砂蘚RcPLD基因的抗旱功能分析
2021-09-04 03:16:14馬天意張時通朱巍巍張梅娟李瑩瑩彭疑芳
西北植物學報 2021年7期
馬天意,張時通,朱巍巍,張梅娟,李瑩瑩,彭疑芳,沙 偉
(1 抗性基因工程與寒地生物多樣性保護黑龍江重點實驗室, 齊齊哈爾大學 生命科學與農林學院, 黑龍江齊齊哈爾 161006; 2 東北農業大學 生命科學學院, 哈爾濱 150030)
干旱是頻發的全球性自然災害之一,嚴重威脅植物的生存和生長,導致糧食產量下降[1]。中國糧食生產過程中遭受到的干旱災害尤為嚴重,多年來中國農業干旱受災面積約占總自然災害受災面積的一半左右,干旱導致的農作物種植面積縮減問題直接制約了糧食的增產速度[2-3]。利用環境干旱脅迫已經篩選出了許多具有抗旱性的植物物種,再利用遺傳學和分子生物學手段從具有抗旱能力的植物中尋找能夠提高植物抗旱性的基因,借以培育新的抗旱作物品種,是使植物適應干旱環境生長的一種行之有效的方法[4]。
砂蘚(Racomitriumcanescens)為紫萼蘚科(Grimmiaceae)砂蘚屬(Racomitrium)植物,廣泛分布于各種生境,是黑龍江省五大連池地區火山熔巖地貌的優勢種[5],也在中國大部分地區廣泛分布[6]。早在幾十年前就已經證實砂蘚是具有耐脫水性的蘚類植物[7],干燥數年的砂蘚材料可在復水后數分鐘內恢復正常生長狀態,在植物抗旱性和耐脫水性機制研究方面顯示出較高的利用價值[8],砂蘚中與強耐脫水性功能相關的基因可能具有提高植物抗旱能力的作用。在早期研究中,對正常生長的砂蘚材料進行了24 h的快速脫水處理,使用轉錄組測序技術來尋找可能參與砂蘚耐脫水性響應的功能基因,并試圖探討這些基因提高植物抗旱性的能力。對發現的多個基因進行了初步的研究,其中砂蘚的ATP合酶Ⅱ亞基基因RcatpG[9]、生長素受體基因RcTIR1[10]、超氧化物歧化酶基因RcSOD[11]、丙二烯氧化物環化酶基因RcAOC1[12]、MYB轉錄因子基因RcMYB[13]、bZIP轉錄因子基因RcbZIP1[14]、過氧化氫酶基因RcCAT[15]和小G蛋白結合蛋白質基因RcRAB[16]的表達在各種脫水和復水處理中發生了變化,而且使用RcMYB構建的過表達轉基因煙草獲得了更強的耐旱性和復水能力[17]。這些研究結果表明砂蘚中的基因資源能夠用來提高植物的抗旱能力,很多基因抗旱功能有待于被發現和證實。
近期報道了砂蘚中一個被注釋為磷脂酶D(phospholipase D, PLD)的編碼基因RcPLD能夠在砂蘚的不同脫水和復水處理中發生顯著的差異表達,通過對該基因的編碼序列克隆和序列基本特征分析[8],初步證實RcPLD可能參與砂蘚的耐脫水脅迫響應。
PLD的生物化學本質是磷酸二酯酶,其根據水解磷脂時的不同作用位置被分為磷脂酶A1、A2、B、C、D等5類[18]。這些磷酸二酯酶能夠將磷脂水解為二脂酰甘油、乙酰膽堿、磷脂酸和三磷酸肌醇等產物,這些水解產物都能夠作為第二信使在植物細胞的各種生命活動中發揮重要的作用[19]。典型的PLD具有的保守結構域包括2個PLD家族結構域HKD(HxKxxxxD, domain)、一個C2(proteinkinase C-conserved-domain 2)結構域、一個磷脂酰肌醇二磷酸[phosphatidylinositol-(4,5)-bisphosphate, PIP2]結構域及潛在的IYIENQFF結構,少數PLD會缺少其中的某些結構域而由其他結構域代替[8]。