PRKAG3基因在雞不同部位肌肉中的表達及其與肌纖維類型的相關性
2021-09-11 01:27:46章明單艷菊姬改革巨曉軍劉一帆屠云潔鄒劍敏束婧婷
江蘇農業科學 2021年16期
章明 單艷菊 姬改革 巨曉軍 劉一帆 屠云潔 鄒劍敏 束婧婷


摘要:PRKAG3是影響畜禽肉品質的關鍵基因。為研究PRKAG3基因在雞不同部位肌肉中的表達及其與肌纖維類型的相關性,以隱性白羽肉雞為研究對象,用ATPase堿孵育法對不同部位肌肉肌纖維類型進行分析,采用實時熒光定量PCR法檢測PRKAG3基因在這些肌肉組織中的表達并將其與肌纖維類型進行關聯分析。結果表明,PRKAG3基因在Ⅰ型肌纖維比例相對較高的比目魚肌和縫匠肌中表達較低,在Ⅱa型比例較高的髂脛外側肌、腓骨長肌以及只含有Ⅱb型肌纖維的胸大肌、胸小肌中表達較高,且差異顯著(P<0.05)。相關性分析結果顯示,肌肉中PRKAG3基因的表達與Ⅰ型肌纖維比例呈顯著負相關(P<0.05),與Ⅱa型和Ⅱb型肌纖維比例均呈正相關,但差異不顯著(P>0.05)。提示,在雞骨骼肌中PRKAG3很可能是AMPK在白肌中發揮作用的主要γ調節亞基。
關鍵詞:PRKAG3基因;雞;肌纖維類型;表達;相關性;肌肉;AMPK
中圖分類號: S831.2 文獻標志碼: A 文章編號:1002-1302(2021)16-0144-04
一磷酸腺苷激活蛋白激酶(AMP-activated protein kinase,AMPK)是一種對能量敏感的蛋白激酶,是由一個催化亞基α、2個調節亞基β和γ構成的異源三聚體。其中催化亞基α存在α1、α2 2個亞型,調節亞基β存在β1、β2 2個亞型,調節亞基γ存在γ1、γ2、γ3這3個亞型[1]。PRKAG3(protein kinase adenosine monophosphate-activated γ3-subunit)是編碼調節亞基γ3的基因,又稱AMPKγ3。在人、大鼠和小鼠等哺乳動物中的研究表明,不同于AMPK調節亞基γ的另外2個亞型(γ1和γ2)在體內廣泛分布,PRKAG3在骨骼肌中高度特異性表達[1]。AMPKγ3基因敲除小鼠(Prkag3-/—)易疲勞,AMPK激活劑和低血糖濃度等誘導的特異AMPK應答和功能均消失,表示AMPK其他三聚體不能補償包含有γ3的AMPK三聚體在體內的作用[2-3]。自從2000年Milan等報道PRKAG3的第200個密碼子Arg突變為Gln是引起漢普夏豬酸肉效應的根本原因[4]后,PRKAG3作為影響畜禽肉品質的重要候選基因,其多態性常被用于與畜禽肉品質、屠宰性狀等經濟性狀進行關聯研究[5-9]。然而值得注意的是,PRKAG3基因的這種突變主要影響白肌纖維占很大比例的骨骼肌中AMPK的酶活性[1,10]。另有研究表明,PRKAG3基因R200Q突變豬慢肌肌球蛋白重鏈亞型比例增加[11]。可見,PRKAG3在骨骼肌肌纖維類型組成上也發揮著重要作用。
骨骼肌的肌纖維類型是影響肉品質的重要因素。根據肌纖維的形態、功能和生理生化特性,禽類將肌纖維分為慢肌纖維(紅肌纖維、Ⅰ型、氧化型肌纖維)、快紅肌纖維(Ⅱa型、快速氧化型纖維)和快白肌纖維(快速酵解型纖維、Ⅱb型)[12-13]。總體上,慢肌纖維含量多的肌肉肉色紅、嫩度高、風味好,而快白肌纖維含量多的肌肉肉色蒼白、屠宰后容易產生“水豬肉”[14-15]。目前,在雞上還未見有關PRKAG3基因表達與肌纖維類型相關的研究報道。本研究以成年隱性白羽肉雞為試驗對象,研究不同部位肌肉中PRKAG3基因的表達情況并將其與肌纖維類型進行關聯分析,研究結果將為揭示PRKAG3在雞骨骼肌中的作用以及雞的肉品質研究提供理論依據。
