LncRNAMEG8影響結腸癌細胞惡性進展
2021-09-08 05:53:45徐王彥劉忠臣
安徽理工大學學報·自然科學版 2021年3期
關鍵詞:結腸癌
徐王彥 劉忠臣


摘 要:目的 探究lncRNA MEG8對結腸癌細胞增殖、侵襲和凋亡的作用,闡明其作用機制。方法 利用實時熒光定量PCR(Real Time-Quantitative PCR,RT-qPCR)檢測MEG8和miR-1827表達;Western blotting和CCK-8檢測蛋白表達和細胞增殖;Transwell和流式細胞術檢測細胞侵襲和凋亡;雙熒光素酶報告基因試驗和RNA免疫沉淀反應(RNA Immunoprecipitation,RIP)驗證MEG8與miR-1827結合。結果 MEG8通過結合miR-1827負調控miR-1827表達。與人正常結腸上皮細胞相比,MEG8在結腸癌細胞中表達下調,而miR-1827表達上調。過表達MEG8或干擾miR-1827抑制HT29細胞增殖和侵襲,促進凋亡。沉默miR-1827逆轉干擾MEG8誘導的細胞增殖和侵襲上升,凋亡和細胞外調節蛋白激酶(extracellular regulated protein kinases,ERK)/ c-Jun氨基末端激酶(c-Jun N-terminal kinase,JNK)通路活性下降。結論 LncRNA MEG8通過下調miR-1827,促進ERK/JNK通路活性,阻礙結腸癌細胞HT29增殖和侵襲,促進凋亡。
關鍵詞:結腸癌;lncRNA MEG8;miR-1827;增殖;凋亡;侵襲
Abstract:Objective To explore the role of lncRNA MEG8 in the proliferation, invasion and apoptosis of colon cancer cells and the possible molecular mechanisms involved. Methods Real Time-Quantitative PCR (RT-qPCR)was used to detect the expression of MEG8 and miR-1827. Western blotting and CCK-8 assays were used to detect protein levels and cell proliferation. Transwell assay and flow cytometry were performed to measure cell invasion and apoptosis. Dual luciferase reporter gene and RNA Immunoprecipitation (RIP)assays were used to verify the relationship between MEG8 and miR-1827. Results MEG8 negatively regulated miR-1827 expression by binding with miR-1827. MEG8 was down-regulated in colon cancer cells, while miR-1827 was up-regulated, compared with their expression in normal human colon epithelial cells. The overexpression of MEG8 or interference to miR-1827 inhibited HT29 cells proliferation and invasion and promoted