杉木(Cunninghamia lanceolate)人工林生長狀況與根系生物量相關(guān)性研究
2021-09-09 04:47:10廖迎春段洪浪施星星孟慶銀劉文飛沈芳芳樊后保朱濤
生態(tài)環(huán)境學(xué)報(bào) 2021年6期
廖迎春 ,段洪浪 ,施星星,孟慶銀,劉文飛 ,沈芳芳 ,樊后保 *,朱濤
1. 南昌工程學(xué)院水利與生態(tài)工程學(xué)院,江西 南昌 330099;2. 江西省退化生態(tài)系統(tǒng)修復(fù)與流域生態(tài)水文重點(diǎn)實(shí)驗(yàn)室,江西 南昌 330099;3. 福建沙縣官莊國有林場,福建 沙縣 365050
根系是植物吸收養(yǎng)分和水分的重要器官,在土壤養(yǎng)分、水分以及碳循環(huán)方面扮演著重要角色(McCormack et al.,2012)。根系生態(tài)過程是森林生態(tài)系統(tǒng)能量流動和物質(zhì)循環(huán)的關(guān)鍵環(huán)節(jié),影響和決定著從植物個體到生態(tài)系統(tǒng)等尺度上眾多的生理、生態(tài)過程(Vogt et al.,1996;Antoninka et al.,2009;Gambetta et al.,2013)。在植物根系中,著生在根系分枝前端的 1—2級根直徑小、根長短、比根長高(Pregitzer et al.,1997)、數(shù)量龐大(Pregitzer et al.,2002)、表面積高(Guo et al.,2004),是根系吸收養(yǎng)分和水分的主要部位(Guo et al.,2008;Pregitzer,2008),為共生真菌侵染的主要場所(Eissenstat et al.,1999;Guo et al.,2008),是根系最主要的吸收根(Guo et al.,2008;Pregitzer,2008)。運(yùn)輸細(xì)根(3—5級根)和粗根(5級以上)在根的生物量中占主要地位,是養(yǎng)分運(yùn)輸和碳水化合物的主要貯藏器官,但由于表面積小、活性低、皮層脫落、無菌根,因此吸收水分和養(yǎng)分的能力以及根土相互作用較弱(Eissenstat,1992)。
在不同環(huán)境條件下,植物能夠通過調(diào)整根系生物量以提高對土壤資源的吸收(Finér et al.,2007;Helmisaari et al.,2007;Ostonen et al.,2011)。例如:當(dāng)土壤資源受到限制時,樹木會分配更多生物量到細(xì)根(Vogt et al.,1996;Finér et al.,1998;Helmisaari et al.,2007)以獲取土壤資源。Blume-Werry et al.(2017)對北極苔原植物進(jìn)行研究發(fā)現(xiàn)隨海拔升高,植物的生物量降低,細(xì)根生物量增加;Sprunger et al.(2017)對不同種植方式的作物進(jìn)行研究發(fā)現(xiàn)間作方式的農(nóng)作物細(xì)根生物量占植株生物量的比例高于單作方式的農(nóng)作物;Comas et al.(2013)發(fā)現(xiàn)在干旱條件下雷斯克勒(Physaria)和水稻(Oryza)兩種作物通過減小根直徑、增加比根長和根長密度使根向深層土壤延伸以維持植株的生產(chǎn)力。