研究發現RcPLD所編碼的蛋白質序列中僅包含1個HKD,這種非典型的PLD在之前鮮有報道,其功能與典型PLD的異同也不得而知。前人發現,不同植物物種的PLD能夠影響植物的冷脅迫和干旱脅迫耐受能力,并對其中的作用機制進行了相關的研究[20-21],但苔蘚植物中PLD的相關研究還較為缺乏,特別是砂蘚的非典型PLD(RcPLD)在環境脅迫中的具體作用尚不清楚。
因此,本研究使用已克隆得到的RcPLD編碼序列構建了過表達轉基因擬南芥(Arabidopsisthaliana)株系rcpld-oe,發現rcpld-oe具有強于野生型擬南芥的干旱脅迫耐受能力,對不同的擬南芥株系進行了基本的生理生化指標測定,以對RcPLD增強擬南芥抗旱性的機制進行解釋。本研究結果初步證明了RcPLD可用于提高植物耐旱能力,為更多苔蘚植物抗逆相關基因資源的研究、開發和利用奠定基礎。
1 材料和方法
1.1 RcPLD過表達轉基因擬南芥株系的構建
使用前期已連入RcPLD編碼序列的pMD-19T重組質粒構建RcPLD的過量表達載體。在構建克隆載體時已在引物RcPLD-F和RcPLD-R中分別加入限制性內切酶SalⅠ和EcoR Ⅰ的酶切位點(表1),使用TaKaRa公司(大連)生產的QuickCutSalⅠ和QuickCutEcoR Ⅰ限制性內切酶對RcPLD的克隆載體和pRI101-AN空表達載體(TaKaRa,大連)進行雙酶切,反應結束后使用1%瓊脂糖凝膠電泳對產物進行分離,回收目的條帶后使用北京索萊寶科技有限公司的DNA瓊脂糖凝膠回收試劑盒對酶切后的載體和RcPLD編碼序列進行回收,使用TaKaRa公司生產的T4DNA Ligase對基因和載體片段進行連接。連接產物轉化至大腸桿菌(Escherichiacoli)DH5α株系感受態細胞后進行培養和篩選鑒定,方法同沙偉等[8]。根據Kong等方法,將提取后的重組質粒轉化至實驗室保存的根癌農桿菌(Agrobacteriumtumefaciens)EHA105后進行篩選、鑒定和培養,使用浸花法對實驗室保存的columbia-0(col-0)野生型擬南芥進行侵染,使用含有50 μg/mL卡那霉素的1/2 MS培養基對侵染后獲得的擬南芥種子進行篩選,篩選后獲得的植株即為T1代RcPLD過表達轉基因擬南芥植株(rcpld-oe)[22]。
表1 實驗引物序列信息
篩選結束后將T1代rcpld-oe植株移栽至滅菌的混合土基質(蛭石∶草炭土比例為2∶1)中,放置于封閉光照培養室中進行培養(16 h光照/8 h黑暗,(25±1) ℃,空氣相對濕度45%~55%)。當T1代rcpld-oe植株長出6片真葉時取1片真葉進行基因組DNA提取,以檢測篩選得到的擬南芥植株中是否插入了RcPLD片段。使用TaKaRa公司生產的DNAiso Reagent試劑對rcpld-oe和野生型擬南芥植株進行DNA提取,提取后將基因組DNA稀釋至100 ng/μL,每植株取100 ng基因組DNA進行PCR反應。反應體系為:基因組DNA 1 μL,RcPLD-F、RcPLD-R引物各1 μL,Premix TaqTM(Ex TaqTMVersion 2.0)反應液10 μL,無菌去離子水7 μL;PCR反應程序為:95 ℃預變性5 min,95 ℃變性30 s,55 ℃退火30 s,72 ℃延伸1 min,共30循環,72 ℃終延伸10 min。使用等量野生型擬南芥基因組DNA作為陰性對照,無菌去離子水作為空對照,含有RcPLD編碼序列的pRI101-AN重組質粒作為陽性對照。