their apoptosis,while the silence of miR-1827 would reverse such MEG8-induced effects and decrease the pathway activity of ERK (extracellular regulated protein kinases)/JNK (c-Jun N-terminal kinase). Conclusion LncRNA MEG8 will increase the activity of ERK/JNK pathway with miR-1827 down-regulaed, thereby inhibiting the proliferation and invasion of the HT29 colon cancer cells, and promoting their apoptosis.
Key words:colon cancer; LncRNA MEG8; miR-1827; proliferation; apoptosis; invasion
結直腸癌是世界上第三種最常見的癌癥,占人類癌癥相關死亡人數的1/10。由于其發展迅速且隱蔽,每年新增結腸癌人數超過12萬,死亡率超過33%[1-2]。盡管診斷和治療策略不斷進步,但結腸癌患者的預后在過去十年間未發生顯著變化[3]。目前只有少數生物標記物可用于結腸癌的預防,因此,鑒定早期診斷結腸癌的分子標志物具有重要意義。
長非編碼RNA(LncRNA)是長度超過200nt但沒有蛋白質編碼能力的RNA分子。LncRNA可以作為腫瘤中的癌基因或抑癌基因,參與腫瘤增殖、轉移、自噬和凋亡等調控[4]5 752。多種lncRNA異常與結腸癌進展密切相關。據報道,與正常結腸組織相比,母系表達基因8(matemally expressed gene 8,MEG8)在結腸癌組織中表達下調,但其參與結腸癌發病的潛在機制尚不清楚[5]。MicroRNA(miRNAs)是一類約22個核苷酸的非編碼小RNA,其失調與多種生物學過程,包括細胞增殖,分化和凋亡相關。前人研究表明,miR-1827在結腸癌組織中上調[6]5 107,但其對結腸癌細胞增殖、侵襲及凋亡的作用還有待探究。
本研究主要探討lncRNA MEG8對結腸癌細胞增殖、侵襲和凋亡的影響,并闡明其涉及的主要分子機制。
1 材料方法
(1)細胞與試劑 NCM460、HT29、CACO2、SW480、HCT116、HEK293T細胞均購自上海生命科學研究院;Magna RIP試劑盒購自美國Millipore公司;NC siRNA、MEG8 siRNA、miR-1827mimic、NC mimic、miR-1827 inhibitor、NC inhibitor均由上海吉瑪公司合成;細胞凋亡試劑盒購自中國上海聯科生物公司;CCK-8試劑盒購自美國Abcam公司;TRIzol試劑盒購自美國Invitrogen公司;反轉錄和RT-qPCR試劑盒購自中國大連Takara公司; 總的和磷酸化的細胞外調節蛋白激酶1/2(t-ERK1/2 和p-ERK1/2)、 總的和磷酸化的c-Jun氨基末端激酶(t-JNK和p-JNK)抗體購自美國CST公司;流式細胞儀和酶標儀購自德國Thermo Fisher和美國Bio-Rad公司;凝膠成像儀和CO2細胞恒溫培養箱購自美國CARESTREAM Gel Logic和德國Thermo scientific公司。
(2)細胞培養和轉染 將NCM460、HT29、CACO2、SW480、HCT116、HEK293T細胞置于含10% 胎牛血清的DMEM培養基中,并于37℃,含5% CO2的恒溫培養箱中培養。將細胞接種于6孔板,待密度達到60%,使用LipofectamineTM3000轉染試劑按照說明書進行轉染。
