草科植物的恢復(fù)息息相關(guān),所以生物結(jié)皮有利于促進(jìn)黑土灘的生態(tài)恢復(fù);苔蘚較其他類(lèi)型BSCs具有更大的光合和抗逆潛力;禾本科高度和蓋度、銨態(tài)氮含量和pH均是BSCs呼吸和光合作用的重要影響因子。綜上,生物結(jié)皮尤其是苔蘚結(jié)皮提高了人工草地碳源能力,同時(shí)對(duì)促進(jìn)黑土灘退化草地向高寒草甸的演替有積極重要的作用。