將鑒定含有RcPLD序列的植株繼續培養,單獨收集每一株擬南芥植株的種子,然后播種于含有50 μg/mL卡那霉素的1/2 MS培養基中進行篩選,篩選后的植株移入混合土基質中繼續培養,獲得T2代rcpld-oe植株。在培養過程中,當植株生長至21 d時,每個植株剪去4片未衰老的蓮座葉,迅速放入2 mL離心管中,用液氮速凍后放置于-80 ℃冰箱中保存,供后續試驗中提取RNA。單獨收集每株T2代rcpld-oe植株的種子,使用含有50 μg/mL卡那霉素的1/2 MS培養基對種子進行篩選,每個培養基上播種同一植株的50粒種子,每個植株播種3次,若排除自然萌發率影響后,同一植株的3次篩選中所有種子都能正常萌發生長,則認為該T2代rcpld-oe植株為RcPLD純合體,可用于后續試驗分析。當T2代rcpld-oe植株被鑒定為純合體后,使用提前采集并保存的蓮座葉材料進行RNA提取,提取方法同沙偉等[8]。用獲得的RNA合成cDNA后,采用qPCR對材料中RcPLD的表達量進行檢測,以擬南芥Actin1(AT2G37620)基因作為內參基因,進行獨立的3次生物學重復,試驗方法同沙偉等[8],所用引物序列如表1所示。
1.2 干旱和復水處理
將消毒后的rcpld-oe純合T3代種子和野生型擬南芥種子同時播種于1/2 MS平板固體培養基中,消毒方法同Kong等[22],于4 ℃黑暗條件下春化3 d后移至光照周期為8 h光照/16 h黑暗的培養室中進行培養,待幼苗生長至具有2片蓮座葉后,將幼苗移至含混合土基質的帶托盤花盆中進行培養。當植株生長至播種21 d時,選取不同擬南芥株系中長勢一致的足量植株進行干旱脅迫處理。將含植株花盆移至不含水的托盤中進行自然干旱處理,同時以相同培養條件下托盤中正常澆水的相同株系植株作為對照,此時設置為干旱處理的0 d(D0),隨后每天對植株的生長情況進行觀察和圖像采集,每7 d收集一批擬南芥蓮座葉材料,共收集5次(D0、D7、D14、D21、D28),干旱處理28 d后使用去離子水對擬南芥植株進行復水處理,此時記為復水0 d(R0),復水2 d后(R2)觀察植株形態特征并進行圖像采集。
1.3 生理生化指標測定
采集干旱處理中各株系擬南芥的蓮座葉作為生理生化指標測定的材料。使用磺基水楊酸法對游離脯氨酸含量進行測定,使用蒽酮法對可溶性糖含量進行測定,使用考馬斯亮藍法對可溶性蛋白質含量進行測定,使用三氯乙酸/硫代巴比妥酸法對丙二醛進行提取和含量測定,使用丙酮提取法對總葉綠素進行提取,以上生理生化指標測定和計算方法及相對含水量測定方法如Yin等所述[23]。每個指標的每次測定中分別使用0.1 g樣品材料,干旱處理和正常生長條件下的每個株系的植株均單獨取樣測定,測定均進行3次生物學重復。
1.4 數據統計與分析
用SPSS 20.0軟件對獲取的數據進行統計學計算分析,所有數據用Duncan法進行one-way ANOVA計算分析不同樣品間的差異顯著性。
2 結果與分析
2.1 RcPLD過表達擬南芥純合體株系的構建