1 材料與方法
1.1 試驗動物
120日齡隱性白羽肉雞由江蘇省家禽科學研究所國家地方雞種資源基因庫提供。選取體質量變異系數在5%以內的隱性白羽肉雞母雞10只。每個個體分別采集胸大肌、胸小肌、縫匠肌、髂脛外側肌、腓骨長肌和比目魚肌等樣品,置于液氮速凍后,轉入-80 ℃冰箱保存。
1.2 總RNA提取和cDNA合成
按照TRNzol Universal總RNA提取試劑[DP424,天根生化科技(北京)有限公司]說明書提取肌肉樣品RNA,經瓊脂糖凝膠電泳檢測提取的RNA純度和完整性后,核酸定量儀(Thermo NanoDrop One,美國)測定其濃度。按照HiScript Ⅲ RT SuperMix for qPCR(R323-01,南京諾唯贊生物科技有限公司)說明書進行cDNA合成,合成產物保存于-20 ℃備用。
1.3 引物設計
根據GenBank中雞PRKAG3 mRNA序列(登錄號:NM__001031258.2)和DNA序列(登錄號:NC_006094.5)以及雞GAPDH mRNA序列(GenBank登錄號:NM_204305.1)和DNA序列(GenBank登錄號:NC_006088.5),運用在線Primer 3,跨基因內含子設計引物,送生工生物工程(上海)股份有限公司合成。PRKAG3基因上游引物序列為5′-GGATGCTCACCATCACTG-3′,下游引物序列為5′-TCTCCACGTCTCAATCTTGT-3′,片段大小為104 bp;GAPDH基因上游引物序列為 5′-CGATCTGAACTACATGGTTTAC-3′,下游引物序列為5′-TCTGCCCATTTGATGTTGC-3′,片段大小為153 bp。
1.4 實時熒光定量PCR
實時熒光定量PCR采用SYBR Green Ⅰ法,優化退火溫度、引物濃度、模板濃度等。反應體系為2×ChamQ SYBR Color qPCR Master Mix 10μL,50×ROX Reference Dye2 0.4 μL,上下游引物(10 μmol/L)各0.4 μL,模板2 μL,加ddH2O至20 μL。反應條件為95 ℃ 30 s;95 ℃ 10 s,60 ℃ 30 s,95 ℃ 15 s,40個循環;95 ℃ 15 s,60 ℃ 60 s,95 ℃ 15 s。每個樣品設3個重復,每次反應均設空白樣品為陰性對照。
1.5 冰凍切片制作與肌纖維類型分析
在恒溫冷凍切片機(Leica CM1850,德國)內進行肌肉樣品冰凍切片的制作,每個樣品垂直于肌纖維延展方向做橫切切片,切片厚度12 μm。切片在室溫下干燥5 min后,參考文獻[13]的方法進行肌球蛋白ATPase堿孵育法染色,染好的切片置于正置顯微鏡(Leica DM1000,德國)下觀察并拍照,每個個體隨機抽取5個完整肌束用Imag-Pro Plus 6.0軟件分析肌纖維類型。
1.6 統計分析
實時熒光定量PCR的結果采用2-△△CT法進行處理,將腓骨長肌設為對照組,計算不同肌肉組織中PRKAG3基因的相對表達量,用SPSS 20.0統計軟件中One-way Anova分析組織對PRKAG3基因表達量和肌纖維類型比例的影響,并用Bivariate Correlation分析PRKAG3基因表達量與肌纖維類型比例的相關性。所有數據以平均值±標準誤表示,P<0.05為差異顯著性水平。
2 結果與分析
2.1 不同部位肌肉的肌纖維類型組成
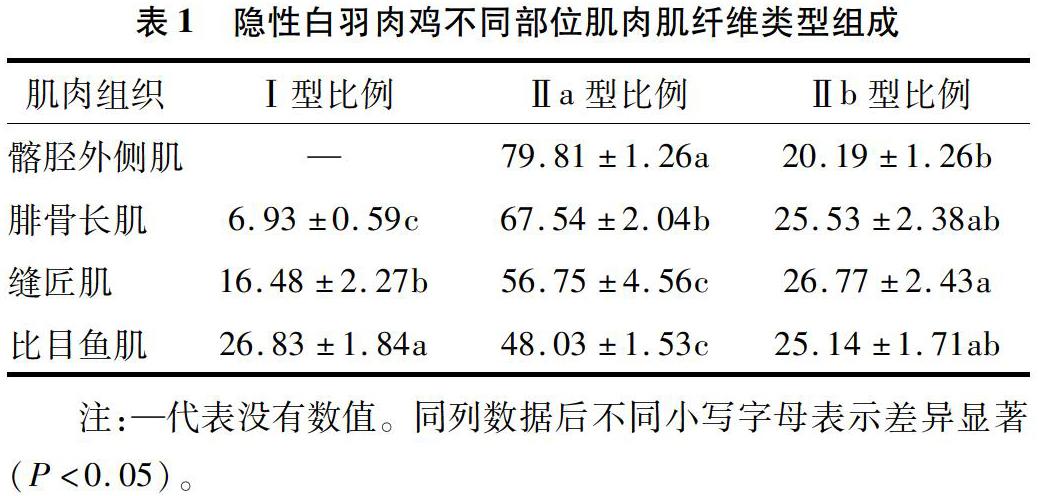