以上研究結(jié)果說明植物根系生物量分配與外界環(huán)境密切相關(guān),根系生物量的大小在一定程度上反映了植物對環(huán)境的適應(yīng)能力(Rivero et al.,2007;Schachtman et al.,2008),并且與植物生產(chǎn)力存在一定的關(guān)系,即樹木生長存在差異,細(xì)根生物量也可能存在差異(徐偉強(qiáng)等,2016)。因此,開展根系生物量及其與林分生長狀況相關(guān)性研究,對進(jìn)一步認(rèn)知植物對環(huán)境的適應(yīng),深化根系生態(tài)學(xué)具有重要科學(xué)意義。
杉木(Cunninghamia lanceolata)是中國亞熱帶地區(qū)重要造林樹種,在中國南方地區(qū)分布較廣,在林業(yè)生產(chǎn)中占有重要地位。然而由于立地條件類型未做到適地適樹,非高產(chǎn)區(qū)大面積造林(孫長忠等,2001)以及長期連栽導(dǎo)致的土壤養(yǎng)分下降造成了大面積低產(chǎn)低效林的產(chǎn)生(O’Hehir et al.,2010;陳龍池等,2004;劉福德等,2005)。鑒于此,本研究在杉木分布的主要地區(qū)——福建沙縣,以杉木中齡級(10—15年生)人工林為對象,研究杉木人工林生長狀況與根系生物量關(guān)系。由于林分生物量是林分生產(chǎn)力的重要指標(biāo),體現(xiàn)了林分環(huán)境質(zhì)量的優(yōu)劣(謝建文,2020),因此本研究選擇林分生物量為“高”(CH)、“中”(CM)、“低”(CL)的 3 個杉木人工林進(jìn)行野外調(diào)查和取樣,研究杉木不同組分根系的生物量密度、土壤C含量、土壤N含量、土壤C?N以及根系生物量與林分生物量的關(guān)系。本研究將對完善樹木適應(yīng)環(huán)境機(jī)制理論、指導(dǎo)杉木人工林精準(zhǔn)施肥以及加強(qiáng)中國人工林生態(tài)系統(tǒng)管理具有重要意義。
1 材料與方法
1.1 研究區(qū)概況
試驗(yàn)樣地位于福建省沙縣官莊國有林楊(26°30′47″N,117°43′29″E)。試驗(yàn)地海拔為 200 m,氣候?yàn)榈湫蛠啛釒Ъ撅L(fēng)氣候,多年平均氣溫20.6 ℃,年平均降雨量 1653 mm(數(shù)據(jù)來源于中國氣象數(shù)據(jù)網(wǎng) 1961—2019年氣象數(shù)據(jù),http://data.cma.cn/data/cdcdetail/dataCode/A.0029.00 05.html),土壤類型為山地紅壤。
標(biāo)準(zhǔn)地設(shè)置:2019年10月,在7個12—14年生杉木人工林分別設(shè)置3個20 m×20 m標(biāo)準(zhǔn)地,標(biāo)準(zhǔn)地總數(shù)為:3個重復(fù)×7種林分=21個標(biāo)準(zhǔn)地。對每個標(biāo)準(zhǔn)地進(jìn)行林分調(diào)查,記錄株數(shù)、胸徑以及林下植被種類和蓋度,并用相對生長模型估算杉木人工林生物量(方精云等,1996;劉琪璟等,2017)。林分生物量計(jì)算方法如下:

式中:
B 為林分生物量(Stand biomass,t·hm?2);
V 為林分材積量(Forest volume,m3·hm?2)。

式中:
V 為林分材積量(Forest volume,m3·hm?2);
D 為標(biāo)準(zhǔn)地人工林密度(Density,trees·hm?2)。

式中:
Vi(單木材積量)通過胸徑(Diameter at breast height,DBH-i,cm)數(shù)據(jù)和福建三明市杉木一元立木材積表估算得出(劉琪璟等,2017),公式如下:

根據(jù)7個林分的調(diào)查結(jié)果,最終選擇了林分生物量差異顯著(P<0.05)的3個人工林作為研究對象,并根據(jù)林分生物量的大小分別標(biāo)記為“高”、“中”、“低”(CH、CM、CL)。3個林分的林下植被均很稀少,主要類型有五節(jié)芒(Miscanthus floridulus)、芒萁(Dicranopteris olichotoma)、蕨類(Pteridum aquilinum var. latiusculum),蓋度在8%—10%左右。林分基本概況見表1。

表1 供試林地的基本概況Table 1 Characteristics of the sampling sites
1.2 樣品采集和分析方法
樣品采集:2020年生長季中后期(8月),在選定的各個標(biāo)準(zhǔn)地采用相同的方法進(jìn)行取樣。用土鉆法(內(nèi)徑8 cm,高10 cm)對根系和土壤進(jìn)行取樣。由于0—40 cm土層的細(xì)根占杉木細(xì)根總生物量的80%以上(Liao et al.,2014;陳光水等,2004;張雷等,2009;匡冬姣等,2015),因此取樣深度定為40 cm。每個標(biāo)準(zhǔn)地內(nèi)隨機(jī)選取8個取樣點(diǎn)(避開杉木主根和側(cè)根)用土鉆進(jìn)行取樣,每個取樣點(diǎn)分為3層(淺層土0—10 cm、中層土10—20 cm和深層土20—40 cm),共取根系樣品數(shù)量為:3個林分×3個標(biāo)準(zhǔn)樣地×8個取樣點(diǎn)×3個土層=216個。取樣時將土鉆取得的各層土柱樣品(包括土壤與根系)放在塑料布上,仔細(xì)去掉根系上的土壤(要特別注意保持根段的完整性),將取出根段放入己編號封口袋,立即放入溫度為 0—4 ℃的冷藏箱內(nèi),運(yùn)回實(shí)驗(yàn)室放入?40 ℃冰箱冷凍保存以盡可能減少對根系組織和離子含量的影響(Clemensson-Lindell et al.,1992)。在采集根系樣品的同時,對各土鉆內(nèi)去除根系后的土壤進(jìn)行取樣,共取土壤樣品為216個。土壤樣品晾干研磨過100目篩,用于土壤C、N的測定。
根系分級:在實(shí)驗(yàn)室進(jìn)行,用低溫去離子水輕輕洗去每個根系樣品表面殘留的土壤顆粒及其他雜質(zhì),放入盛有低溫(2—4 ℃)去離子水的玻璃皿上,根據(jù)根段顏色和外形等特征區(qū)分出杉木根系和灌草根根系。在挑選出的杉木根系中,按照Fitter(1985)根系序列位置命名方法對完整的根段進(jìn)行分級,即位于根軸最遠(yuǎn)端沒有分枝最細(xì)小的根稱為1級根,1級根著生在2級根上,依次類推,對于不完整的根段,則和已分好類別的根段進(jìn)行對比,進(jìn)行歸類,將每個土鉆中取得的根系分為杉木吸收根(1—2級細(xì)根,以下簡稱吸收根)、杉木運(yùn)輸細(xì)根(3—5級細(xì)根,以下簡稱運(yùn)輸細(xì)根)、杉木粗根(5級以上,以下簡稱粗根)和灌草根(林地中灌木和草本根系,不區(qū)分根級和種類)