以前期成功構建的RcPLD克隆載體作為模板,構建含有RcPLD編碼序列的植物過量表達載體。在成功獲得RcPLD表達載體后,將重組質粒轉化入根癌農桿菌EHA105株系中,對抗生素篩選后的陽性菌落進行PCR鑒定,獲得含有RcPLD表達載體的農桿菌菌株(圖1,A)。對篩選獲得的農桿菌菌落進一步培養,并用浸花法侵染col-0野生型擬南芥,收集成熟后種子,使用含有50 μg/mL卡那霉素的1/2 MS培養基對T1代轉基因植株進行篩選(圖1,B)。提取篩選獲得的植株基因組DNA,用RcPLD的克隆引物來鑒定RcPLD基因是否已插入篩選得到的植株基因組DNA中,最終獲得3個T1代RcPLD過表達轉基因擬南芥植株rcpld-oe-1、rcpld-oe-2和rcpld-oe-3(圖1,C)。
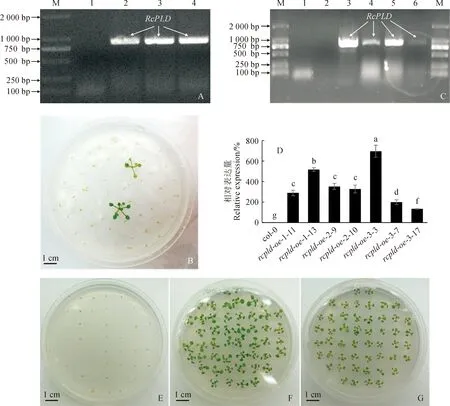
A.含有RcPLD表達載體的農桿菌菌液PCR:M.DL2000; 1. pRI101-AN(對照);2. RcPLD表達載體; 3-4. 含有RcPLD表達載體的農桿菌菌液; B. T1代rcpld-oe植株的抗生素篩選; C. T1代rcpld-oe植株基因組DNA的PCR驗證: M. DL2000; 1. 水(對照); 2. 野生型擬南芥基因組DNA; 3. RcPLD表達載體; 4-6. T1代rcpld-oe植株基因組DNA; D. T2代rcpld-oe植株中RcPLD相對表達量的qRT-PCR檢測, rcpld-oe-3-17植株中RcPLD的表達量被定義為100%, 誤差線表示標準偏差, n=3, 誤差線上不同小寫字母表示在0.05水平存在顯著差異(P<0.05),下同; E-G. rcpld-oe純合體株系的篩選:E.野生型擬南芥; F. rcpld-oe-2-10; G. rcpld-oe-3-17圖1 RcPLD過表達擬南芥純合體株系的構建A. PCR with suspension of Agrobacterium containing RcPLD expression vectors: M. DL2000; 1. pRI101-AN (control); 2. RcPLD expression vector; 3-4. Agrobacterium suspension containing RcPLD expression vectors; B. Screening of T1 generation rcpld-oe plants by antibiotics; C. Genomic DNA PCR identification of T1 generation rcpld-oe plants: M. DL2000; 1. Water (control); 2. Genomic DNA of wild-type Arabidopsis; 3. RcPLD expression vector; 4-6. Genomic DNA of T1 generation rcpld-oe plants; D. qRT-PCR detection of relative expression of RcPLD, RcPLD expression of rcpld-oe-3-17 were defined as 100%, error bars indicate standard deviations, n=3, the different