隱性白羽肉雞腓骨長肌、縫匠肌、髂脛外側肌、比目魚肌、胸大肌和胸小肌等肌肉肌纖維ATPase堿孵育法染色結果見圖1,染色淺的是ATPase活性弱的慢肌纖維(Ι型),染色深的是ATPase活性強的快白肌纖維(Ⅱb型),染色效果居中的是ATPase活性居中的快紅肌纖維(Ⅱa型)。由圖1可見,胸大肌和胸小肌中只有Ⅱb型一種肌纖維類型,髂脛外側肌中有Ⅱa和Ⅱb2種肌纖維類型,腓骨長肌、縫匠肌和比目魚肌中可以區分出Ⅰ、Ⅱa和Ⅱb等3種肌纖維類型。
對髂脛外側肌、腓骨長肌、縫匠肌和比目魚肌這4個部位肌肉肌纖維類型組成比例進行分析,結果見表1。由表1可見,比目魚肌的 Ι型肌纖維比例最高,顯著高于縫匠肌和腓骨長肌(P<0.05),縫匠肌的Ι型肌纖維比例又顯著高于腓骨長肌(P<0.05);髂脛外側肌的Ⅱa型肌纖維比例最高,顯著高于腓骨長肌、縫匠肌和比目魚肌(P<0.05),腓骨長肌的Ⅱa型肌纖維比例又顯著高于縫匠肌和比目魚肌(P<0.05),而縫匠肌和比目魚肌的Ⅱa型肌纖維比例差異不顯著(P>0.05);Ⅱb型肌纖維比例在縫匠肌中最高,在髂脛外側肌中最低,除縫匠肌與髂脛外側肌的Ⅱb型肌纖維比例差異達顯著水平外(P<0.05),其余肌肉的Ⅱb型肌纖維比例差異均不顯著(P>0.05)。
2.2 PRKAG3基因在不同部位肌肉中的表達
由圖2可見,PRKAG3基因在髂脛外側肌中表達水平最高,在比目魚肌中表達水平最低;PRKAG3基因在髂脛外側肌、腓骨長肌、胸大肌和胸小肌中的表達差異不顯著(P>0.05),但均顯著高于縫匠肌和比目魚肌(P<0.05);PRKAG3基因在縫匠肌和比目魚肌中的表達量差異不顯著(P>0.05)。
2.3 不同部位肌肉中PRKAG3基因的表達與肌纖維類型相關分析
由表2可見,肌肉中PRKAG3基因的表達與Ⅰ型肌纖維比例呈顯著負相關(P<0.05),與Ⅱa型和Ⅱb型肌纖維比例均呈正相關,但差異不顯著(P>0.05)。
3 討論與結論
肌纖維是肌肉的構成單位,肌纖維類型不僅與肉色、pH值、系水力、嫩度、肌內脂肪和風味物質含量等肉品質指標密切相關,還直接影響骨骼肌的代謝[16-20]。了解肌肉肌纖維類型分布規律對研究動物生長、代謝和肉品質均具有重要意義。不同部位的肌肉,其肌纖維類型組成往往不同[21]。本研究用ATPase堿孵育法對成年隱性白羽肉雞不同部位肌肉肌纖維類型組成進行分析,結果在位于小腿部位的腓骨長肌和比目魚肌以及位于大腿部位的縫匠肌中檢測到Ⅰ、Ⅱa和Ⅱb 3種肌纖維類型,在位于大腿部位的髂脛外側肌中檢測到Ⅱa和Ⅱb 2種肌纖維類型,而在軀干肌肉胸大肌和胸小肌中均僅檢測到Ⅱb型纖維。這與Zhang等在AA肉雞[22]以及Oshima等在絲羽、蛋用和肉用3個品種雞[23]的胸大肌中均僅檢測到Ⅱb型纖維的結果一致。
越來越多的研究表明,AMPK在控制骨骼肌葡萄糖攝取、脂肪酸氧化和基因表達方面發揮著重要作用。AMPK不同亞單位在不同類型肌纖維中的獨特表達模式,很可能與AMPK發揮作用有關[24]。在大鼠和小鼠骨骼肌中,PRKAG3基因的表達與肌肉中白肌Ⅱb型比例呈強正相關,強烈暗示PRKAG3基因在該組織中的作用[1]。束婧婷等用表達譜芯片挖掘雞骨骼肌不同類型肌纖維差異表達基因時發現,PRKAG3可能是調控雞肌纖維類型組成基因網絡中的重要節點基因[25]。本研究表明,PRKAG3基因在Ⅰ型肌纖維比例相對較高的比目魚肌和縫匠肌中表達較低,在Ⅱa型比例較高的髂脛外側肌、腓骨長肌以及只含有Ⅱb型肌纖維的胸大肌、胸小肌中表達較高,且差異達顯著水平(P<0.05)。相關性分析結果顯示,肌肉中PRKAG3基因的表達與Ⅰ型肌纖維比例呈顯著負相關(P<0.05),與Ⅱa型和Ⅱb型肌纖維比例均呈正相關,但差異不顯著(P>0.05)。提示,在雞骨骼肌中PRKAG3很可能是AMPK在白肌中發揮作用的主要γ調節亞基。