根系生物量密度:將不同標(biāo)準(zhǔn)地土鉆中分離出的吸收根、運(yùn)輸細(xì)根、粗根和灌草根于 65 ℃下烘干至恒定質(zhì)量,然后用電子天平(+0.0001 g)稱質(zhì)量,分別計(jì)算出吸收根、運(yùn)輸細(xì)根、粗根和灌草根生物量密度。計(jì)算方法如下:

式中:
DRB為根系生物量密度(g·cm?3);
BR為土鉆中根系生物量(g);
VC為土鉆體積(cm3)。

式中:
B′為根系生物量(kg·hm?2);
DRB為根系生物量密度(g·cm?3);
H為土層深度(cm);
S為標(biāo)準(zhǔn)地面積(cm2×108)(以1 hm2計(jì)算)。

土壤C、N測定:利用杜馬斯高溫燃燒法進(jìn)行測定。將土壤樣品烘干磨細(xì)并通過100目篩后,利用元素分析儀(VarioMicrocube;Elementar,Hanau,Germany)測定土壤全C和全N,并計(jì)算土壤C?N。
1.3 數(shù)據(jù)分析
使用單因素方差分析(One-way ANOVA)檢驗(yàn)胸徑、林分生物量、土壤C含量、土壤N含量、土壤C?N、根系生物量密度、根系生物量的差異性。顯著性水平設(shè)置為P<0.05。以上分析均使用 SPSS 16.0統(tǒng)計(jì)軟件完成。利用SigmaPlot 12.5分析吸收根、運(yùn)輸細(xì)根、粗根和灌草根生物量與林分生物量的相關(guān)性。文中的圖均用SigmaPlot 12.5進(jìn)行繪制。
2 結(jié)果與分析
2.1 土壤 C、N、C?N
土壤 C、N含量隨土壤深度增加而顯著降低(P<0.05)。表層土(0—10 cm)、中層土(10—20 cm)和深層土(20—40 cm)土壤C含量分別在1.91%—2.08%、1.22%—1.41%和0.78%—0.96%之間;土壤 N含量在 0.12%—0.13%、0.08%—0.09%和0.06%—0.08%之間(圖1)。土壤C、N在各土層均表現(xiàn)為 CH>CL(P<0.05)。土壤 C?N表現(xiàn)為隨土層加深而降低,在0—10 cm土層最高,大小在15.8—16.4之間,顯著高于10—20 cm和20—40 cm土層(P<0.05)。表層土(0—10 cm)和中層土(10—20 cm)土壤C?N不同林分間并無顯著差異,而在深層土壤(20—40 cm)中CL下降明顯,顯著低于表層土(0—10 cm)和中層土(10—20 cm)(P<0.05),以及低于CH和CM的相同土層(20—40 cm)(P<0.05)(圖 1)。

圖1 土壤C,N,C?NFig. 1 Soil C, soil N and soil C?N
2.2 根系生物量密度