(3)RNA免疫沉淀反應(RNA Immunoprecipitation,RIP)將HT29細胞裂解液與包含磁珠(磁珠與anti-Ago2或IgG的抗體偶聯)的RIP緩沖液在4℃下孵育過夜。用蛋白酶K消化免疫沉淀物以純化沉淀的RNA,再用qPCR分析免疫沉淀的RNA。
(4)雙熒光素酶報告基因試驗 使用StarBase v2.0預測MEG8與miR-1827的結合位點。使用psiCHECK-2載體構建野生型和突變型psiCHECK-2-MEG8(MEG8-Wt、MEG8-Mut)載體。將HEK293T細胞接種于24孔板,并將MEG8-Wt(或MEG8-Mut)分別與miR-1827mimic或NC mimic共轉染。24h后,按照說明書的操作檢測熒光素酶活性。
(5)細胞凋亡 使用Annexin V-FITC /碘化丙啶(PI)凋亡檢測試劑盒進行凋亡分析。簡言之,細胞處理完成后收集細胞,用PBS洗滌并重懸細胞,隨后依次添加5μLAnnexin V-FITC和5μLPI。使用流式細胞儀檢測細胞凋亡。
(6)細胞增殖 將處理后的細胞接種于96孔板,并分別培養24、48或72h。在每個時間點添加10μL CCK-8溶液,并將細胞繼續孵育2h。使用酶標儀測量450nm處的吸光度值。
(7)細胞侵襲 將人工基底膜涂于Transwell室底膜上腔表面,并置于37℃放置30min。將細胞(2×105個)懸浮于500μL無血清培養基中,并添加到上腔室中。在下腔室中添加含10%胎牛血清的RPMI-1640培養基。溫育36h后,將下腔室中的細胞用4%多聚甲醛固定并用結晶紫染色。選擇5個隨機區域計算侵襲細胞數量并計算平均值。
(8)Western blotting 使用RIPA裂解液提取細胞總蛋白質,使用BCA試劑盒測量蛋白質濃度。通過10%SDS-PAGE分離蛋白質樣品,然后電轉至PVDF膜。將膜在室溫用5%脫脂奶粉封閉
2h后,與稀釋后的一抗在4℃孵育過夜。隨后將膜與辣根過氧化物酶標記的二抗在室溫孵育1h。在ChemiDoc XRS成像系統中檢測蛋白質條帶。使用Image J軟件進行條帶灰度分析。
(9)實時熒光定量PCR(Real Time-Quantitative PCR,RT-qPCR)采用TRIzol提取細胞總RNA,進行反轉錄。使用SYBR PrimeScript RT-PCR試劑盒檢測mRNA表達。反應條件設計如下:95℃,3min,進行35個循環,94℃,15s;55℃,25s;72℃,30s。mRNA相對表達通過2-ΔΔCT方法計算。
(10)統計學分析 采用SPSS 22.0軟件進行統計分析。測量的數據表示為均數±標準差,組間數據比較采用學生t檢驗和雙向方差分析。P<0.05視為差異有統計學意義。
2 結果
(1)過表達MEG8抑制HT29細胞惡性進展。如圖1所示。與人正常腸上皮細胞(NCM460)相比,結腸癌細胞系(HT29,CACO2,SW480,HCT116)中MEG8表達顯著下調(P<0.05),如圖1(a)所示。與轉染Vector相比,在HT29細胞中轉染Ad-MEG8后,MEG8的表達顯著上調(P<0.01),如圖1(b)所示,顯示過表達效率較高。與Vector組相比,過表達MEG8后細胞增殖和侵襲顯著下降(P<0.05), 如圖1(c)~(e)所示,而細胞凋亡顯著上升(P<0.05), 如圖1(f)~(g)所示。