參考文獻:
[1]Mahlapuu M,Johansson C,Lindgren K,et al. Expression profiling of the gamma-subunit isoforms of AMP-activated protein kinase suggests a major role for gamma3 in white skeletal muscle[J]. American Journal of Physiology-Endocrinology and Metabolism,2004,286(2):E194-E200.
[2]Barnes B R,Glund S,Yc L,et al. 5′-AMP-activated protein kinase regulates skeletal muscle glycogen content and ergogenics[J]. Faseb Journal,2005,19(7):773-779.
[3]Barnes B R,Marklund S,Steiler T L,et al. The 5′-AMP-activated protein kinase γ3 isoform has a key role in carbohydrate and lipid metabolism in glycolytic skeletal muscle[J]. The Journal of Biological Chemistry,2004,279(37):38441-38447.
[4]Milan D,Jeon J T,Looft C,et al. A mutation in PRKAG3 associated with excess glycogen content in pig skeletal muscle[J]. Science,2000,288(5469):1248-1251.
[5]Ryan M T,Hamill R M,OHalloran A M,et al. SNP variation in the promoter of the PRKAG3 gene and association with meat quality traits in pig[J]. BMC Genetics,2012,13(1):66.
[6]DallOlio S,Scotti E,Costa L N,et al. Effects of single nucleotide polymorphisms and haplotypes of the protein kinase AMP-activated non-catalytic subunit gamma 3 (PRKAG3) gene on production,meat quality and carcass traits in Italian Large White pigs[J]. Meat Science,2018,136:44-49.
[7]焦 斐. 牦牛PRKAG3、FAS基因多態性與肉質相關性研究[D]. 蘭州:甘肅農業大學,2012.
[8]王 華,魏玉兵,童建偉,等. 綿羊 PRKAG3基因 SNPs 與肉質性狀的相關性分析[J]. 華北農學報,2016,31(1):134-140.