杉木根系主要分布在表層土壤。各林分 0—20 cm土壤中吸收根、運(yùn)輸細(xì)根和粗根生物量密度分別占0—40 cm土層的84.2%—85.9%、84.6%—85.2%和78.6%—80.0%(表2)。不同組分根系生物量密度在0—10 cm和10—20 cm土層之間并無顯著差異,但在20—40 cm土層顯著下降(表2)。從不同林分來看,吸收根生物量密度在0—10 cm和10—20 cm土層表現(xiàn)為CH>CL(P<0.05),但在20—40 cm土層各林分間無顯著差異;運(yùn)輸細(xì)根生物量密度在0—10 cm表現(xiàn)為CH>CL(P<0.05),而在10—20 cm和20—40 cm土層各林分間無顯著差異;粗根生物量密度在0—10 cm、10—20 cm和20—40 cm土層各林分均無顯著差異。由于林下植被稀少,灌草根生物量密度在各林分中均很低,大小僅在 0.04—0.07 g·cm?3,不同林分和不同土層間均無顯著差異。
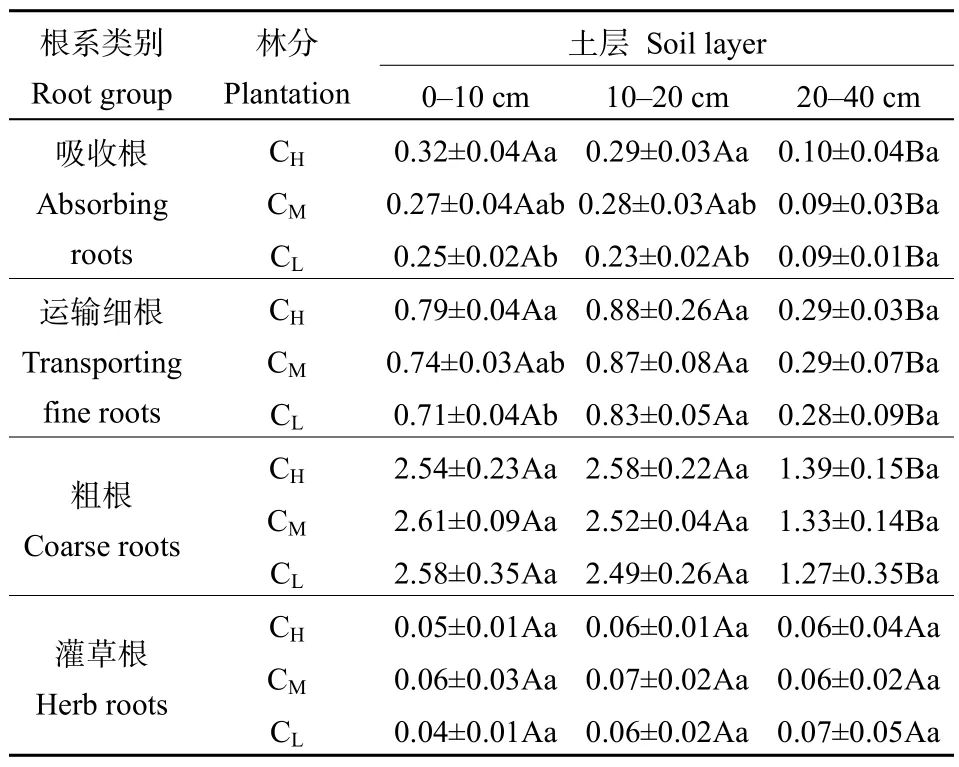
表2 根系生物量密度Table 2 Root biomass density mg·cm?3
2.3 根系生物量密度分配
從不同組分的根系來看,粗根生物量密度最大,在總根生物量密度中的比例隨著土層的深度增加而增大(圖2)。0—10 cm土層中,粗根生物量密度在總根生物量密度中所占比例為68.6%—72.0%,10—20 cm土層中所占比例為67.4%—68.9%。而在20—40 cm土層中,粗根生物量密度占總根生物量密度比例上升為74.5%—75.5%。吸收根生物量密度在總根生物量密度中所占比例較低,且隨著土層深度的增加所占比例有減小的趨勢:0—10 cm土層,吸收根生物量密度在各組分的根中所占比例為 6.9%—8.7%;在10—20 cm土層中為6.4%—7.7%;而在20—40 cm土層,吸收根生物量密度所占比例下降到5.0%—5.4%。灌草根生物量密度占總根生物量密度的比例最低,僅為1.2%—4.0%。
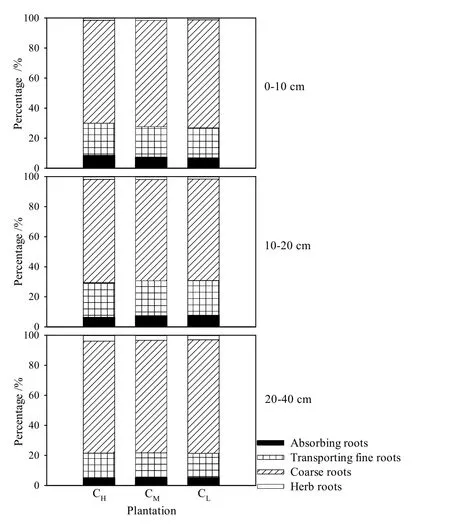
圖2 根系生物量密度分配Fig. 2 Root mass density distribution
從不同林分來看,各組分的根所占總根生物量密度的比例無顯著差異。
2.4 根系生物量
從林分角度來看,各組分根系生物量大小表現(xiàn)為粗根 (7614.8—7898.7 kg·hm?2)>運(yùn)輸細(xì)根 (2108.5—2260.6 kg·hm?2)>吸收根 (649.3—812.8 kg·hm?2)>灌草根 (229.0—252.9 kg·hm?2)。杉木吸收根生物量表現(xiàn)為CH顯著高于CL(P<0.05)(圖3),而運(yùn)輸細(xì)根、粗根和灌草根生物量不同林分間差異并不顯著。

圖3 根系生物量(B ′)Fig. 3 Root biomass
2.5 根系生物量與林分生物量相關(guān)性研究
將0—40 cm土層根系生物量和林分生物量進(jìn)行回歸分析發(fā)現(xiàn):不同組分的根系中,吸收根生物量與杉木人工林生物量呈顯著正相關(guān)關(guān)系(P<0.05)(圖 4),也就是林分生物量越大,其吸收根生物量也越高。但是運(yùn)輸細(xì)根、粗根和灌草根生物量與林分生物量之間并無顯著相關(guān)性。

圖4 根系生物量與林分生物量相關(guān)性Fig. 4 Linear regressions of root biomass and stand biomass