lowercased letters above error bars stand for significant difference among materials at 0.05 level (P<0.05), the same as below; E-G. Screening of rcpld-oe homozygotes: E. Wild-type Arabidopsis; F. rcpld-oe-2-10;G. rcpld-oe-3-17Fig.1 Construction and identification of RcPLD-overexpressed Arabidopsis thaliana homozygote lines
對3個過表達擬南芥株系繼續培養,收集其成熟種子并再次進行播種和篩選,繼續培養T2代擬南芥植株的同時收集蓮座葉材料和種子,使用獲得的T3代種子來鑒定哪些T2代rcpld-oe株系為RcPLD的純合體株系,對鑒定為純合體的T2代株系進行RNA提取,并對植株中RcPLD進行qPCR檢測(圖1,D),部分純合體rcpld-oe株系篩選結果如圖1,E-G所示。由圖1,D可以看出,野生型(col-0)擬南芥中RcPLD的表達量可被認定為儀器系統誤差,其他所有被檢測的T2代rcpld-oe植株中都具有較高的RcPLD表達量。綜合考慮基因表達量和所獲T3代植株的種子數量,最終選取rcpld-oe-1-13、rcpld-oe-2-9和rcpld-oe-3-3進行后續研究。
2.2 RcPLD過表達擬南芥的耐旱性分析
將野生型(col-0)植株和過表達株系rcpld-oe-1-13、rcpld-oe-2-9和rcpld-oe-3-3植株同時進行干旱處理,檢測RcPLD過量表達是否能夠提高擬南芥的干旱脅迫耐受能力。圖2顯示,在正常澆水條件下,擬南芥3個rcpld-oe株系植株的生長速率慢于野生型植株,其相同苗齡植株的蓮座葉面積明顯小于野生型植株,且抽薹時間明顯較晚(圖2,A)。在干旱處理0~21 d,3個rcpld-oe株系的植株蓮座葉片面積小于野生型植株,而野生型植株又明顯小于其同齡相應的正常澆水處理植株;但3個rcpld-oe株系植株的體積在干旱處理7 d時大于其正常澆水處理的植株,而在干旱處理14~21 d時均小于其正常澆水處理的植株(圖2,A、B)。在干旱處理28 d時,野生型擬南芥已全部死亡,rcpld-oe-1-13和rcpld-oe-2-9植株的生長受到較低程度的抑制,rcpld-oe-3-3植株受到較高程度干旱抑制,但好于同期野生型擬南芥;復水2 d后,3個rcpld-oe株系的植株絕大部分能夠維持生存并繼續生長,而野生型擬南芥植株基本無復蘇征兆(圖2,B)。以上結果說明RcPLD的過量表達使擬南芥的抗旱能力和旱后復水能力在一定程度上得到了提升。
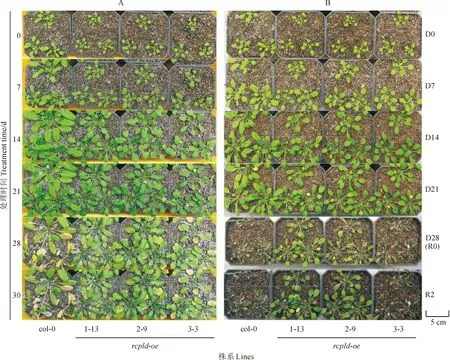