[9]周娜娜. 優質雞PRKAG3基因單核苷酸多態性與屠宰性狀和肉質性狀的相關研究[D]. 雅安:四川農業大學,2008:44-48.
[10]Lebret B,Le Roy P,Monin G,et al. Influence of the three RN genotypes on chemical composition,enzyme activities,and myofiber characteristics of porcine skeletal muscle[J]. Journal of Animal Science,1999,77(6):1482-1489.
[11]Park S K,Gunawan A M,Scheffler T L,et al. Myosin heavy chain isoform content and energy metabolism can be uncoupled in pig skeletal muscle[J]. Journal of Animal Science,2009,87(2):522-531.
[12]Bandman E,Rosser B W. Evolutionary significance of myosin heavy chain heterogeneity in birds[J]. Microscopy Research and Technique,2000,50(6):473-491.
[13]單艷菊,束婧婷,胡 艷,等. 不同鴨種肌纖維早期發育特性及其與骨骼肌生長發育的相關性研究[J]. 農業生物技術學報,2014,22(2):225-231.
[14]李伯江,李平華,吳望軍,等. 骨骼肌肌纖維形成機制的研究進展[J]. 中國農業科學,2014,47(6):1200-1207.
[15]Wang X F,Li J L,Cong J H,et al. Preslaughter transport effect on broiler meat quality and post-mortem glycolysis metabolism of muscles with different fiber types[J]. Journal of Agricultural and Food Chemistry,2017,65(47):10310-10316.
[16]Klont R E,Brocks L,Eikelenboom G. Muscle fibre and meat quality[J]. Meat Science,1998,49(S1):S219-S229.
[17]Lee S H,Joo S T,Yc R. Skeletal muscle fiber type and myofibrillar proteins in relation to meat quality[J]. Meat Science,2010,86(1):166-170.
[18]Lee S H,Choe J H,Choi Y M,et al. The influence of pork quality traits and muscle fiber characteristics on the eating quality of pork from various breeds[J]. Meat Science,2012,90(2):284-291.
[19]Kim G D,Yc R,Jy J,et al. Relationship between pork quality and characteristics of muscle fibers classified by the distribution of myosin heavy chain isoforms[J]. Journal of Animal Science,2013,91(11):5525-5534.
[20]劉 冰. 不同品種雞的肌纖維發育規律及其與肉質的關系[D]. 北京:中國農業大學,2005:7-8.
[21]Ariano M,Armstrong R B,Edgerton V R. Hindlimb muscle fiber populations of five mammals[J]. The Journal of Histochemistry and Cytochemistry,1973,21(1):51-55.
[22]Zhang L,Yue H Y,Zhang H J,et al. Transport stress in broilers:I. Blood metabolism,glycolytic potential,and meat quality[J]. Poultry Science,2009,88(10):2033-2041.
[23]Oshima I,Iwamoto H,Tabata S,et al. Comparative observations on the growth changes of the histochemical property and collage architecture of the musculus puboischio-femoralis pars medialis from silky,layer-type and meat-type cockerels[J]. Animal Science Journal,2008,79(2):255-269.
[24]Winder W W,Hardie D G,Mustard K J,et al. Long-term regulation of AMP-activated protein kinase and acetyl-CoA carboxylase in skeletal muscle[J]. Biochemical Society Transactions,2003,31(1):182-185.
[25]束婧婷,姬改革,單艷菊,等. 基于表達譜芯片挖掘雞骨骼肌不同類型肌纖維的差異表達基因[J]. 中國農業科學,2017,50(14):2826-2836.
猜你喜歡
藝術科技(2016年9期)2016-11-18 18:24:22
藝術科技(2016年9期)2016-11-18 16:48:09
校園英語·上旬(2016年10期)2016-11-16 19:03:00
人間(2016年28期)2016-11-10 00:08:22
文藝生活·中旬刊(2016年10期)2016-11-04 05:28:04
青年文學家(2016年27期)2016-11-02 17:46:42
科技視界(2016年21期)2016-10-17 17:37:34
中國實用醫藥(2016年24期)2016-10-17 04:31:12
中國實用醫藥(2016年24期)2016-10-17 03:37:40
中國實用醫藥(2016年24期)2016-10-17 03:35:06