(2)MEG8與miR-1827結合并下調其表達如圖2所示。 與轉染NC mimic相比,轉染miR- 1827mimic后,MEG8-Wt報告載體熒光素酶活性顯著下降(P<0.05),如圖1(b)所示,而MEG8-Mut報告載體熒光素酶活性無顯著變化。RIP實驗結果顯示,分別過表達miR-1827和MEG8后,與IgG抗體組相比,Ago2抗體組的MEG8和miR-1827顯著富集(P<0.05),如圖1(c)所示。與轉染Vector相比,在HT29細胞中轉染Ad-MEG8顯著抑制miR-1827表達(P<0.05),如圖1(d)所示。而與轉染NC siRNA相比,轉染MEG8 siRNA顯著促進miR-1827表達(P<0.05),如圖1(d)所示。
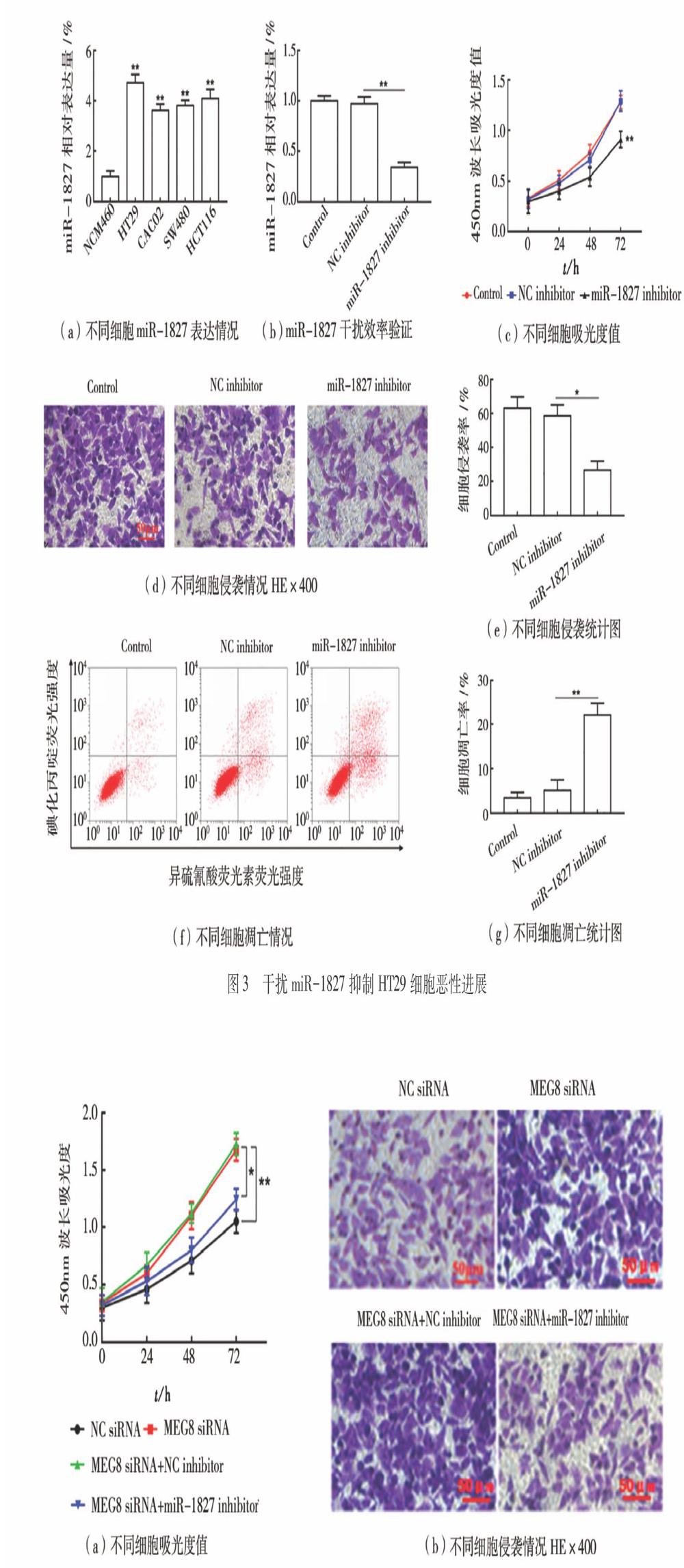
(3)干擾miR-1827抑制HT29細胞惡性進展如圖3所示。與人正常腸上皮細胞(NCM460)相比,結腸癌細胞系(HT29,CACO2,SW480,HCT116)中miR-1827表達顯著上調(P<0.05),如圖3(a)所示。與轉染NC inhibitor相比,在HT29細胞中轉染miR-1827 inhibitor顯著抑制miR-1827表達(P<0.05),如圖3(b)所示,顯示干擾效率較高。與NC inhibitor組相比,干擾miR-1827組的細胞增殖和侵襲顯著下降(P<0.05),如圖3(c)~(e)所示,而細胞凋亡顯著上升(P<0.05),如圖3(f)~(g)所示。
(4)MEG8通過抑制miR-1827調控HT29細胞進展如圖4所示。與轉染NC siRNA相比,在HT29細胞中轉染MEG8 siRNA顯著促進細胞增殖和侵襲(P<0.05),如圖4(a)~(c)所示,抑制凋亡(P<0.05),如圖4(d)~(e)所示。而與轉染MEG8 siRNA+NC inhibitor相比,共轉染MEG8 siRNA和miR-1827 inhibitor,細胞增殖和侵襲顯著下降(P<0.05),如圖4(a)~(c)所示,而凋亡顯著上升(P<0.05),如圖4(d)~(e)所示。
(5)MEG8通過抑制miR-1827調節ERK/JNK通路活性如圖5所示。與轉染NC siRNA相比,在HT29細胞中轉染MEG8 siRNA顯著抑制p-ERK1/2和p-JNK的蛋白質表達(P<0.05),如圖5(a)~(c)所示,而t-ERK1/2和t-JNK的蛋白質表達不變。與轉染MEG8 siRNA+NC inhibitor相比,共轉染MEG8 siRNA和miR1827 inhibitor,p-ERK1/2和p-JNK蛋白質表達顯著上升(P<0.05),如圖5(a)~(c)所示,而t-ERK1/2和t-JNK的蛋白質表達不變。
3 討論
結腸癌是一種常見的消化系統癌癥,在全球與癌癥相關死亡中排名第四[7]。癌細胞的無限增殖和高轉移特性導致結腸癌的高死亡率。在2014a和2015a,結腸癌導致約70萬人死亡[8]2 007。結腸癌的早期治療結果通常令人滿意,并且大于95%的患者在積極治療后生存率超過5a[8]2 007。然而,由于其隱蔽和發展迅速的特征,大多數患者在首次診斷時就處于晚期[4]5 752,導致多數患者生存結果較差。結腸癌發生是一個復雜過程,包括遺傳和表觀遺傳學變化。因此,闡明腫瘤發生的分子機制,為早期診斷,預后和治療評估尋找新的分子標志物意義重大[9]。
LncRNA通過多種機制介導基因表達,包括lncRNA-miRNA相互作用,lncRNA-蛋白質相互作用和lncRNA-mRNA相互作用。前人研究表明,lncRNAs參與各種生物過程調控,包括細胞增殖、侵襲、轉移、癌變和癌癥進展[10]。多種lncRNA失調與結腸癌進展相關。LncRNA BCAR4通過激活Wnt/β-catenin信號傳導途徑促進結腸癌腫瘤生長和轉移[11]。LncRNA HOTAIR在結腸癌中表達上調,與結腸癌遠端轉移和不良預后有關[4]5 754。LncRNA MALAT1通過海綿miR-129-5p調控HMGB1促進結腸癌發展[12]。本研究表明,與人正常腸上皮細胞相比,MEG8在結腸癌細胞中表達下調,與文獻報道一致。過表達MEG8顯著抑制結腸癌細胞系HT29增殖和侵襲,促進凋亡,顯示MEG8在結腸癌中作為腫瘤抑制基因,可能作為結腸癌的預測指標。研究還發現,MEG8與miR-1827直接結合,負調控miR-1827的表達,表明MEG8可能通過抑制miR-1827發揮腫瘤抑制作用。