A.正常澆水處理的擬南芥植株; B.干旱脅迫處理的擬南芥植株:D0、D7、D14、D21、D28分別表示干旱處理0、7、14、21、28 d, R0、R2分別表示復水處理0、2 d;1-13、2-9、3-3分別代表RcPLD過表達株系rcpld-oe-1-13、rcpld-oe-2-9和rcpld-oe-3-3植株。下同圖2 干旱和復水處理下擬南芥野生型(col-0)和rcpld-oe過表達株系的生長情況A. A. thaliana plants growing under normal watering condition; B. A. thaliana plants growing under drought stress treatments: D0, D7, D14, D21, D28 stand for drought treatment for 0, 7, 14, 21, 28 d, respectively, while R0, R2 stand for rehydration treatment for 0, 2 d, respectively; 1-13, 2-9, 3-3 are representing RcPLD overexpression plants of rcpld-oe-1-13, rcpld-oe-2-9 and rcpld-oe-3-3 lines. The same as belowFig.2 Growth condition of wild-type (col-0) and rcpld-oe Arabidopsis thaliana lines under drought and rehydration treatments
2.3 干旱脅迫過程中RcPLD過表達擬南芥的生理生化指標變化特征
RcPLD過量表達株系和野生型擬南芥植株蓮座葉基本生理生化指標測定結果顯示:正常澆水條件下,野生型和rcpld-oe植株蓮座葉的相對含水量均能維持在95%以上;在干旱處理過程中,野生型和rcpld-oe植株蓮座葉的相對含水量隨著處理時間的延長都顯著下降,但干旱處理21 d時rcpld-oe擬南芥相對含水量的下降幅度顯著低于野生型植株,其中又以rcpld-oe-1-13植株下降幅度最小(圖3,A)。野生型和rcpld-oe植株蓮座葉脯氨酸含量在正常生長條件下基本無變化且維持在較低的水平,而在干旱處理過程中都有顯著的提升,尤其在干旱處理后期迅速大幅度提升,且各rcpld-oe植株在處理7 d時提升幅度就達到顯著水平,而野生型植株在處理14 d時才顯著升高;在干旱處理過程中,各rcpld-oe植株蓮座葉脯氨酸含量增加幅度始終顯著高于同期野生型擬南芥(圖3,B)。野生型和rcpld-oe株系植株蓮座葉可溶性糖和可溶性蛋白質含量隨著處理時間的變化趨勢和幅度基本一致,rcpld-oe-1-13和rcpld-oe-2-9中可溶性糖含量在干旱處理過程中基本高于野生型擬南芥,可溶性蛋白質含量在干旱處理后期顯著高于野生型擬南芥,而rcpld-oe-3-3中的含量低于野生型擬南芥(圖3,C、D)。隨著擬南芥植株進入生殖生長階段和逐漸衰老,野生型和過表達植株蓮座葉中丙二醛含量都有增加的趨勢;在正常澆水條件下,野生型植株中丙二醛含量隨著植株的生長而穩定增加,而rcpld-oe植株在生長前期含量變化幅度小于野生型擬南芥,整體丙二醛含量也較低;在干旱處理過程中,野生型擬南芥蓮座葉中丙二醛含量隨著處理時間的延長而顯著提升且速率較穩定,rcpld-oe植株的丙二醛含量在干旱處理中也有所提升,但提升幅度始終低于野生型擬南芥(圖3,E)。在總葉綠素含量方面,rcpld-oe植株在正常澆水條件下與野生型擬南芥表現有所不同,其蓮座葉總葉綠素積累的速度較慢;在干旱處理過程中,野生型擬南芥中總葉綠素含量的積累受到了顯著的抑制,rcpld-oe植株中的總葉綠素含量也有所降低,但受抑制程度顯著低于野生型擬南芥(圖3,F)。以上結果說明明RcPLD的過量表達使擬南芥在干旱過程中的滲透調節物質含量顯著提高,進而使水分的損失程度降低,同時通過降低膜損傷程度和光合作用抑制水平來對植株進行保護。
