環割和刻芽對‘紅光2號’蘋果1年生和2年生枝條萌芽特性的影響*
2021-09-12 03:20:42王愛斌孟紅志杜宜南王立飛解金斗周宏宇
中國果樹 2021年9期
王愛斌,孟紅志,閆 帥,杜宜南,王立飛,解金斗,周宏宇
(1 河北農業大學科教興農中心,保定071001)(2 中國消防救援學院)(3 遷安市自然資源和規劃局)(4 中國農業科學院果樹研究所)(5 河北威縣省級農業科技園區)
近年來,國內在積極推廣蘋果矮砧密植栽培模式,其中,幼樹整形尤為關鍵,措施不當會造成靠近中心干基部的枝條萌芽率低,中心干附近枝量少,嚴重影響幼樹枝量和產量增加[1-3]。蘋果具有較強的頂端優勢,‘國光’品種尤為明顯,枝條萌發率極低,常出現“掃帚頭”現象。生產中常利用拉枝、刻芽、環割、環剝、摘心、短截和藥劑處理等消除蘋果樹頂端優勢,促進枝條萌發[4]。不同處理方式對蘋果樹萌芽、成枝影響不同[5-6],環割和刻芽操作簡便,是最常用和最環保的方式。環割和刻芽后,抽生的枝條可牽制背上枝,抑制背上枝生長,從而能夠有效調整枝類比,維持中庸樹勢[7-9]。研究表明,環割和刻芽可改變蘋果枝條芽體中內源激素和營養物質的平衡[5,8-13],如刻芽可改變蘋果幼樹萌芽期芽體中GA3[10]和ABA的含量[11],而環割可顯著影響蘋果幼樹芽體可溶性糖和淀粉含量[12]。另外,梨幼樹芽體中可溶性糖、淀粉和蛋白質含量均會受到促萌芽措施的影響[13]。果樹環割和刻芽引起的內源激素和營養物質變化,對于研究芽體萌發能力和成枝能力非常重要。雖然前人對于果樹環割和刻芽促發枝的研究有了一些報道,但針對‘國光’的研究鮮有報道。
‘國光’是我國北方蘋果產區的主栽品種之一,近些年來因其著色和成枝力差等問題給產業發展造成了嚴重的影響[14]。張家口和承德地區是我國‘國光’主產區,‘紅光2號’是從兩地‘國光’中選出的紅色芽變品種[15],解決了‘國光’著色差的問題,但其成枝力差的難題還未完全解決[16]。因此,采取合理的措施增加樹體中心干基部枝量,是提高單株產量的重要保障。前人的研究[7,10-12]多集中在對蘋果樹1年生枝條芽體成枝情況,針對1年生和2年生枝條同時開展芽體成枝情況的研究很少。本研究以‘紅光2號’1年生和2 年生枝條為材料,探究環割和刻芽對其芽體萌芽率、成枝比率、內源激素、可溶性糖、淀粉和蛋白質含量的影響,以期找出‘紅光2號’促枝的最佳處理方式和芽體萌發生理生化特性的差異,為今后‘紅光2號’的栽培提供理論基礎。
1 材料與方法
1.1 試驗材料
試驗在河北省懷來縣永安村進行。試驗園土壤為壤土,0~50 cm 土層有機質含量為40.39 g/kg,速效氮含量為115.85 mg/kg,速效磷含量為32.41 mg/kg,速效鉀含量為152.51 mg/kg,pH 值為7.7。
試驗園東西行向,行株距4 m×2 m,樹形為細長紡錘形。供試‘紅光2 號’樹基砧為八棱海棠,中間砧為‘SH38’,4 年生,高3.5 m,長勢一致。
1.2 環割和刻芽處理