3 討論
3.1 土壤C、N的變化
樹木的生長與外界環(huán)境密切相關(guān)。土壤是根系直接接觸到的環(huán)境,其養(yǎng)分含量能夠直接對樹木根系生長產(chǎn)生調(diào)控作用,決定植物根系碳水化合物的分配(Hendrick et al.,1993),是影響樹木生長的重要因素(Yuan et al.,2010;程云環(huán)等,2005)。本研究對林分生物量不同的杉木人工林土壤C、N含量進(jìn)行分析發(fā)現(xiàn)林分生物量“高”的林分,其土壤C、N含量也相對較高,表現(xiàn)在表層土(0—10 cm)、中層土(10—20 cm)和深層土(20—40 cm)的土壤C、N含量均顯著高于林分生物量低的杉木林。然而在林分生物量不同的杉木林中,土壤C?N在表層土(0—10 cm)和中層土(10—20 cm)各林分間差異并不顯著,但在深層土(20—40 cm)分化明顯,主要表現(xiàn)為林分生物量“低”的林分,土壤C?N顯著高于林分生物量“中”和“高”的林分。這可能是由于林分生物量“低”的林分,其土壤N尤其是深層土壤N含量較低造成C?N較高,而相對較高的C?N反而會使微生物在分解有機(jī)質(zhì)的過程中存在氮限制,不利于植物的生長(王建林等,2014),造成林分生物量偏低。
3.2 根系生物量密度
在不同林分中,林分生物量“高”的林分,吸收根,運(yùn)輸細(xì)根的生物量密度也相對較高,尤其是在表層土壤(0—10 cm)和中層土壤(10—20 cm)更為顯著,表現(xiàn)為CH>CL(表2)。這個結(jié)果說明生物量高的林分,其土壤C、N含量也更高,樹木生長更好,但同時需要更多的吸收根吸收土壤水分和養(yǎng)分,更多運(yùn)輸細(xì)根和粗根輸導(dǎo)水分和養(yǎng)分以及固定植物體(紀(jì)嬌嬌等,2020)。該結(jié)論與前人研究結(jié)果一致(Vicca et al.,2012;陳勁松等,2001;紀(jì)嬌嬌等,2020)。Vicca et al.(2012)曾對全球49個林分進(jìn)行研究,發(fā)現(xiàn)土壤養(yǎng)分較高的林分會分配更多的生物量到地上部分。在養(yǎng)分及環(huán)境適宜的條件下,樹木會通過增加吸收根的生物量來實(shí)現(xiàn)生長的需求(陳勁松等,2001),同時也需要更多的運(yùn)輸細(xì)根和粗根輸導(dǎo)水分和養(yǎng)分以及固定植物體,林分生物量也更高(紀(jì)嬌嬌等,2020)。然而本研究結(jié)果與Helmisaari et al.(2007)的研究結(jié)果相反,他們通過對分布在芬蘭不同地區(qū)的8個55—140年生的挪威云杉Norway spruce(Piceaabies L. Karst.)和8個歐洲赤松(Pinussylvestris L.)林分的細(xì)根生物量和土壤養(yǎng)分關(guān)系進(jìn)行研究發(fā)現(xiàn)土壤氮有效性越低的地區(qū),細(xì)根生物量越高。研究結(jié)果的差異可能是因?yàn)镠elmisaari et al.(2007)的研究對象是分布在不同地區(qū)、不同年齡階段的樹種,在不同生長階段樹木養(yǎng)分和水分的運(yùn)轉(zhuǎn)狀況和環(huán)境因子存在差異(孔令侖等,2017),導(dǎo)致了研究結(jié)果可能不一致。