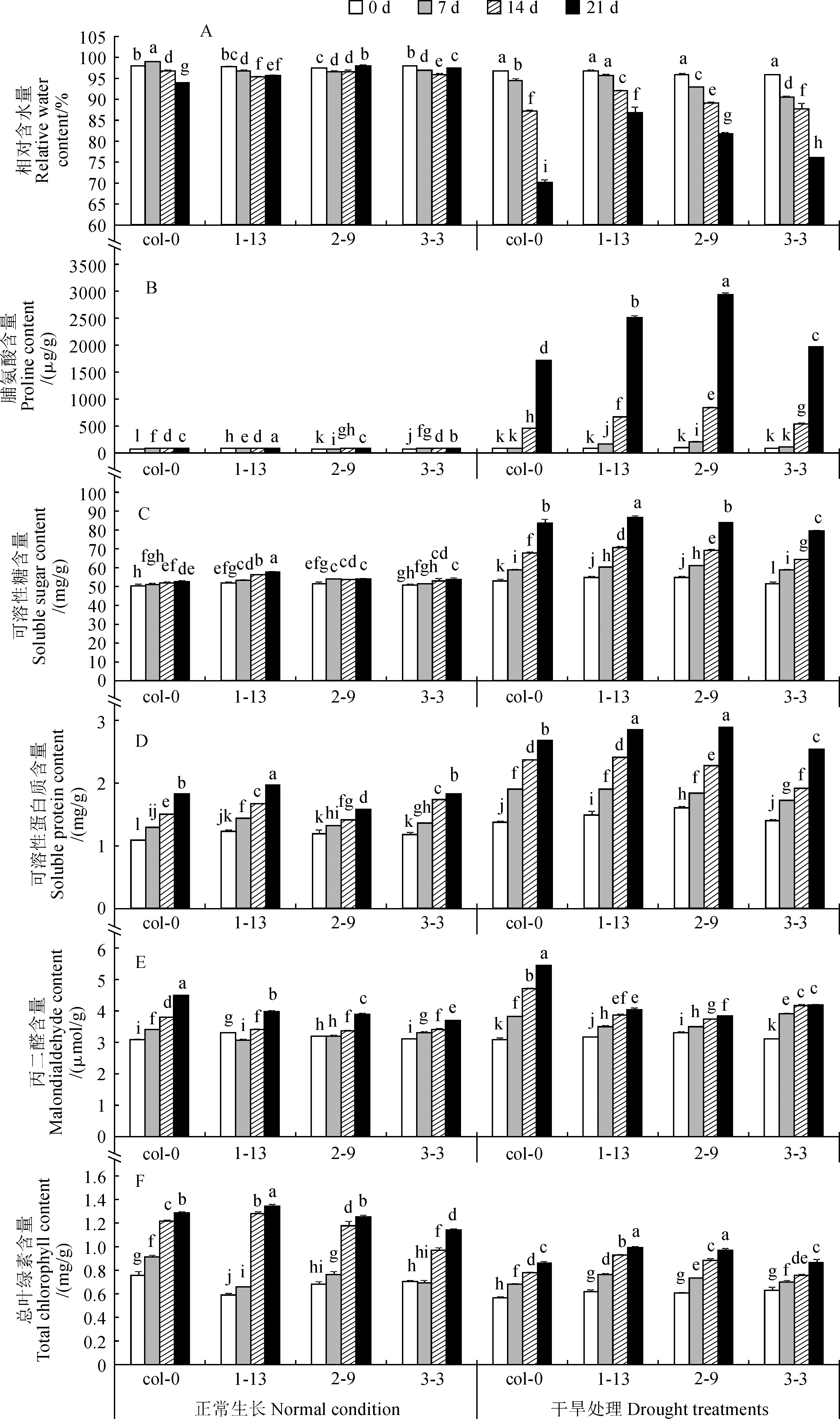
圖3 干旱處理過程中野生型和rcpld-oe擬南芥植株蓮座葉相關指標的變化Fig.3 Changes of related index in rosette leaf of wild-type and rcpld-oe Arabidopsis thaliana under the drought treatment
3 討 論
在不同植物中,不同的PLD基因被證實能夠影響植物的脅迫耐受能力。擬南芥中PLDα缺陷突變體的葉片蒸騰失水率顯著高于野生型,pldα1-1在干旱脅迫下表現出比野生型更高的敏感性[24-25];菊花(Chrysanthemummorifolium)CmPLDα基因參與菊花的多種脅迫過程,包括機械損傷、高溫脅迫、缺磷脅迫以及鹽脅迫[26];而在苔蘚植物中PLD的抗逆功能還缺乏報道。本研究構建的不同RcPLD過表達擬南芥株系rcpld-oe具有不同的RcPLD表達量,但在正常生長條件下表現的特征基本一致,基本表現為營養階段下植株體積明顯小于野生型擬南芥,蓮座葉較小,營養生長時期較長,抽薹期延后等。
在干旱處理條件下,本研究發現擬南芥不同rcpld-oe株系植株總體抗旱能力明顯強于野生型擬南芥,3周齡的rcpld-oe幼苗在自然干旱處理28 d后基本能夠存活,多數植株在復水后能夠恢復生長,而野生型擬南芥無法恢復。與正常生長的植株相比,野生型擬南芥植株體積在干旱脅迫過程中明顯下降,而rcpld-oe植株在形態上受干旱脅迫抑制程度較小,甚至在干旱處理初期植株體積還略大于野生型擬南芥。值得注意的是,RcPLD賦予擬南芥抗旱性的情況并不完全與RcPLD的表達量呈正相關,在進行干旱處理的3個rcpld-oe株系中,rcpld-oe-3-3中RcPLD的表達量最高,但在3個株系中的抗旱能力表現為最弱,我們推測RcPLD對擬南芥抗旱能力的提升需要一個最適區間,也可能存在RcPLD在擬南芥基因組插入位點的影響,但3個rcpld-oe株系在表型特征上的一致性顯示rcpld-oe-3-3在抗旱性方面的表現更可能與表達量相關,這個推論將在后續的工作中進一步進行分析驗證。
本研究對野生型和rcpld-oe株系擬南芥干旱處理前21 d的材料進行了部分生理生化指標的測定,以初步分析RcPLD在提高擬南芥抗旱功能中可能的作用。首先,在水分保持能力方面,正常生長條件下野生型和rcpld-oe株系蓮座葉相對含水量水平相似,都能夠維持在90%以上,相對而言rcpld-oe株系相對含水量在生長過程中變化更平穩;在干旱處理過程中,野生型植株相對含水量下降的程度顯著高于rcpld-oe植株,在干旱處理21 d時已接近70%,而除rcpld-oe-1-13和rcpld-oe-2-9植株的相對含水量都能夠維持在80%以上,說明RcPLD的過量表達可能增強了擬南芥在干旱條件下防止水分散失的能力。曾有研究表明PLD的活性增加能夠促進脫落酸誘導的氣孔關閉過程來降低植物在干旱脅迫下的水分散失[27],我們認為這可能是RcPLD在過表達植株中提升抗旱性的機制之一。同時,氣孔關閉程度提升會導致CO2攝入量減少,進而影響有機物同化量而導致植物體生長速率的減慢[28],這也與我們觀察到的形態特征表現相一致,但RcPLD更精確的抗旱功能機制還需進一步探索。