2019年4 月14日,選取位置相同、長勢一致的‘紅光2號’1年生和2 年生枝條,分別環割或刻芽;對照按上述標準選枝,不處理,做好標記。每株同時進行環割和刻芽處理,并設對照,每個處理2 個枝,單株小區,9 次重復。
環割,在芽體上方5 mm 處環割1圈,深達木質部。刻芽,用小刀在芽上方5 mm 處橫切,深達木質部,長度為枝條周長的1/3。由于基部芽體發育較差,為保證試驗的可靠性,從枝條基部向上數第5個芽開始進行,每個芽環割1 刀或刻1刀,連續進行到第20 個芽為止。
1.3 萌芽成枝情況調查
2019 年10 月20 日調查和統計各處理的萌芽成枝情況,每個處理調查6 株樹,共12個枝條。
1.4 內源激素、可溶性糖、淀粉和蛋白質含量的測定
2019 年5月17日,每個處理隨機選取3株樹,每株樹在處理枝上取20 個芽(未萌發),1株為1次重復,3次重復,放入液氮罐帶回實驗室保存待測。可溶性糖及淀粉含量采用硫酸蒽酮比色法測定[17],蛋白質含量采用考馬斯亮藍法測定[18]。采用酶聯免疫法[11](ELISA)測定上述樣品中的吲哚乙酸(IAA)、赤霉素(GA3)、脫落酸(ABA)和玉米素核苷(ZR)含量,ELISA 試劑盒由中國農業大學提供。
1.5 數據處理與分析
采用Microsoft Excel進行數據統計和制表,采用SPSS 19.0進行Duncan’s差異顯著性檢驗。
2 結果與分析
2.1 環割和刻芽對‘紅光2號’枝條萌芽和成枝的影響
由表1可知,‘紅光2 號’1年生枝環割、刻芽的萌芽率分別為96.3%和81.0%,均顯著高于對照(36.5%)。2年生枝環割后萌芽率為91.9%,顯著高于刻芽和對照;刻芽后萌芽率為54.4%,顯著高于對照(11.7%)。各處理1年生枝萌芽率均高于同樹的2 年生枝。
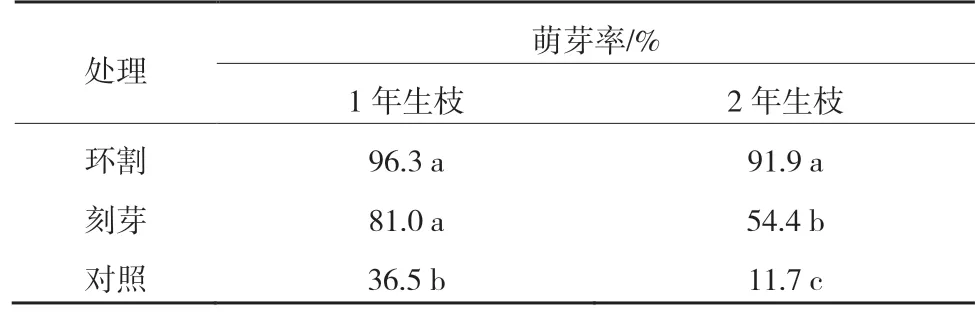
表1 ‘紅光2號’環割和刻芽后的萌芽率
從表2可以看出,‘紅光2 號’1年生和2年生枝環割處理短枝比率均顯著低于刻芽,二者短枝比率均顯著低于對照;環割處理中枝比率均顯著高于刻芽,二者中枝比率均顯著高于對照;環割處理長枝比率均顯著高于刻芽,對照均未抽生出長枝。
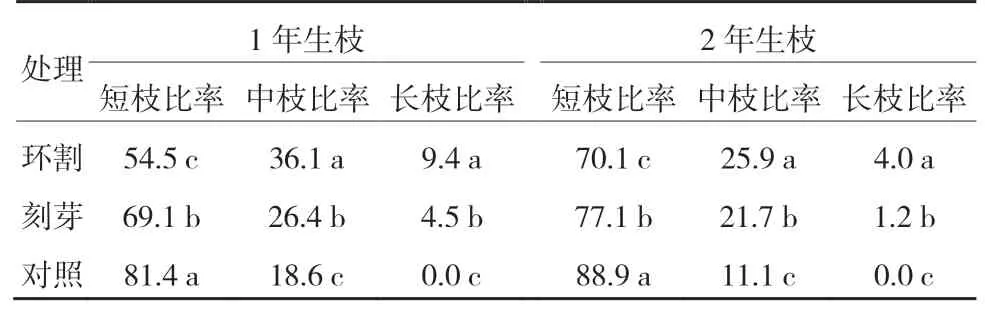
表2 ‘紅光2號’環割和刻芽后的枝類比 %
2.2 環割和刻芽對‘紅光2號’芽體內源激素含量及GA3/(IAA+ABA+ZR)值的影響
2.2.1 內源激素含量
春季環割和刻芽可增加‘紅光2 號’1年生枝和2年生枝芽內GA3含量,降低IAA、ABA和ZR含量。從表3可以看出,‘紅光2 號’1年生枝環割和刻芽處理芽內IAA、ABA和ZR 含量均顯著低于對照,環割處理ABA 和IAA含量均顯著低于刻芽;環割和刻芽處理GA3含量均顯著高于對照,對照GA3含量最低,為4.04 ng/g,環割和刻芽處理GA3含量差異不顯著。
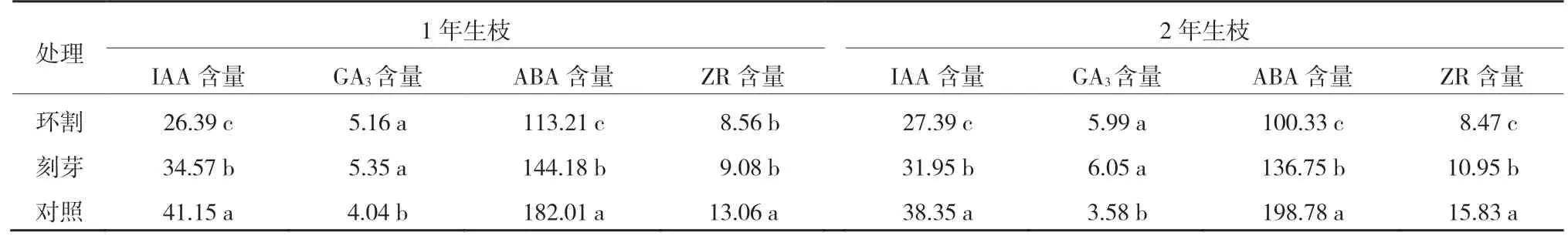
表3 ‘紅光2號’1年生枝和2年生枝環割和刻芽后的芽體內源激素含量 ng/g