MiRNA通過與mRNAs的3非翻譯區結合,在基因表達中發揮轉錄后調控作用,導致翻譯抑制或基因沉默。MiRNA基因只代表人類基因組一小部分,但它們調控幾乎1/3的人類基因。研究表明,miRNAs與多種生理病理過程相關, 包括細胞分化、 凋亡、 增殖、 胰島素分泌、 膽固醇合成和腫瘤生成。MiRNAs與結腸癌進展緊密相關。MiR-144通過下調SMAD4抑制結腸癌生長和轉移[13];MiR-192在結腸癌中發揮抑癌作用,辛伐他汀通過激活miR-192抑制癌細胞生長[14];MiR-215抑制低氧誘導的結腸癌干細胞活性[15]。文獻[16]報道,MiR-1827通過靶向MYC在肺腺癌中發揮抑癌作用。文獻[6]5 108報道,miR-1827通過調控Wnt信號通路,在結腸癌中發揮致癌作用。本研究結果顯示,與人正常腸上皮細胞相比,miR-1827在結腸癌細胞中表達上調,與前人研究結果吻合。在HT29細胞中干擾miR-1827,細胞增殖和侵襲下降,而凋亡上升。此外,轉染miR-1827 inhibitor逆轉沉默MEG8誘導的細胞增殖和侵襲上調,凋亡下降。
越來越多證據表明,ERK/JNK信號通路在結腸癌細胞進展中發揮重要作用。據報道,ERK-JNK信號通路上調通過CC趨化因子配體7和CC趨化因子受體3之間的串擾促進結腸癌細胞轉移。衣霉素通過下調ERK-JNK信號通路抑制結腸癌細胞生長和轉移[17]。Mahalingam等人證明,在結腸癌中,舒尼替尼通過增強JNK活性,降低凋亡抑制蛋白質水平,促進TRAIL介導的凋亡[18]。本研究數據顯示,干擾MEG8顯著抑制HT29細胞中p-ERK1/2、p-JNK蛋白質水平。而干擾miR-1827反轉MEG8對ERK/JNK通路的作用,表明MEG8和miR-1827對HT29細胞增殖和凋亡的影響很可能通過調控ERK/JNK通路實現。
綜上所述,與正常腸上皮細胞相比,MEG8在結腸癌細胞中表達下調,過表達 MEG8抑制HT29細胞增殖和侵襲,促進凋亡。機制研究顯示,MEG8可能通過負調控miR-1827,激活ERK/JNK通路,進而調控HT29結腸癌細胞增殖、侵襲和凋亡。此項研究,一定程度上補充了影響結腸癌細胞增殖、侵襲和凋亡的免疫學理論,也為結腸癌免疫治療方面提供一條潛在的方法。
參考文獻:
[1] PASCHKE S,JAFAROV S,STAIB L,et al.Are colon and rectal cancer two different tumor entities? A proposal to abandon the term colorectal cancer[J].Int J Mol Sci,2018,19(9):2 577.
[2] HURTADO CG,WAN F,HOUSSEAU F,et al.Roles for interleukin 17 and adaptive immunity in pathogenesis of colorectal cancer[J].Gastroenterology,2018,155(6):1 706-1 715.
[3] LIU J,DU W.LncRNA FENDRR attenuates colon cancer progression by repression of SOX4 protein[J].OncoTargets Ther,2019,12(1):4 287.