根系的分布還受到土壤水分、養(yǎng)分以及溫度等因素的影響(蔡瑛瑩等,2018)。本研究中,吸收根和運(yùn)輸細(xì)根主要分布在表層(0—10 cm)和中層(10—20 cm)土壤,在深層土壤(20—40 cm)的分布有所減少。盡管杉木粗根在深層土壤(20—40 cm)的生物量密度降低,其在總根生物量密度中所占比例卻隨土壤深度的增加而增大(圖2)。這可能是由于表層和中層土壤養(yǎng)分含量相對較高,吸收根更容易獲得足夠的養(yǎng)分供樹木生長(Zhou et al.,2007;黃林等,2012;王福根等,2020)。而隨著土層深度增加,土壤養(yǎng)分含量降低,進(jìn)而影響吸收根吸收養(yǎng)分和水分,運(yùn)輸細(xì)根和粗根輸送和貯存養(yǎng)分,因此吸收根、運(yùn)輸細(xì)根和粗根生物量密度在深層土壤(20—40 cm)顯著降低(Cormier et al.,2015)。根系的這種分布特征可能與植物地下部分對環(huán)境的適應(yīng)策略有關(guān),它們通過分配生物量至土壤表層細(xì)根(Freschet et al.,2015)和土壤深層的粗根,以便更有效地獲得養(yǎng)分(McCormack et al.,2020)和固定植物體。
3.3 根系生物量與林分生物量的關(guān)系
關(guān)于植物地上和地下部分,很早就有學(xué)者提出相關(guān)理論。Brouwer(1963)提出功能性均衡(Functional equilibrium)理論,認(rèn)為高等植物的地下和地上部分存在相關(guān)性。本研究通過對同一地區(qū)林分生物量差異顯著的杉木林生物量和不同組分根系生物量比較研究發(fā)現(xiàn)林分生物量較高的杉木人工林根系生物量也較高。在不同組分的根系中,吸收根與林分生物量呈顯著正相關(guān)關(guān)系,其生物量隨林分生物量的增加而增加(圖4)。這可能是由于根枝末端的吸收根承擔(dān)了根系的主要吸收功能(Xia et al.,2010),也是受土壤環(huán)境影響反應(yīng)最為敏感的部分(Eissenstat et al.,2015),為了應(yīng)對環(huán)境條件的改變,吸收根生物量會發(fā)生變化(Zadworny et al.,2016)以適應(yīng)環(huán)境。吸收根的這些變化即使很微小,也可能會影響植物的生長(江洪等,2016),導(dǎo)致地上生產(chǎn)力發(fā)生較大變化(Ostonen et al.,2011)。由于本研究中的杉木人工林在同一地區(qū),具有相同的氣候條件,因此造成林分生物量差異的原因很可能是土壤養(yǎng)分的不同導(dǎo)致。本研究結(jié)果也證實(shí)了土壤養(yǎng)分-根系-植株之間具有相互依存,互相牽制的關(guān)系。
4 結(jié)論
在同一地區(qū),土壤養(yǎng)分是造成根系生物量和林分生物量存在差異的一個主要原因,土壤養(yǎng)分好的林分,其根系生物量也較高,同時擁有較高的林分生物量。在根系中,吸收根對環(huán)境條件的響應(yīng)最為敏感,吸收根生物量大的林分,其林分生物量也更高。