‘紅光2 號’2年生枝環割和刻芽處理芽內IAA、ABA和ZR 含量均顯著低于對照,環割處理IAA、ABA 和ZR 含量均顯著低于刻芽;GA3含量變化與1年生枝結果一致,環割和刻芽處理GA3含量均顯著高于對照,對照GA3含量最低,為3.58 ng/g,環割和刻芽處理GA3含量差異不顯著(表3)。
2.2.2 GA3/(IAA+ABA+ZR)值
環割和刻芽均能提高‘紅光2號’1年生枝和2年生枝芽內GA3/(IAA+ABA+ZR)值,且環割效果更為明顯。從表4 可以看出,‘紅光2號’1 年生枝環割處理GA3/(IAA+ABA+ZR)值顯著高于對照,刻芽處理GA3/(IAA+ABA+ZR)值與環割處理和對照差異均不顯著。2年生枝環割和刻芽處理GA3/(IAA+ABA+ZR)值均顯著高于對照,分別是對照的3.10、2.38倍,環割處理GA3/(IAA+ABA+ZR)值顯著高于刻芽。

表4 ‘紅光2號’1年生枝和2年生枝環割和刻芽后芽內GA3/(IAA+ABA+ZR)值
2.3 環割和刻芽對‘紅光2號’芽內可溶性糖、淀粉和蛋白質含量的影響
‘紅光2 號’1年生枝和2年生枝春季環割和刻芽可明顯提高芽內蛋白質含量,降低可溶性糖含量。從表5可以看出,‘紅光2 號’1年生枝環割和刻芽處理芽內可溶性糖含量均顯著低于對照,環割和刻芽處理芽內可溶性糖含量差異不顯著,對照芽內可溶性糖含量最高,為2.59%;各處理芽內淀粉含量差異均不顯著;環割和刻芽處理芽內蛋白質含量均顯著高于對照,環割和刻芽處理芽內蛋白質含量差異不顯著,環割處理芽內蛋白質含量最高,為6.83 mg/g。
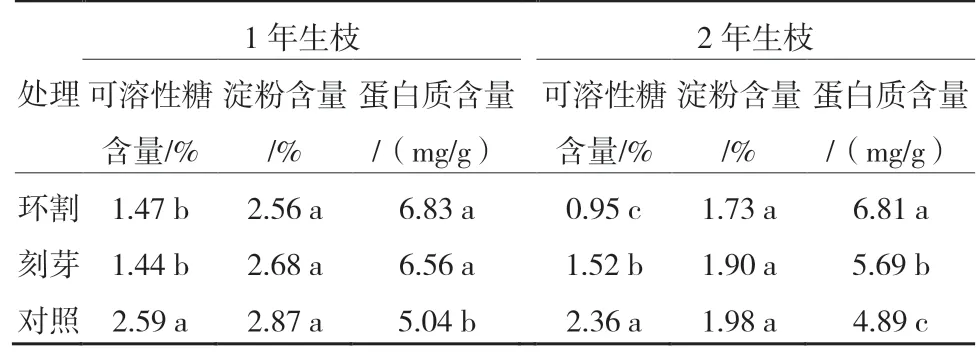
表5 ‘紅光2號’1年生枝和2年生枝環割和刻芽后芽內可溶性糖、淀粉和蛋白質含量
2 年生枝環割和刻芽處理芽內可溶性糖含量均顯著低于對照,環割處理芽內可溶性糖含量顯著低于刻芽處理,環割處理芽內可溶性糖含量最低,為0.95%;各處理芽內淀粉含量差異均不顯著;環割和刻芽處理芽內蛋白質含量均顯著高于對照,環割處理芽內蛋白質含量最高,為6.81 mg/g,顯著高于刻芽處理(表5)。
3 討論與結論
環割和刻芽是果樹生產中最常見的促分枝手段,具有操作簡單和綠色環保等特點[11]。環割和刻芽對提高果樹幼旺枝條的萌芽、增加枝量、花芽形成和緩和樹勢等均有促進作用[10,12]。前人對不同樹種研究發現,環割或刻芽能有效促進‘三季’梨[19]、‘紅富士’蘋果[20]、‘南部魁’蘋果[20]和‘黃冠’梨[21]的芽體萌發成枝。本研究表明,環割和刻芽對‘紅光2號’提高1年生枝和2年生枝萌芽和成枝效果明顯,均能有效提高枝條的萌芽率和中長枝比率;同時,還發現環割促發枝效果明顯優于刻芽,這可能與環割和刻芽對枝條韌皮部造成損傷程度不同有關,不同損傷程度會影響芽內各類營養物質的積累,進而影響芽體的萌發成枝。環割和刻芽均可促使枝條芽體抽生枝,這樣既有利于果樹早實豐產,也有利于結果枝的壽命延長和數量增加,同時對于合理的樹體結構培養也非常重要。但進行環割和刻芽時應注意以下2 個問題:當處理后導致母枝背上部位極性芽出現徒長枝時,需在生長季及時扭梢來控制;對壯枝和芽體較為飽滿的芽可以采用間隔處理,若處理后發枝過密,生長季需有意識地調控過密枝[21]。