[4] PENG C,ZHAO X,WEI C,et al.LncRNA HOTAIR promotes colon cancer development by down-regulating miRNA-34a[J].Eur Rev Med Pharmacol Sci,2019,23(13):5 752-5 761.
[5] KALMR A,NAGY ZB,GALAMB O,et al.Genome-wide expression profiling in colorectal cancer focusing on lncRNAs in the adenoma-carcinoma transition[J].BMC Cancer,2019,19(1):1 059.
[6] FASIHI A,M.SOLTANI B,ATASHI A,et al.Introduction of hsa-miR-103a and hsa-miR-1827 and hsa-miR-137 as new regulators of Wnt signaling pathway and their relation to colorectal carcinoma[J].J Cell Biochem,2018,119(7):5 104-5 117.
[7] WEI F,WU Y,TANG L,et al.Trend analysis of cancer incidence and mortality in China[J].Sci China Life Sci,2017,60(11):1 271-1 275.
[8] BAI J,XU J,ZHAO J,et al.Downregulation of lncRNA AWPPH inhibits colon cancer cell proliferation by downregulating GLUT-1[J].Oncol Lett,2019,18(2):2 007-2 012.
[9] LEE JJ,CHU E.The adjuvant treatment of stage III colon cancer:might less be more?[J].Oncology,2018,32:437-422.
[10] CUI M,CHEN M,SHEN Z,et al.LncRNA-UCA1 modulates progression of colon cancer through regulating the miR-28-5p/HOXB3 axis[J].J Cell Biochem,2019,120(5):6 926-6 936.
[11] BO H,FAN L,LI J,et al.High expression of lncRNA AFAP1-AS1 promotes the progression of colon cancer and predicts poor prognosis[J].J Cancer,2018,9(24):4 677.
[12] WU Q,MENG WY,JIE Y,et al.LncRNA MALAT1 induces colon cancer development by regulating miR-129-5p/HMGB1 axis[J].J Cell Physiol,2018,233(9):6 750-6 757.
[13] SHENG S,XIE L,WU Y,et al.MiR-144 inhibits growth and metastasis in colon cancer by down-regulating SMAD4[J].Bioscience reports,2019,39(3):1-8.
[14] ZHENG XF,LIU KX,WANG XM,et al.MicroRNA-192 acts as a tumor suppressor in colon cancer and simvastatin activates miR-192 to inhibit cancer cell growth[J].Mol Med Rep,2019,19(3):1 753-1 760.
[15] ULLMANN P,NURMIK M,SCHMITZ M,et al.Tumor suppressor miR-215 counteracts hypoxia-induced colon cancer stem cell activity[J].Cancer Lett,2019,450(1):32-41.
[16] FAN G,XU P,TU P.MiR-1827 functions as a tumor suppressor in lung adenocarcinoma by targeting MYC and FAM83F[J].J Cell Biochem, 2020, 121(2):1 675-1 689.
[17] YOU S,LI W,GUAN Y.Tunicamycin inhibits colon carcinoma growth and aggressiveness via modulation of the ERK-JNK-mediated AKT/mTOR signaling pathway[J].Mol Med Rep,2018,17(3):4 203-4 212.
[18] MAHALINGAM D,CAREW JS,ESPITIA CM,et al.Heightened JNK Activation and Reduced XIAP Levels Promote TRAIL and Sunitinib-Mediated Apoptosis in Colon Cancer Models[J].Cancers,2019,11(7):895.
(責任編輯:丁 寒)
猜你喜歡
天津醫科大學學報(2019年6期)2019-08-13 07:04:32
西南國防醫藥(2016年7期)2016-12-01 06:01:15
腹腔鏡外科雜志(2016年10期)2016-06-01 12:10:08
中國衛生標準管理(2015年6期)2016-01-14 05:17:12
中國衛生標準管理(2015年1期)2016-01-14 03:41:26
中國當代醫藥(2015年26期)2015-03-01 02:06:57
西南軍醫(2015年6期)2015-01-23 01:25:50
河南醫學研究(2014年3期)2014-02-27 14:51:48
沈陽醫學院學報(2014年1期)2014-02-16 06:19:24
中華介入放射學電子雜志(2014年1期)2014-02-02 05:24:06