其次,在干旱脅迫條件下,細胞中的滲透調節物質含量對于植物水分調節能力十分重要,隨著干旱脅迫程度的加深,植物細胞往往要增加細胞中的滲透調節物質含量來應對外界提升的滲透勢,游離脯氨酸、可溶性糖和可溶性蛋白質是常在植物遭受滲透脅迫過程中發揮作用的調節物質[23]。本研究對野生型和rcpld-oe擬南芥植株在干旱處理過程中這3類物質的含量進行了檢測,發現野生型植株和rcpld-oe植株在可溶性糖和可溶性蛋白質含量變化方面相差不大,雖然在干旱處理過程中存在顯著差異,但在差異幅度方面較小,而rcpld-oe植株在干旱處理過程中脯氨酸含量增加的速度和積累量都明顯優于野生型植株,這說明過量表達的RcPLD在擬南芥的抗旱過程中可能參與了滲透調節物質的調控,但對可溶性糖和可溶性蛋白質總量的影響程度較低,可能主要參與了增加游離脯氨酸累積的過程。
再次,丙二醛作為膜脂過氧化的終產物常被用于指示細胞膜完整性的代謝產物,通常認為丙二醛含量越低,膜脂過氧化程度越低,膜完整性越高[23]。從本研究結果可知,正常生長條件下野生型擬南芥蓮座葉片丙二醛含量的增加幅度就明顯高于rcpld-oe植株,這可能是由于野生型擬南芥與rcpld-oe植株相比蓮座葉片本身衰老的速度就較快有關;在干旱處理過程中可以發現,野生型擬南芥葉中丙二醛含量提升的速度和幅度顯著增強,而rcpld-oe植株葉中丙二醛含量增加的幅度顯著小于野生型植株,這說明在干旱處理過程中RcPLD一定程度上降低了rcpld-oe植株葉片細胞膜系統的過氧化傷害。
另外,在不同植物應對不同環境脅迫的過程中,光合作用采取的機制不盡相同,本研究檢測了干旱處理過程中擬南芥野生型和rcpld-oe植株蓮座葉總葉綠素的含量,探索RcPLD在提升擬南芥抗旱能力的過程中對光合作用可能產生的影響。研究發現在正常生長條件下,rcpld-oe植株總葉綠素的累積速度比野生型植株慢,這可能與rcpld-oe植株蓮座葉生長較慢的現象有關;與正常生長條件下相比,干旱處理過程中野生型和rcpld-oe植株總葉綠素的含量都有所減少,但相對而言野生型植株中葉綠素含量的減少程度更大。
綜上所述,本研究對前期發現響應砂蘚脫水脅迫的磷脂酶D編碼基因RcPLD在植物應對干旱脅迫時的作用進行了初步探討,利用已克隆的RcPLD編碼序列構建了植物中的過量表達載體,構建了RcPLD的過表達轉基因擬南芥株系rcpld-oe,并獲得了多個T3代rcpld-oe純合體株系。這些擬南芥rcpld-oe植株的抗旱能力整體上強于野生型擬南芥植株,在正常生長條件下與野生型擬南芥植株相比在營養生長階段體積較小,但營養生長期較長,抽薹較晚,蓮座葉衰老速率較慢。同時,過量表達的RcPLD使擬南芥在干旱脅迫條件下的水分散失速率降低,在一定程度上降低了干旱脅迫的膜損傷和光合作用抑制程度,但對滲透調節物質的含量變化影響較小。因此,本研究認為砂蘚中的RcPLD可以用于提高植物的抗旱性,其更深入的作用機制值得進一步研究。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