植物內源激素在植物體內含量甚微,但其生理生化效應明顯[22],果樹萌芽成枝是一個較為復雜的生理生化和形態變化過程,內源激素IAA、GA3、ABA、ZR 與果樹枝條的萌芽密切相關[11]。IAA具有促進生長、頂端優勢和延長休眠的特性,GA3具有促進營養生長和打破休眠的特性,ABA具有抑制生長和促進脫落的特性,ZR具有促進細胞分裂和調控分化的特性,各激素因其濃度不同也會導致其作用程度和效果存在差異[22-24]。牛自勉等[10]研究認為,GA3含量的提高和ABA含量的降低是蘋果枝條萌芽率變化的內在因素。本研究結果表明,‘紅光2號’1年生枝和2 年生枝在春季環割和刻芽后,對應芽體內GA3含量會升高,ABA含量降低,同時環割和刻芽后的萌芽率也明顯增高,這個結果與牛自勉等的研究結果一致。新梢芽內IAA含量增加到一定程度,生長就會停止[25]。
前人研究表明[26],IAA隨濃度升高,其促進芽體萌發的能力越強,但IAA具有雙重性,達到一定濃度后,越高對芽體促萌效果越差。本研究結果發現,‘紅光2 號’1年生枝和2年生枝在春季環割和刻芽后,對應芽體內IAA含量下降,且環割下降程度最為明顯,這可能是由于環割和刻芽對韌皮部造成損傷程度不同進而影響激素運輸阻力不同,并最終導致IAA 在芽體內的積累產生差異。通過本研究的結果推測,環割和刻芽促進‘紅光2 號’1年生枝和2 年生枝分枝的原因,可能是由于這2項農藝操作改變了芽體內源激素的平衡,即GA3促進激素含量的升高和ABA抑制激素含量的降低以及IAA激素雙重性抑制作用的體現。
前人研究認為,果樹萌芽期芽體中ZR含量會升高,進而促進側芽的萌發和生長[13,27]。相反,本研究結果表明,環割和刻芽降低了芽體中ZR的含量,此結果與艾沙江·買買提等[11]研究刻芽對‘富士’蘋果3 年生幼樹促萌芽的結果一致。這可能是由于根源ZR通過木質部隨蒸騰液流先運輸到莖尖,然后經過韌皮部運輸到側芽處,環割和刻芽損傷了韌皮部,故導致ZR含量降低。同時,也說明ZR 激素可能對促進‘紅光2 號’萌芽的作用較小,或者ZR 激素含量的變化不能改變激素綜合調控的平衡。
‘紅光2號’1年生枝和2 年生枝環割和刻芽后,各處理芽體中促進性激素和抑制性激素之間的比值發生了變化,芽內GA3/(IAA+ABA+ZR)值均明顯提高,前人也有類似結果[11],說明環割和刻芽打破了原有內源激素之間的平衡,進而有利于促進芽體的萌發成枝。各內源激素含量和比值的變化可能是‘紅光2 號’枝條萌芽率發生變化的內在誘因之一。
綜上所述,環割和刻芽均能有效提高‘紅光2號’1年生枝和2年生枝的萌芽率和中長枝比率,環割效果明顯優于刻芽。環割和刻芽處理可使芽內蛋白質含量明顯升高,可溶性糖含量明顯降低,淀粉含量變化不明顯。同時,還發現環割和刻芽后處理芽內IAA、ABA和ZR含量明顯降低,而GA3含量和GA3/(IAA+ABA+ZR)值明顯升高。
